
The mononuclear phagocyte system in hepatocellular carcinoma
2022-12-09 02:52:12DuanRuiQiaoGuanYueShanShuaiWangJunYaChengWeiQunYanHaiJunLi
World Journal of Gastroenterology 2022年45期
Duan-Rui Qiao, Guan-Yue Shan, Shuai Wang,Jun-Ya Cheng, Wei-Qun Yan, Hai-Jun Li
Abstract The mononuclear phagocyte system (MPS) consists of monocytes, dendritic cells and macrophages, which play vital roles in innate immune defense against cancer.Hepatocellular carcinoma (HCC) is a complex disease that is affected or initiated by many factors, including chronic hepatitis B virus infection, hepatitis C virus infection, metabolic disorders or alcohol consumption. Liver function, tumor stage and the performance status of patients affect HCC clinical outcomes. Studies have shown that targeted treatment of tumor microenvironment disorders may improve the efficacy of HCC treatments. Cytokines derived from the innate immune response can regulate T-cell differentiation, thereby shaping adaptive immunity, which is associated with the prognosis of HCC. Therefore, it is important to elucidate the function of the MPS in the progression of HCC. In this review, we outline the impact of HCC on the MPS. We illustrate how HCC reshapes MPS cell phenotype remodeling and the production of associated cytokines and characterize the function and impairment of the MPS in HCC.
Key Words: Mononuclear phagocyte system; Hepatocellular carcinoma; Monocyte;Dendritic cell; Macrophage
INTRODUCTION
Hepatocellular carcinoma (HCC) is the most common primary liver cancer and is predicted to be the sixth most-diagnosed cancer and the fourth-leading cause of death among all types of cancers[1-3]. HCC is influenced or initiated by many factors, including chronic hepatitis B virus (HBV) infection, hepatitis C virus (HCV) infection, metabolic disorder or chronic alcohol consumption[4-7]. More than 85% of HCC cases are accompanied by HBV infection in China[8-9]. Evidence has shown that HCC is a type of tumor with low or moderate immunogenicity[10]. Chronic inflammation creates an immunosuppressive microenvironment in the liver, facilitating HCC tumorigenesis and progression[11]. Although great progress has been made in the treatment of HCC in recent decades, the long-term survival rate of HCC is still poor[12]. Many factors, such as liver function, tumor stage and the performance status of patients,affect HCC clinical outcomes. According to previous studies, targeted treatment of dysregulated tumor microenvironments may improve the efficacy of HCC treatments[11,12].
The mononuclear phagocyte system (MPS) includes monocytes, dendritic cells (DCs) and macrophages and was proposed by Van Furth in the late 1960s and early 1970s[13,14]. Monocytes, DCs and macrophages are important members of the innate immune system and are capable of modulating adaptive immune responses. The immune system is precisely balanced between immune activation and tolerance. Within this system, antigen presenting cells (APCs) play a critical role in orchestrating the immune response[15]. Although stellate cells, endothelial cells and hepatocytes in the liver have the potential to present antigens to T cells, the MPS plays a major role in determining the nature of the immune response and keeps the liver as an immune tolerance organ. The MPS appears to be superior at sampling the environment through phagocytosis and presenting antigens to T cellsviaMHC class II molecules[16]. APCs of the MPS appear to have a commensurately increased expression of antigen presentation and costimulatory molecules and are potent secretors of modulatory cytokines[17].
In addition to its role in tissue development, homeostasis, inflammation and innate immune defense against pathogens, the MPS also plays a vital role in cancer[17,18]. Innate immune cell-derived cytokines can regulate T-cell differentiation, thereby shaping adaptive immunity, which is associated with the prognosis of HCC. It is important to illustrate the critical role of the MPS in the progression of HCC. In this review, we discuss how the HCC microenvironment remodels MPS cell phenotypes and cytokine production and describe the function and impairment of MPS components in HCC.
IMPACT OF THE MPS ON HCC
Monocytes and HCC
Monocytes originate in the bone marrow and spleen and account for approximately 5%-10% of human peripheral blood mononuclear cells[13]. Monocytes transform into macrophages and DCs during inflammation[19-23]. It has been reported that the increase in activated monocytes [human leukocyte antigen (HLA)-DRhighCD68+cells] in the liver is related to disease progression[24]. Chemokine (C-C motif) ligand 15 (CCL15) recruits CCR1+CD14+monocytes to the edge of HCC tissue. High expression of CCL15 is associated with poor clinical prognosis. CCR1+CD14+monocytes suppress antitumor immunity, facilitate tumor metastasis and promote tumor cell proliferation and invasion[25]. Blocking CCL2/CCR2-mediated macrophage accumulation has been proposed as a treatment strategy for HCC[26-29].
Myeloid-derived suppressor cells (MDSCs) are a heterogeneous population of myeloid progenitors and immature myeloid cells[30]. MDSCs in the tumor microenvironment, which are essential for tumor progression and are effective inhibitors of natural killer (NK) cells in HCC patients, play an important role in immunosuppression[31-33]. In humans, MDSCs usually express CD11b and CD33 and have low or no expression of HLA-DR. Monocyte-derived MDSCs (mMDSCs) express CD14, while granulocytederived MDSCs named gMDSCs express CD15 and CD66b[34]. MDSCs inhibit T and NK-cell proliferation and have an inhibitory effect on conventional NK cells in a TIGIT-dependent manner. Compared with conventional NK cells, adaptive NK cells express lower levels of TIGIT and resist the inhibitory effect of MDSCs[35]. MDSCs express CD155, which is induced by reactive oxygen species, and inhibit NK-cell functionviathe CD155-TIGIT interaction[35]. Compared with normal liver tissues, hepatoma tissues are rich in granulocytes and mononuclear cells. Mononuclear cells and MDSCs suppress the CD8+T-cell response[36].
S100A9 is a marker that can be used to distinguish MDSCs from monocytes[37]. Studies have identified CD14+HLA-DRlow/negmonocytes as MDSCs in HCC patients. The frequency of CD14+HLA-DRlow/negMDSCs in the peripheral blood and ascites fluid of these patients is increased. CD14+HLA-DRlow/negMDSCs cannot induce T-cell proliferation. MDSCs induce FoxP3+regulatory T (Treg) cell proliferation,while CD14+HLA-DR+monocytes induce Th17-cell proliferation and IL-17 secretion[33]. MDSCs induce Treg cell generationviaall-trans retinoic acid[38]. In human HCC, the fibrotic tissue around the tumor is rich in mMDSCs, which are significantly associated with a low survival rate[26].
DCs and HCC
DCs are a class of bone marrow-derived cells in the blood, epithelia and lymphoid tissues and are the most powerful professional APCs[39]. DCs participate in the regulation of innate and adaptive immunity[40]. DCs have antigen delivery capabilities, which make them attractive carriers for therapeutic tumor vaccines and platforms for vaccine development[41]. Human DCs are usually induced from monocytes isolated from peripheral blood mononuclear cells by M-CSF and IL-4 stimulation and develop into mature DCs (mDCs) after being loaded with antigens[42].
Human DCs express CD45 and HLA-DR and are divided into different populations according to CD11c, CD123 and IL-3Rα expression. Myeloid DCs (mDCs) are important antigen-presenting cells and express high levels of CD11c but low levels of CD123. Plasmacytoid DCs (pDCs) express high levels of CD11c, CD123, Toll-like receptor (TLR)-7 and TLR-9[43,44]. In normal liver tissues, CD141+hepatic DCs express CLEC9A, ILT3 and ILT4, but cell percentages and functions are damaged in hepatocellular carcinoma[45-47]. Evidence indicates that DCs have high antitumor and cytotoxic activity against HCC cellsin vitroandin vivo[48]. DCs inhibit the growth of HCC cells by loading LCSC antigens[49].
Peptide-pulsed DCs present antigens to naive T cells, thereby activating and inducing naive T cells to become antigen-specific cytotoxic T lymphocytes (CTLs) to kill tumor cells[50]. DCs incubated with epithelial adhesion molecule peptides have a significant inhibitory effect on tumor growth[51]. DC precursors that are sensitized by dexamethasone can effectively trigger the major histocompatibility complex class I (MHC I)-restricted CTL response, allowing DCs to make full use of secondary antigen peptides, thereby maximizing the specificity of the HCC immune response[52].
Macrophages and HCC
Macrophages originating in the liver are generally described as Kupffer cells (KCs)[53]. KCs express MHC class II and have varying levels of costimulatory markers, such as CD40, CD80, and CD86, and express the inhibitory marker Z39Ig[20,54]. The activation of STAT3 is involved in the differentiation of monocytes into macrophages[55]. Although tumor-infiltrating macrophages are generally believed to be derived from circulating monocytes, emerging evidence suggests that tissue macrophages can be maintained through self-renewal[56]. Liver resident macrophages are established by progenitor cells derived from the fetal liver and are maintained by self-proliferation and monocyte migration[57,58].KCs in the liver are negatively correlated with patient prognosis by inhibiting T-cell antitumor functions[59].
Tumor-associated macrophages (TAMs) are the most abundant immune cells in the tumor microenvironment and play a key role in immunosuppression[60]. TAMs are important in tumor-related inflammation and can be polarized into M1 or M2 phenotypes[61]. M1 macrophages are induced by LPS and IFN-γ; M2 macrophages are induced by IL-4 and IL-13[62]. It is generally believed that M1 TAMs have antitumor functions, while M2 TAMs promote tumor proliferation[63]. Studies have shown that IL-10 is involved in the polarization of TAMs to the M2 phenotype[64]. CD68 and CD163 are two popular TAM cell surface markers[65,66]. It has been reported that CD68+HLA-DR+TAMs enhance HCC proliferation through the NFκB/FAK pathway[67].
TAMs induce CD8+T-cell exhaustionviathe PD-1/PD-L1 and Tim3signaling pathways in the tumor microenvironment[68,69]. CCR2+TAMs accumulate and express the inflammatory marker S100A9 at the border of highly vascularized HCC, while CD163+TAMs accumulate in the center area of HCC tissue[70]. Silencing SIRT4 in TAMs regulates macrophage activation and significantly promotes HCC cell growth[71]. Downregulation of SIRT4 expression is related to an increase in macrophage infiltration in HCC tissues and a high ratio of M2/M1 macrophages[72]. The miR-148B/CSF1 pathway regulates the entry of TAMs into HCC tissue. A lack of miR-148B induces CSF1 expression and macrophage infiltration, thereby promoting liver cancer metastasis and poor prognosis[73].
THE REGULATORY FUNCTION OF HCC ON THE MPS
Studies have shown that the tumor environment can trigger early activation of monocytes in the surrounding tumor tissues[74,75]. Tumor environmental factors induce monocytes to express PD-L1 transiently in the early stage, activate monocytes, and induce PD-L1 expression in an autocrine cytokine-dependent manner[75]. HCC cells promote monocyte-to-macrophage differentiationviathe PKM2 pathway[76]. In addition, monocytes in human peripheral blood differentiate into immature dendritic cells[77]. Monocyte-derived TNF-α works synergistically with tumor-derived soluble factors to induce neutrophils to produce the metastasis-promoting factor OSM. Tumor-infiltrating monocytes mediate the production of CXCL2 and CXCL8 through the PFKFB3/NFκB signaling pathway. The level of PFKFB3 and the production of CXCL2/CXCL8 in monocytes are positively correlated with the infiltration of OSM-producing neutrophils in human HCC tissues[78].
The tumor microenvironment inhibits DC maturation and activation, leading to an immunosuppressive phenotype and function[79]. In addition, hepatoma cells recruit immunosuppressive DCs and MDSCs to suppress CD8+T cells, leading to tumor escape from immune surveillance[80]. Tumorderived exosomes impair the differentiation and maturation of DCs through the IL-6/STAT3 signaling pathway, which prevents myeloid precursor cells from differentiating into CD11c+DCs and induces apoptosis, thereby reducing T-cell activation and mediating immunosuppression[81,82].
The proliferation of tumor-infiltrated macrophages is more powerful than that of macrophages in nontumor tissues. Increased levels of macrophage proliferation are positively correlated with the density of macrophages in the tumor and the poor prognosis of HCC patients. Proliferating macrophages can be induced by small soluble adenosine, which is derived from tumor cells. GM-CSF released by tumors stimulates macrophages to express the A2A receptor, which then coordinates with adenosine to cause HCC-infiltrating macrophage proliferation[83]. Mitochondrial nucleoid structural changes lead to the release of mitochondrial DNA (mtDNA) into the cytoplasm, which is considered to be mtDNA stress, thereby regulating innate immunity[84].
CSF-1 and CCL2 in the tumor microenvironment polarize macrophages to the M2 phenotype[85,86].Crosstalk between macrophages and liver tumor cells can also affect the polarization of TAMs through the Wnt/β-catenin signaling pathway[87]. Compared to those in M1 cells, β-catenin, c-Myc and Axin2 are more enriched in M2 TAMs. It is believed that the c-Myc signaling pathway promotes the polarization of TAMs toward the M2 phenotype[88]. Neurotensin-induced IL-8 polarizes TAMs to the M2 phenotype and promotes epithelial-mesenchymal (EMT) in HCC through the MAPK and NFκB pathways[89]. In addition, hypoxia-inducible factors 1α (HIF-1α) and HIF-2α cause TAM polarization through hypoxia in the tumor microenvironment, leading to HCC malignancy[90,91]. Hypoxia-induced high mobility group protein box 1 may affect tumor progression by regulating TAMs, leading to poor clinical prognosis[92].
CONCLUSION
Components of the MPS have important but distinct roles in HCC. The MPS is associated with specific cell phenotypes and functions in the HCC microenvironment. Monocytes are progenitors of DCs and macrophages, which play an important role in inhibiting the immune response. In addition, MDSCs are a heterogeneous population composed of immature myeloid cells and myeloid progenitor cells.Increasing evidence suggests that tumor progression is associated with the accumulation of MDSCs,which cause local and systemic immunosuppression[93]. Furthermore, increased accumulation of MDSCs is associated with early tumor recurrence and is considered to be one of the predictors of poor prognosis in HCC patients who undergo clinical treatment[94]. Cytokine-induced killer (CIK) cells include a wide variety of T-cell receptor-specific effector cells, which are a mixed cell population. CIK cells have cytotoxic activity against tumor cells, which is not MHC-restricted. Studies have shown that adjuvant immunotherapy for HCC using CIK cells can reduce tumor recurrence and improve overall survival[95]. MDSCs inhibit CIK cell lysisviaARG1 and iNOS.
Human DCs are usually induced from monocytes isolated from peripheral blood. DCs have antigen delivery abilities, which make them attractive carriers for therapeutic tumor vaccines and platforms for vaccine development. In normal liver tissues, hepatic DCs have powerful antigen presentation functions and immunomodulatory abilities, but their percentages and functions are damaged in hepatocellular carcinoma[45]. Monocytes act as precursor cells that differentiate into DCs and macrophages under normal physiological conditions. The following are questions that remain to be investigated. Will monocyte impairments be inherited by DCs and macrophages? Will DCs and macrophages acquire immunosuppressive phenotypes and functions in HCC after differentiation?
Although the majority of previous studies have focused on individual MPS components, interactions and cooperation among various MPS components are common[96-98]. For instance, KCs are the main immune cells in liver tissues and play a key role in DC recruitment to the liver[97]. pDC survival and IFN-α production are affected by IL-10 and TNF-α secretion by monocytes. MDSCs produce increased levels of IL-10, which leads to pDC apoptosis. How do MDSCs interact with pDCs in the HCC microenvironment? Additionally, monocyte- or macrophage-derived IL-15 and pDC-derived IFN-α synergistically stimulate IFN-γ production in NK cells during HCV infection[96]. Does this interaction occur in HCC? Other forms of cooperation between different MPS components remain to be analyzed.
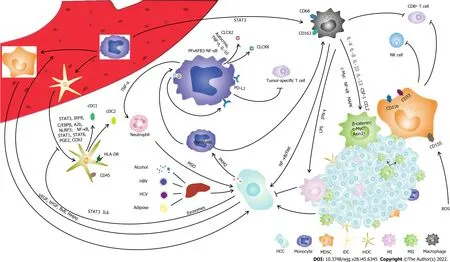
Figure 1 The interaction of mononuclear phagocyte system and hepatocellular carcinoma in the tumor microenvironment. (1) Activated monocytes secrete TNF-α and IL-10 to increases the expression of PD-L1 on the surface of monocytes, inhibiting the function of Tumor-specific T cell. TNF-α, CXCL2 and CXCL8 produced by monocytes stimulate neutrophils to produce MSO which promote tumor development; (2) Tumor-derived growth factors stimulate myeloid-derived suppressor cell differentiation through STAT3, IRF8, A2B, NLRP3 pathways and PEG2, COX2 produced by tumor stroma stimulate myeloid-derived suppressor cell differentiation through NK-κB, STAT1, STAT6 pathways. Myeloid-derived suppressor cell promotes tumor progression by producing VEGF, BV8 and MMP9. Myeloid-derived suppressor cells inhibits the function of NK cells and CD8+ T cells. Reactive oxygen species induce the expression of CD155 on myeloid-derived suppressor cells inhibiting natural Killer cell function; (3) The exosomes PKM2 produced by hepatocellular carcinoma in tumor microenvironment mediate the production of CXCL2 and CXCL8 through the PFKFB3-NF-κB signaling pathway; (4) Monocytes in peripheral blood mononuclear cells differentiate into macrophages through STAT3 pathway. LPS and IFN-γ stimulated tumor-associated macrophages polarizing into M1 phenotype, but IL-4, IL-8, IL-10, IL-13, CSF-1 and CCL2 stimulated tumor-associated macrophages polarizing into M2 phenotype through c-Mcy, NFκB and MAPK signaling pathway; and (5) Monocytes in peripheral blood mononuclear cells can be differentiated into immature dendritic cell, loaded with antigen to produce mature dendritic cell. Tumor-derived exosomes impair the activation and maturation of dendritic cells through IL-6 STAT3 signaling pathway. HCC: Hepatocellular carcinoma; MDSC: Myeloid-derived suppressor cells; iDC: Immature dendritic cell; mDC: Mature dendritic cell; M1: M1-type macrophages; M2: M2-type macrophages; ROS: Reactive oxygen species.
In this review, we described the impact of HCC on each component of the MPS and the immune function of the MPS in HCC development (Figure 1). Monocytes are precursors of DCs and macrophages, which play an important role in the regulation of immune function in HCC. Through crosstalk with hepatoma cells or regulation of the tumor microenvironment, monocytes are capable of producing high levels of IL-10, resulting in an inhibitory microenvironment. DCs can suppress the stimulation of T cells and inhibit efficient antitumor T-cell functions. TAMs are the most abundant immune cells in the tumor microenvironment and play a key role in immunosuppression. The tumor microenvironment inhibits DC maturation and activation, leading to an immunosuppressive phenotype and function. The HCC microenvironment remodels MPS cell phenotypes and cytokine production and impairs MPS components in HCC.
FOOTNOTES
Author contributions:Shan GY and Wang S collected the information; Cheng JY drew and modified the illustrations;Qiao DR wrote the paper; Li HJ and Yan WQ revised the paper.
Supported byNational Natural Science Foundation of China, No 81970529; and The Natural Science Foundation of Jilin Province, No. 20200201387JC.
Conflict-of-interest statement:All the authors report no relevant conflicts of interest for this article.
Open-Access:This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BYNC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is noncommercial. See: https://creativecommons.org/Licenses/by-nc/4.0/
Country/Territory of origin:China
ORCID number:Duan-Rui Qiao 0000-0001-6875-2143; Guan-Yue Shan 0000-0002-3380-9418; Shuai Wang 0000-0003-4659-3676; Jun-Ya Cheng 0000-0002-6161-3244; Wei-Qun Yan 0000-0002-4527-1301; Hai-Jun Li 0000-0002-2515-3076.
S-Editor:Liu GL
L-Editor:A
P-Editor:Liu GL
 World Journal of Gastroenterology2022年45期
World Journal of Gastroenterology2022年45期
- World Journal of Gastroenterology的其它文章
- Rifabutin as salvage therapy for Helicobacter pylori eradication: Cornerstones and novelties
- Germline BRCA2 variants in advanced pancreatic acinar cell carcinoma: A case report and review of literature
- Meta-analysis on the epidemiology of gastroesophageal reflux disease in China
- Endoscopic mucosal resection-precutting vs conventional endoscopic mucosal resection for sessile colorectal polyps sized 10-20 mm
- Best therapy for the easiest to treat hepatitis C virus genotype 1b-infected patients
- Deep learning based radiomics for gastrointestinal cancer diagnosis and treatment: A minireview