
热应激对妊娠后期奶牛和犊牛的影响及其调控措施研究进展
2022-12-08 12:41伍梓斌佘远航刘德武
动物营养学报 2022年11期
伍梓斌 赵 超 佘远航 刘德武
(华南农业大学动物科学学院,广州 510642)
热应激是指奶牛在高温、高湿环境下,奶牛自身产热超过自身散热能力,进而导致机体所作出的非特异性生理反应的总和,在研究中常用温湿度指数(temperature-humidity index,THI)对奶牛是否产生热应激进行评价[1]。热应激不但会对奶牛的直肠温度、呼吸频率等生理指标以及血液的生化指标产生影响[2],还会影响奶牛的躺卧行为以及肠道微生物区系的多样性[3]。尤其在我国南方地区,每年热应激不仅给畜牧业带来了巨大的经济损失,而且在一定程度上制约了奶牛养殖业的持续发展[4]。众所周知,妊娠后期是胎儿生长发育的重要时期。有研究表明,胎儿在妊娠的最后2个月积累大约60%的出生体重[5]。此外,妊娠后期也是奶牛乳腺上皮细胞发育的关键期,热应激会影响妊娠后期奶牛乳腺发育进而对奶牛下一泌乳周期的产奶量产生影响[6]。如果妊娠后期受到热应激,不仅会损害奶牛的生产力,还会对其后代产生负面的影响。鉴于妊娠后期对奶牛和新生犊牛的重要性,本文围绕着妊娠后期热应激对奶牛以及其后代影响的相关研究进行综述,旨在为妊娠后期奶牛的管理提供指导,为提高牛群生产性能和健康水平提供依据。
1 热应激对妊娠后期奶牛的影响
1.1 对妊娠后期奶牛乳腺发育的影响
热应激影响妊娠后期奶牛的乳腺发育。有研究表明,热应激会增加乳腺组织中抗凋亡热应激蛋白的表达,并使参与细胞生长、促进细胞凋亡等相关基因的表达下调,同时,热应激会阻碍乳腺上皮细胞合成蛋白,并造成乳腺退化[7]。另外,热应激还通过调节激素的分泌和蛋白质的合成直接或间接影响乳腺的功能[8],如催乳素(PRL)作为一种细胞因子,可以通过抑制脂肪组织中的脂蛋白脂肪酶参与脂肪代谢调控[9]。Do Amaral等[10]试验结果表明,妊娠后期受到热应激会引起奶牛血液中PRL含量升高,进而抑制了脂肪组织中脂蛋白脂肪酶活性,降低了脂质水解及游离脂肪酸(NEFA)含量。Zarzyńska等[11]研究发现,PRL可通过下调胰岛素样生长因子结合蛋白-5(IGFBP-5)基因的表达,优化生长因子胰岛素样生长因子-1(IGF-1)的作用,从而在体外降低乳腺细胞凋亡。也有研究表明,PRL在乳腺发育、乳汁分泌和乳蛋白基因表达中起着重要的调节作用,而奶牛发生热应激会造成血液中PRL含量变化,从而影响奶牛的泌乳性能[12]。除此之外,Collier等[13]发现在妊娠后期受到热应激的奶牛不仅能增强热休克蛋白(HSP)基因表达,而且能降低血液中雌激素含量,而HSP可作为分子伴侣协助蛋白质折叠,并通过干扰主要的细胞凋亡途径来抑制细胞凋亡;雌激素可以促进乳腺上皮细胞的自噬和细胞增殖[14]。因此,热应激可以通过增加HSP的合成和降低雌激素的含量,从而抑制乳腺上皮细胞的凋亡。
干乳期是奶牛乳腺二次发育的关键期,而奶牛发生热应激不仅不利于乳腺上皮细胞的增殖,还会对其乳腺的微观结构产生损害,进而对奶牛的下一个泌乳阶段造成不良影响。已有研究表明,与缓解热应激组相比,热应激组奶牛的产奶量平均下降约4.5 kg/d(表1)。Dado-Senn等[15]研究表明,与缓解热应激组相比,热应激组奶牛的乳腺中乳腺泡数量减少了15.4%,乳腺结缔组织比例增加了约7.9%,这表明乳腺上皮细胞的数量较低。因此,热应激可通过减少乳腺泡的数量从而导致乳汁合成与储存能力降低[16]。

表1 妊娠后期热应激和缓解热应激对奶牛自身产奶量的影响
1.2 对妊娠后期奶牛营养摄入和代谢的影响
妊娠后期奶牛要满足胎儿迅速生长发育、泌乳和保持自身体重3方面的营养需要,因此,充足的能量供应对满足机体的正常需求是必不可少的。然而,热应激会导致奶牛产前采食量下降,进而对奶牛的营养代谢、血液生化指标和生产性能产生影响[22]。在热应激条件下,奶牛干物质采食量不足,会导致奶牛处于能量负平衡而引起牛奶合成降低[23],改变奶牛体内参与养分合成与分解的激素水平[24]。Lamp等[25]研究发现,妊娠后期受到热应激的奶牛采食量下降约50%,采食量的下降会导致能量摄入减少,进而使热应激组奶牛产前增重低于缓解热应激组[26]。另有研究表明,尽管采食量较低,但是热应激组奶牛在分娩前其葡萄糖、NEFA[27]和胰岛素的循环浓度无显著差异。Tao等[26]对热应激组和缓解热应激组奶牛进行了葡萄糖耐量试验(GTT)和胰岛素刺激试验(IC),发现在分娩前2周其葡萄糖或胰岛素反应效率没有观察到差异。这说明分娩前采食量降低对葡萄糖或胰岛素浓度的影响不显著。
但有报道指出,热应激会影响奶牛分娩后血浆中葡萄糖、胰岛素和NEFA浓度。Tao等[26]试验结果表明,热应激降低了奶牛分娩后血浆中NEFA浓度,增加了血浆中葡萄糖和胰岛素浓度。Do Amaral等[28]研究发现,缓解热应激组奶牛的乳腺和肝组织在分娩后有更高比例的预成型脂肪酸,这与缓解热应激组奶牛产后血浆中胰岛素浓度低于热应激组奶牛相一致,因为胰岛素是一种能够促进脂肪合成的抗脂代谢因子。同时,热应激还能激活奶牛丙氨酸循环,但抑制了乳酸循环(图1),阻止了骨骼肌脂肪酸氧化的增加,并利用增加的丙酮酸进行糖异生,使得葡萄糖的利用程度更高[29]。Shahzad等[30]研究报道,热应激还会下调与脂肪酸氧化相关基因的表达,这证实了上述的观点。然而,关于妊娠后期受到热应激后对营养代谢的影响是否会导致下一次泌乳性能受损的根本原因尚不清楚,有必要继续深入研究。

图1 热应激对妊娠后期奶牛体内丙氨酸循环和乳酸循环的影响
1.3 对妊娠后期奶牛免疫功能的影响
热应激不仅会影响妊娠后期奶牛的先天性免疫,也会对后天的获得性免疫产生影响。在发生热应激时,奶牛体内多种免疫因子的含量均会变化,进而影响免疫细胞的生长发育,导致机体的免疫功能下降[31]。同时,Thompson等[32]研究发现,热应激组奶牛血液中循环中性粒细胞数量低于缓解热应激组。Do Amaral等[18]研究发现,热应激对奶牛在产犊前中性粒细胞呼吸爆发功能没有影响,但是在产犊后第20天时,其中性粒细胞呼吸爆发功能低于缓解热应激组。因此,我们推测热应激可能对妊娠后期奶牛的先天性免疫功能影响存在一定的延续效应。
获得性免疫与先天性免疫不同,获得性免疫会直接受到妊娠后期热应激的影响。体液免疫功能主要由B细胞分化为浆细胞并分泌抗体来实现的,热应激会使奶牛体内皮质酮含量处于较高水平,进而诱导细胞发生长时间的溶解反应,使得奶牛体内免疫球蛋白合成量降低,最终导致免疫功能下降[33]。此外,Adin等[17]发现热应激还会降低奶牛初乳中的免疫球蛋白G(IgG)含量,而初乳中IgG的含量取决于通过特定的运输机制从血液流过乳腺屏障的含量。这表明,热应激阻碍了血液中IgG向乳腺的转移,损害了奶牛的体液免疫,但是具体的机制还有待进一步研究。然而,Monteiro等[19]和Skibiel等[34]的研究结果与上述结果不一致,这可能是因为两者进行试验的奶牛之间存在个体差异造成的。此外,热应激会影响妊娠后期奶牛的获得性免疫。Lacetera等[35]发现在夏季产犊的奶牛,其外周血单核细胞(PBMC)的数量减少。同样地,与缓解热应激组相比,从热应激组奶牛中分离出来的PBMC不利于其细胞增殖和肿瘤坏死因子-α(TNF-α)的产生,因此,妊娠后期受到热应激会对奶牛获得性免疫产生负面影响[36]。热应激不仅能够直接降低细胞免疫和体液免疫的功能,而且还能间接地通过其他的活性物质影响机体的细胞免疫和体液免疫。
有研究表明,当机体遭受热应激时,体内通过大脑对免疫功能的调控以及肾上腺皮质激素和交感神经的免疫修饰来抑制体内的免疫器官、组织和细胞功能[37]。另有研究表明,热应激会激活下丘脑-垂体-肾上腺皮质(HPA)轴,使得糖皮质激素水平上升,进而抑制免疫细胞的功能[38]。妊娠后期热应激对奶牛免疫功能的影响见图2。有研究表明,细胞因子会对热应激诱导糖皮质激素的作用产生影响[39]。同时,辅助性T细胞(Th)1会激活细胞免疫和炎症反应,Th2则调节体液免疫和促进抗炎反应[40]。糖皮质激素通过抑制白细胞介素-12(IL-12)影响Th1和Th2的平衡,导致体内细胞免疫受到抑制,向Th2介导的体液免疫优先转移[41]。Caroprese等[42]研究表明,热应激使奶牛血液皮质醇浓度升高,而皮质醇能够与DNA结合,从而抑制T细胞活化和细胞因子产生的相关基因表达。因此,妊娠后期热应激会损害细胞免疫反应。然而,Tao等[43]报道热应激使奶牛的血液皮质醇浓度有所降低,这可能是因为长时间的热应激能够降低HPA轴对热环境的敏感性,进而引起皮质醇浓度的降低。因此,维持Th1和Th2平衡可能是减少热应激对妊娠后期奶牛免疫功能影响的关键因素。
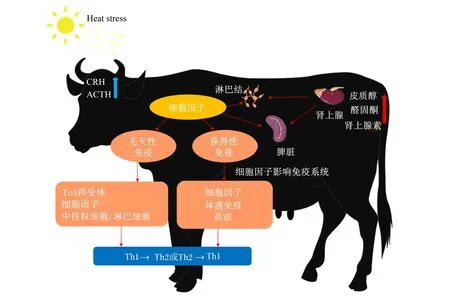
Heat stress:热应激;CRH:促肾上腺皮质激素释放激素 corticotropin releasing hormone;ACTH:促肾上腺皮质激素 adrenocorticotropic hormone;Th1:辅助性T细胞1 helper T cell 1;Th2:辅助性T细胞2 helper T cell 2。
1.4 对妊娠后期奶牛健康的影响
热应激会影响妊娠后期奶牛的生理机能,进而对奶牛的健康产生影响变化,直接导致奶牛呼吸频率以及心率加快、大量出汗以及精神萎靡,给妊娠后期奶牛的健康状态带来严重影响[44]。热应激会使妊娠后期奶牛的躺卧时间减少,进而使蹄病发病率增加[45]。此外,热应激还会导致妊娠后期奶牛乳房炎发病率的增加,发生热应激的奶牛体内免疫细胞的数量和活性均会显著下降,使得奶牛的免疫能力降低而更易被病原菌感染,而且夏季牧场采用“风扇+喷淋”的热应激防控措施增加了环境湿度,给病原菌的滋生提供了更适宜的环境,最终导致奶牛患乳房炎的发病率急剧增长[46]。
2 妊娠后期热应激对初生犊牛的影响
2.1 对初生犊牛生长性能的影响
妊娠后期热应激不仅能够损害奶牛的生理机能,还可能对后代的初生重产生负面影响。有研究表明,妊娠后期是胎儿生长发育的关键时期,胎儿增重的80%~90%是在此阶段完成[47]。在热应激环境下,胎儿的体温相对于母体更高[48]。胎儿体温升高会影响犊牛的免疫功能,导致犊牛在出生后对疾病的抵抗力下降,进而影响胎儿的生长发育[49]。热应激会减少胎盘重量,并缩短妊娠时间[50]。Monteiro等[19]、Almoosavi等[21]和Dahl等[51]发现热应激组奶牛子代生长性能受限,初生体重和断奶体重均较低。Dado-Senn等[52]试验表明,热应激组奶牛所产的犊牛初生体重平均减少了5.6%,断奶体重减少了8.1%。这种变化可能是因为高温环境下可降低动物的干物质采食量,干奶期奶牛营养不足影响了胎儿的发育,或热应激组奶牛体内循环的胎盘激素水平减少,从而影响了胎盘的正常功能[53]。此外,热应激还会影响奶牛的妊娠时间。热应激组奶牛妊娠期减少4~5 d[53-54],这也可能是导致犊牛初生重减轻的原因之一。
妊娠后期热应激还会影响犊牛的生长发育。犊牛阶段是奶牛生长的一个特殊生理,此阶段对环境变化的抵抗力较差。在热应激的环境下会影响犊牛的采食量,导致其生长发育缓慢甚至停滞,患病率增加,严重时可能会出现死亡[55]。Monteiro等[56]发现在断奶前阶段,热应激组和缓解热应激组犊牛具有相似的平均日增重(ADG),然而Laporta等[20]研究表明,热应激组犊牛断奶前的ADG更小。导致这种差异可能是因为热应激的缓解措施以及热应激的强度和持续时间不同所致。此外,妊娠后期热应激还会影响犊牛的成活率。Monteiro等[56]试验结果中热应激组犊牛的死亡率更高。同样,Laporta等[20]研究发现,热应激组犊牛从出生到动物离开畜群的时间比缓解热应激组犊牛缩短了356 d,热应激组的死胎率为3.8%,而缓解热应激组则无死胎发生。

表2 热应激和缓解热应激对妊娠后期奶牛后代初生体重和断奶体重的影响
2.2 对初生犊牛免疫功能的影响
奶牛妊娠后期受到热应激会改影响犊牛的免疫功能,包括犊牛的被动免疫和细胞免疫。首先,妊娠后期发生热应激会影响奶牛初乳成分,并通过初乳成分的改变影响犊牛的被动免疫。成功的被动免疫转移对新生犊牛的生存至关重要,而被动免疫转移取决于初乳中IgG含量以及犊牛吸收初乳中IgG的能力[60]。关于妊娠后期受到热应激对初乳IgG含量的影响说法不一,因此,初乳IgG含量似乎并不是影响犊牛被动免疫转移的主要因素。为了避免初乳质量对新生犊牛对初乳IgG吸收能力的影响,Almoosavi等[21]在所有犊牛均采用相同的混合初乳时,发现热应激组的犊牛其出生后第1天血清IgG含量较低。同样,Tao等[43]试验结果表明热应激组犊牛血液IgG含量减少了约32.9%,IgG的表观吸收效率降低了约42.9%。Monteiro等[19]和Almoosavi等[21]研究发现,热应激组犊牛的IgG吸收效率较低。这进一步证实了妊娠后期热应激通过降低犊牛吸收IgG的能力而使其被动免疫受损的观点。这可能是由于肠道中IgG吸收能力受损,在妊娠后期受到热应激加快了肠道闭合,从而导致IgG吸收的时间变短或阻碍了小肠细胞内吞,最终导致IgG吸收能力较低。
其次,妊娠后期发生热应激还会影响奶牛细胞炎症因子的分泌量,从而对犊牛的细胞免疫产生影响。机体相关免疫细胞分泌的细胞因子是细胞免疫系统关键的信号因子。细胞炎症因子分泌量与转录水平的高低是免疫功能正常与否的重要标志[61]。有研究表明,TNF-α是一种促炎细胞因子,能够激活中性粒细胞和淋巴细胞,而Toll样受体2(TLR2)是先天免疫系统中的免疫蛋白,可调节炎症细胞因子TNF-α等的表达[62],因此,妊娠后期受到热应激会减弱犊牛体内免疫屏障功能。PBMC是一类重要的免疫效应细胞,主要包括淋巴细胞和单核细胞。Tao等[43]发现妊娠后期受到热应激的犊牛其血液中PBMC增殖较低,而PBMC的数量降低则表明机体免疫功能下降。Monteiro等[19]试验表明,热应激组犊牛第42天全血T淋巴细胞发育和增殖能力比缓解热应激组更弱,这说明热应激对犊牛的免疫系统发育产生不良影响。Ahmed等[57]研究发现,胸腺是T淋巴细胞分化成熟的重要场所,主要参与机体内细胞免疫,与缓解热应激组犊牛相比,热应激组犊牛除了初生体重显著降低,其免疫器官胸腺的重量减少了约22%,肝脏的重量减少了约19.4%。因此,妊娠后期受到热应激后胎儿器官发育受损可能会导致犊牛细胞介导免疫的改变。综上所述,奶牛妊娠后期受到热应激会通过降低犊牛吸收IgG的能力和损害胎儿的免疫器官,从而对犊牛的免疫功能产生负面影响。
2.3 对初生犊牛新陈代谢的影响
妊娠后期热应激还会影响犊牛的代谢反应。妊娠后期对初生犊牛的采食量影响不一,研究表明,妊娠后期热应激组和缓解热应激组的犊牛之间的采食量没有差异,或热应激组的犊牛的采食量有所减少。虽然犊牛的采食量没有差异,但Bell等[50]的试验结果中热应激组犊牛需要人工灌喂的次数大约是缓解热应激组犊牛的2倍,因此我们猜测可能是由于产前受到热应激使犊牛的新陈代谢发生改变,从而降低了犊牛的食欲。
妊娠后期受到热应激会通过影响母牛体内激素的分泌,进而使犊牛体内激素水平发生改变,最终影响犊牛的代谢反应。热应激的发生改变了体内碳水化合物的代谢,主要以胰岛素基础浓度的改变以及胰岛素对葡萄糖变化的反应性改变为特征[63]。胰岛素在葡萄糖和脂肪酸代谢中起到关键作用,在妊娠后期受到热应激的犊牛胰岛素的分泌和反应会受到抑制。Guo等[58]研究发现,热应激组犊牛血浆胰岛素、PRL、IGF-1浓度较低,但是出生后2 h内犊牛血浆葡萄糖、NEFA或β-羟基丁酸(BHBA)浓度没有差异变化。Tao等[53]研究报道,在饲喂初乳前,热应激组犊牛与缓解热应激组犊牛血浆中循环胰岛素相似,但是当饲喂初乳后,热应激组犊牛在出生后第1周体内循环胰岛素浓度高于缓解热应激组。因此,我们推测热应激出生的犊牛在饲喂初乳后胰岛素反应增加可能是胰岛素分泌增强或胰岛素清除受损的结果。Tao等[64]研究发现,热应激组犊牛在GTT和IC期间葡萄糖的清除率较快,且断奶后其全身胰岛素反应更强[48]。虽然妊娠后期热应激奶牛会诱导外周组织的胰岛素抵抗,但改善了犊牛产后胰岛素非依赖性葡萄糖吸收和基础葡萄糖的摄取,因此这些结果初步证明了妊娠后期热应激改善了犊牛早期营养分配。然而,其具体的细胞机制有待进一步研究。
2.4 对初生犊牛健康的影响
初生犊牛的免疫系统机能尚未发育完善,抵抗力较差,易受外界环境影响。对环境温度变化最敏感,其适宜的环境温度为10~23 ℃,当温度高于23 ℃时就会出现热应激反应。研究表明,产前热应激奶牛所产犊牛的初生重和断奶重均显著低于产前采取降温措施奶牛所产的犊牛,而且产前发生热应激奶牛所产犊牛的死胎率显著高于低THI环境下的奶牛,热应激还会导致初生犊牛的疾病率、畸形率显著上升,进而导致犊牛的发育迟缓及成活率下降[56]。Uyeno等[65]研究发现,持续高温所引起的热应激会对犊牛瘤胃微生物区系产生显著影响,导致瘤胃中负责分解纤维素的纤维杆菌属菌的相对丰度降低,进而使奶牛对饲料的消化率降低。有研究表明,当热应激发生时,犊牛对IgG的吸收效率显著降低,导致犊牛体内抗体水平显著下降,免疫力降低,易被病原微生物感染,最终导致犊牛死亡率提高[43]。此外,热应激还会导致犊牛体内TNF-α表达过度,进而引起犊牛的肠道炎症,显著增加犊牛的腹泻发病率[36]。综上所述,妊娠后期受到热应激会降低犊牛对外界环境的抵抗力,导致其他疾病高发,从而影响犊牛健康。
3 缓解奶牛妊娠后期热应激的调控措施
妊娠后期热应激的产生对奶牛自身和犊牛都可产生较大负面影响,严重损害了奶牛的生产力。因此,在妊娠后期缓解热应激是提高奶牛生产效率的重要手段。现阶段缓解奶牛热应激主要通过环境、营养以及饲养管理3个方面进行调控[66]。
3.1 环境调控
适宜的环境条件是保证奶牛生产性能的关键性条件,因此,环境调控缓解热应激是一项首要工作。环境调控主要包括奶牛场的建设和物理降温2部分。首先在奶牛场的选址和建设方面,在南方湿热地区,奶牛场应建造在通风良好的地方,屋顶应采用钟楼式比较合适,做好外围护结构的隔热,采用浅色、光平外表面材料建设牛舍以减少太阳辐射热的吸收。在牛舍布局方面,牛舍要有足够的间距,东西朝向,同时牛场的周围应搭建遮阳棚或种植一些树木,从而起到一定的遮阳作用[67]。
其次,物理降温是奶牛场常用的缓解热应激的措施。奶牛场的物理降温主要分2种途径,分别是湿帘+风扇模式和喷淋+风扇模式。研究表明,湿帘+风扇模式最高可以使奶牛场的气温下降7.5 ℃,同时奶牛场的湿度增加也较少[68]。但是,目前大部分规模化牧场采用喷淋+风扇模式,该模式主要是通过风扇加快水分挥发带走热量,可以达到快速降低奶牛体温的目的,当牛舍温度高于27 ℃时,喷淋+风扇模式可使奶牛体温平均降低1.7 ℃,而对干奶期奶牛效果更显著,可使奶牛下一个泌乳周期产奶量增加3.5 kg/d[69],但是喷淋+风扇模式会使得奶牛场的湿度快速上升,这同样会对奶牛产生不利影响。因此,在建设奶牛场物理降温设施的时候应选用合适的喷嘴,从而达到改善喷淋系统的降温效果的同时又能减少奶牛场湿度的上升。
3.2 营养调控
3.2.1 合理调整饲粮的精粗比
热应激期间动物为减少机体增热和散热负担,会使机体减少营养摄入,其中能量摄入量减少尤为明显,所以应适当加大精料比例(60%~65%,但粗料中性洗涤纤维水平要达到18%以上)有利于减轻奶牛热应激的危害。陈志伟等[70]试验结果表明,通过提高饲粮中的蛋白质含量可以显著提高奶牛的产奶量和干物质采食量。脂肪是一种代谢能值高、热增耗小的营养物质,其一般不在瘤胃内发酵。因此,在热应激条件下适当增加脂肪类物质对缓解奶牛热应激具有一定作用[71]。Drackley等[72]试验结果表明,添加未包被脂肪补充剂能够使NEFA循环浓度升高,进而缓解了发生热应激时奶牛体内能量不足的问题。Moallem等[73]研究发现,在饲粮中添加1.5%脂肪酸钙可显著提高乳蛋白含量和饲料产奶效率。
3.2.2 添加剂的使用
在饲粮中添加一定量的非营养性抗热应激饲料添加剂可有效缓解热应激,提高奶牛生产性能。目前报道较多的抗热应激饲料添加剂主要包括矿物质类、维生素类、酶制剂类、微生态制剂和中草药类等[74]。维生素C有利于机体类固醇激素的合成,在奶牛饲粮中添加适量的维生素C能够降低血液中皮质醇浓度,同时还能有效缓解奶牛体温上升,并提高奶牛干物质采食量[75]。黎智峰[76]和赵春卉等[77]试验发现,饲粮添加烟酸铬和酵母铬,干乳期奶牛直肠温度、呼吸频率降低,利于缓解奶牛的热应激状态。Zhang等[78]试验结果发现,在奶牛饲粮中添加甜菜粕能使发生热应激期间的奶牛直肠温度和呼吸频率保持较低水平,同时还能有效增加奶牛的采食量和产奶量,有利于缓解奶牛的热应激。此外,在我国,中草药在生产上作为抗应激添加剂已经有了有力的理论支持。在夏季高温湿热环境下,饲粮中添加黄芪和柴胡能够显著提高热应激奶牛的干物质采食量和产奶量[79-80]。因此,我们可以通过添加剂的合理使用来达到缓解奶牛热应激的目的,从而提高奶牛产奶量,减轻热应激对奶牛的危害。
3.3 加强奶牛的日常管理
日常的饲养管理是保证营养配方实施的基础,奶牛场必须要注重日常管理,才能减少因热应激对奶牛造成的不良影响。首先,应该减少人对奶牛的应激,尽可能少调群并做好奶牛卧床管理。合理的调群可以使奶牛面临的应激降到最低,从而避免奶量损失,避免在热应激的环境下调群而造成过大的应激。其次,高温环境有利于病原菌的繁殖,特别是对大肠杆菌和金黄色葡萄球菌生物膜的生长有利[81]。因此,应该定期清理奶牛行走通道粪污、做好灭“四害”工作,防止奶牛感染虫媒病。此外,在热应激条件下,奶牛的生产能力受到严重影响,产奶量和乳成分会因高温而发生改变[82]。在生产中可以通过在牛舍中安装热应激检测设备(温湿度计、风速仪、热成像仪)来及时观测奶牛的牛体情况,减少奶牛处于热应激的时间,从而提高牧场的经济效益。最后,在热应激条件下,牛舍内必须要有足够的饮水槽,最好保证15~20头奶牛1个水槽为佳,并保证饮水不间断和水质洁净[83]。
4 小 结
热应激影响妊娠后期奶牛及犊牛的生产性能、代谢反应和健康。热应激可损害妊娠后期奶牛乳腺发育,降低下一胎次的产奶量,同时降低了奶牛和犊牛的免疫功能,从而对实际生产造成重大的经济损失。然而,针对妊娠后期热应激的缓解措施等相关文献相对较少,因此,牧场管理者未来应该要加强妊娠后期热应激的调控。另外,与妊娠后期热应激相关的表型差异是复杂的,涉及到多种机理还尚未清楚,需深入研究。
猜你喜欢
今日农业(2022年3期)2022-11-16
昆明医科大学学报(2021年10期)2021-12-02
今日农业(2021年7期)2021-07-28
今日农业(2021年10期)2021-07-28
现代畜牧科技(2021年4期)2021-07-21
现代畜牧科技(2021年4期)2021-07-21
小天使·一年级语数英综合(2019年12期)2019-01-13
中国生殖健康(2019年5期)2019-01-06
妈妈宝宝(2017年2期)2017-02-21
中国乳业(2016年4期)2016-11-07
