
不同干旱程度对枣树苗木水碳平衡和生物量分配的影响
2022-12-05 06:54:04张拥兵李俊鹏吕英忠
经济林研究 2022年3期
张拥兵,王 林,薛 浩,李俊鹏,吕英忠
(1.山西农业大学 果树研究所,山西 太谷 030815;2.山西农业大学 林学院,山西 太谷 030801)
枣树是我国传统的经济树种,因其环境适应能力强,尤其对干旱的适应能力较强,在黄土高原和西北地区有较大的种植面积[1],但目前对其抗旱的生理机制尚不完全了解,有研究表明枣树属深根性树种,因此对表层土壤缺水的适应能力较强[1],能够较多地利用土壤深层的水分;也有研究表明枣树渗透调节能力强,且在干旱条件下细胞膜稳定性强,且对过氧化的保护能力强[2],因此其对干旱有较好的适应能力。水分代谢和碳代谢是植物存活和生长的基础[3-4],现有研究表明水分输导系统和碳平衡维持能力是植物在干旱条件下存活的基础[3-5],但目前对这方面的研究较少,这也限制了对枣树应对干旱能力和生理机制的全面了解。
本研究拟以不同干旱程度下枣树大苗为研究对象,通过比较在不同干旱程度下的生长、生物量分配、水分状况、光合作用、非结构性碳含量,分析干旱胁迫程度对枣树水碳代谢和生长、生物量分配的影响,研究结果可以揭示枣树对干旱的适应能力,同时揭示枣树在不同干旱程度下的水碳平衡及对生长的影响。本研究关注的问题是:1)枣树在不同干旱程度下的生长和生物量分配;2)不同干旱程度对枣树水分代谢的影响;3)不同干旱程度对枣树碳摄取和非结构性碳分配的影响。
1 材料与方法
1.1 试验材料及试验设计
本研究在山西省太谷区山西农业大学果树研究所进行,于2020年3月中旬选取无病虫害生长基本一致的2年生冷白玉枣树苗定植到直径30 cm、高40 cm 的盆中,培养土采用1/2 园土+1/4 腐熟牛粪+1/4 沙组成。在2 个月后(2020年5月中旬),选取生长基本一致的苗木32 株,随机分为正常供水、轻度干旱、重度干旱和重度干旱4个处理,4 个处理体积含水量分别控制在25%~28%、17%~20%、12%~14%和6%~8%,使用TRME-T3 TDR 便携式土壤水分测定仪(IMKO,Ettlingen,Germany)测定,干旱处理开始后将苗木放置在避雨棚中。在处理4 个月后(2020年9月中旬),测定苗木的水势、导水损失率(Percentage loss of conductivity,PLC)、光合作用、株高、基径、当年生枣头长度、当年生枣头基径、二次枝长度、二次枝基径、叶面积、不同部位的干质量,并测定不同部位的非结构性碳(Non-structural carbohydrate,NSC)含量。
1.2 试验方法
1.2.1 水势和导水损失率测定
小枝凌晨和正午水势用PMS 600D(PMS Albany USA)测定,凌晨水势的测定时间为5:30—6:00,正午水势的测定时间为12:00—13:00,导水损失率的测定时间同正午水势,具体测定方法参照王林等[6-7]的方法进行。
1.2.2 光合速率测定
光合速率和蒸腾速率测定在2020年9月中旬的晴天进行,用Li-6400 光合测定系统(LI-COR,Lincoln,USA),设定光强为1500 μmol·m-2·s-1,该光照强度在枣树的光饱和点之上[1]。
1.2.3 枝条和叶片形态测定
株高、当年生枣头长度、二次枝长度用米尺测定,苗木基径、当年生枣头基径、二次枝基径用游标卡尺测定;叶面积选取树冠中上部向阳面叶片,用YMJ-C 型叶面积仪(托普,杭州,中国)测定。
1.2.4 生物量测定
将苗木收获,每株按叶片、枝条、主干、粗根(≥2 mm)、细根(<2 mm)分开,在75 ℃条件下烘干72 h 后称取其质量。
1.2.5 NSC 测定
取收获烘干后的材料,枝条、粗根用小刀仔细将木质部和韧皮部分开,分别粉碎后过100目筛,用于NSC 含量测定,可溶性糖和淀粉含量测定方法参照Audrey 等[8]的方法进行,NSC 含量为可溶性糖含量和淀粉含量之和。
1.2.6 数据处理
采用SPSS 22.0 软件进行统计分析,不同处理间采用单因素ANOVA 分析(One-way ANOVA),并用Duncan 检验进行多重比较,用Sigmaplot 10.0 软件作图,所有测定指标均有6 个以上重复。
2 结果与分析
2.1 干旱程度对枣树苗木水势、导水损失率的影响
随着干旱程度的增加,凌晨水势、正午水势均呈现下降趋势,除轻度干旱的凌晨水势和正午水势与正常供水处理间没有显著性差异外,其余不同处理间均存在显著性差异(P<0.05)。随着干旱程度的加重,导水损失率呈增加趋势,重度干旱和中度干旱下枣树枝条的导水损失率均显著高于正常供水处理(P<0.05),由正常供水的46.2%±7.72%增加到重度干旱的77.6%±4.06%,轻度干旱下枣树的枝条导水损失率略高于正常供水处理,但没有达到显著性差异水平(图1),这表明中度和重度干旱较大程度地影响了枣树苗木的水分状况,导水损失率增加进一步限制了水分长距离运输。
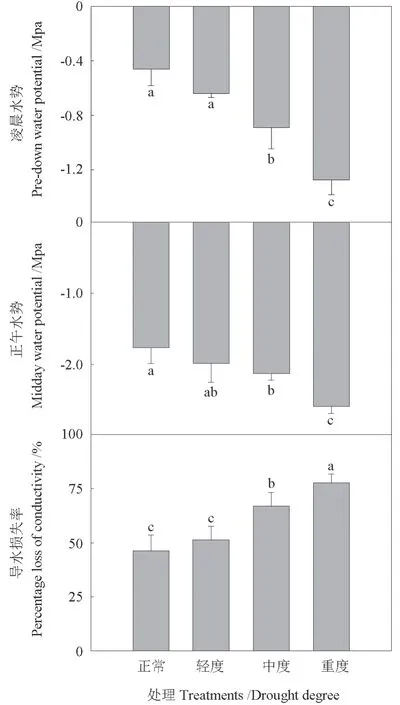
图1 不同干旱程度下枣树苗木的凌晨水势、正午水势、导水损失率Fig.1 Pre-down water potential, midday water potential and percentage loss of conductivity of jujube seedings under different draught treatments
2.2 干旱程度对枣树苗木光合作用的影响
随着干旱程度的增加,枣树苗木的光合速率和蒸腾速率均呈现下降趋势,轻度干旱处理枣树苗木的光合速率和正常供水处理间没有显著性差异,中度干旱和重度干旱枣树叶片光合速率均显著低于正常供水处理(P<0.05),中度干旱和重度干旱的光合速率分别下降27.2%和58.5%。中度干旱和重度干旱蒸腾速率较正常供水处理分别下降36.0%和58.4%。不同处理间的水分利用效率没有显著性差异(图2),随着干旱胁迫的加重略呈现先增加后降低的趋势,这表明中度和重度干旱较大程度地影响了枣树的光合作用。
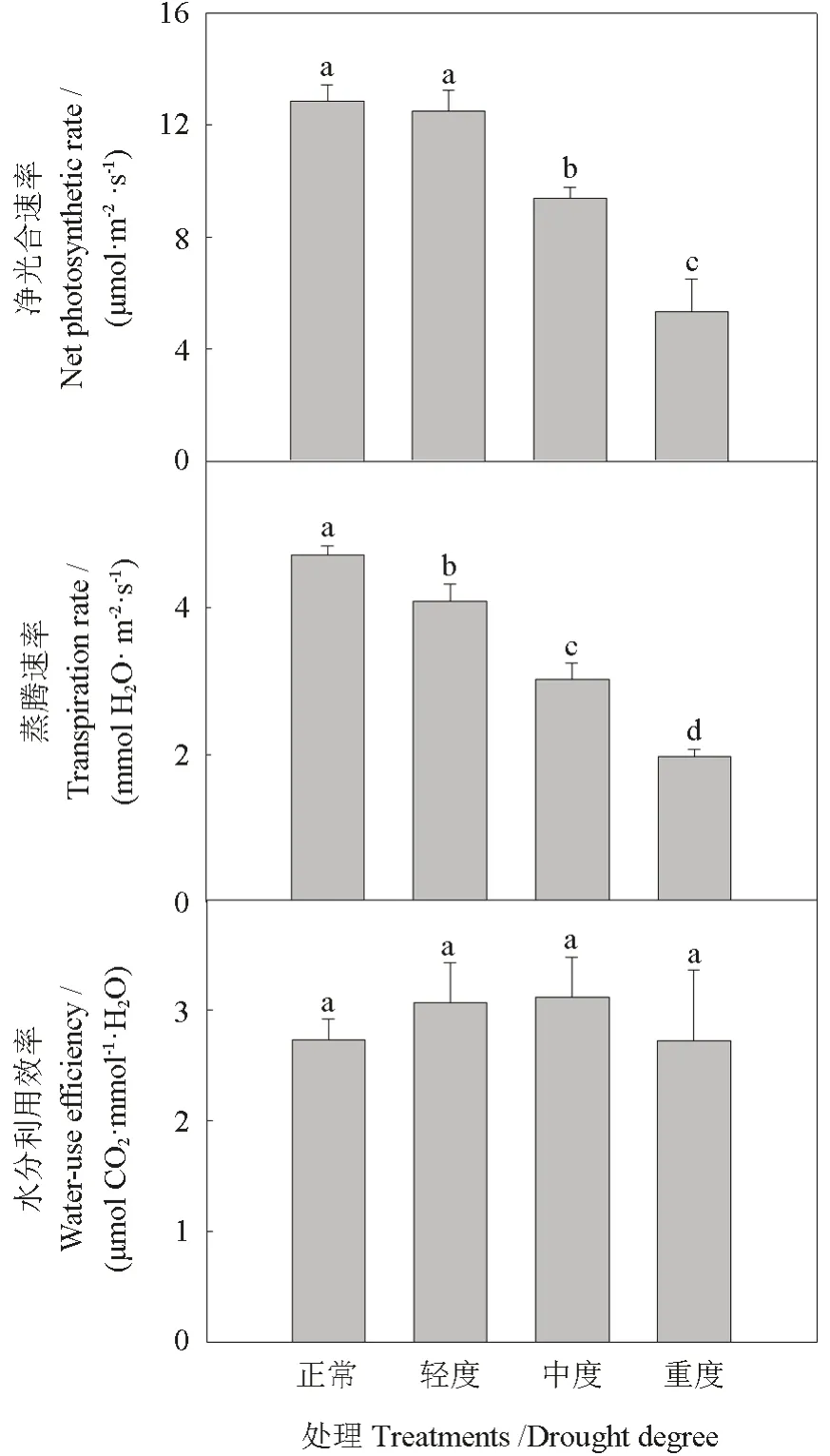
图2 不同干旱程度下枣树苗木的净光合速率、蒸腾速率、水分利用效率Fig.2 Net photosynthetic rate, transpiration rate and water-use efficiency of jujube seedings under different draught treatments
2.2 干旱程度对枣树苗木生长的影响
轻度干旱处理的株高较正常供水处理没有影响,中度和重度干旱显著降低了枣树苗木的株高(P<0.05),中度干旱和重度干旱处理的株高分别比对照低9.0 %和20.6 %。不同处理在苗木基径上没有显著性差异,重度干旱苗木基径略低,但没有显著性差异。重度干旱显著降低了当年生枣头的长度和基径(P<0.05),轻度干旱和中度干旱的当年生枣头长度和基径没有显著性差异。干旱处理对当年生二次枝的长度和基径没有影响。重度干旱显著降低了枣树苗木的叶面积(P<0.05),中度干旱叶面积也有一定程度的降低,但与正常供水处理没有显著性差异(表1),这表明重度干旱限制了枣树苗木的生长和叶面积,轻度和中度干旱影响程度较小。

表1 不同干旱程度下枣树苗木的生长(平均数±标准差)Table 1 Growth of jujube seedings under different draught treatments (mean ± SD)
2.3 干旱程度对枣树苗木生物量和生物量分配的影响
重度干旱处理显著降低了叶、枝条、主干、细根的生物量(P<0.05),但轻度和中度干旱处理与正常供水处理在叶、枝条和主干生物量上没有显著性差异。轻度干旱和中度干旱处理的粗根生物量显著高于重度干旱处理(P<0.05),轻度干旱和中度干旱处理的粗根生物量较正常供水处理也有一定程度地增加,重度干旱粗根生物量较正常供水处理有一定程度地下降,但均没有显著性差异。随着干旱程度的加重,细根生物量呈下降趋势,轻度、重度和重度干旱处理的细根质量较正常供水分别下降8%、12.5%和47.4%。重度干旱显著降低了枣树的总生物量(P<0.05),但轻度和中度干旱与正常供水处理间没有显著性差异。不同处理间在根冠比上没有显著性差异,随着干旱程度的加重,细根生物量占比呈现下降趋势,重度干旱处理的细根生物量显著低于正常供水处理(P<0.05)。随着干旱胁迫的加重,叶片的生物量占比略呈现下降趋势,但不同处理间没有达到显著性差异程度(表2)。这表明重度干旱对枣树的生物量积累造成了较大限制,同时随着干旱程度的较重,叶和细根的生物量占比逐渐降低。
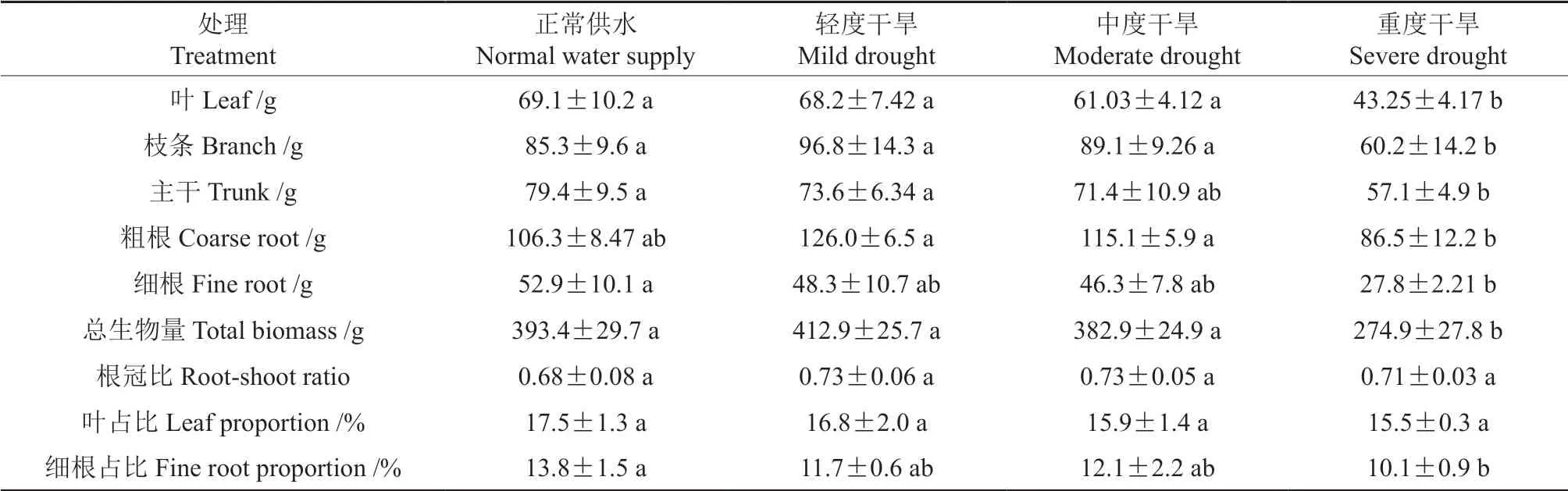
表2 不同干旱程度下枣树苗木的生物量和生物量分配(平均数±标准差)Table 2 Biomass and biomass distribution of jujube seedings under different draught treatments (mean ± SD)
2.4 不同干旱程度对枣树NSC 含量的影响
不同干旱程度下的叶片、枝条韧皮部的可溶性糖含量没有显著性差异,中度干旱处理的枝条木质部、粗根木质部可溶性糖含量显著高于对照(P<0.05),轻度干旱和重度干旱处理的枝条木质部、粗根木质部可溶性糖含量也略高于对照,但没有显著性差异。重度干旱处理显著降低了粗根韧皮部的可溶性糖含量。不同处理间在细根可溶性糖含量上没有显著性差异(图3)。

图3 不同干旱程度下枣树苗木的叶、枝条木质部、枝条韧皮部、粗根木质部、侧根韧皮部、细根可溶性糖含量(平均数±标准差)Fig.3 Soluble sugar content of leaves, branch xylem, branch phloem, coarse root xylem, coarse root phloem and fine root of jujube seedings under different draught treatments (mean ± SD)
轻度干旱和中度干旱处理的叶片和枝条韧皮部NSC 含量显著高于重度干旱处理(P<0.05),正常供水处理的叶和枝条韧皮部NSC 含量叶略高于重度干旱处理。中度干旱处理的粗根木质部NSC 含量显著高于正常供水处理(P<0.05),重度干旱处理的细根和粗根韧皮部NSC 含量显著低于正常供水处理(P<0.05),但轻度和中度干旱对细根和粗根韧皮部NSC 含量没有显著性影响(图4)。以上结果表明:轻度和中度干旱在一定程度上增加了枣树苗木多数部位的NSC 含量,重度干旱显著降低了叶片、粗根韧皮部和细根的NSC 含量。

图4 不同干旱程度下枣树苗木的叶、枝条木质部、枝条韧皮部、粗根木质部、侧根韧皮部、细根NSC 含量(平均数±标准差)Fig.4 NSC content of leaves, branch xylem, branch phloem, coarse root xylem, coarse root phloem and fine root of jujube seedings under different draught treatments (mean ± SD)
3 讨 论
3.1 不同干旱程度对枣树苗木水分状况的影响
水力结构功能维持是植物在干旱条件下存活的基础,水势是体现植物水分状况的直接指标,通常情况下凌晨水势是植物一天中的最高值,体现植物最佳水分状况,正午水势是水势日变化中的最低值,体现植物的最差水分状况,在本研究中水势呈现相同的趋势,即随着水分状况的变差,枣树苗木水势降低。前人的大部分研究均呈现这一规律[2,9-10]。
在干旱条件下,水势降低,造成了木质部导管内树液的拉力增加,在植物蒸腾拉力驱动的负压水分输导系统中,较大的负压易于导致木质部导管的气穴栓塞,破坏植物水力结构的完整性,限制水分长距离输导,对植物水力学的研究表明水分长距离运输障碍是干旱导致植物死亡的主要原因[10-11]。在本研究中,随着干旱胁迫程度的加重,枝条导水损失率显著增加,正常供水处理的枝条导水损失率为46.2%±7.2%,重度干旱处理达到77.6%±4.0%。导水损失率大小受两方面因素的影响:一是植物的栓塞脆弱性,即栓塞脆弱性越大,在较大负压下越容易发生气穴栓塞[11-12];另一方面是栓塞修复能力,植物可以通过根压和渗透调节确定导管内气穴栓塞的短期修复,也可以通过生长进行较长时间内的修复[13]。干旱通常会造成栓塞修复能力的下降,严重的干旱也会导致栓塞脆弱性的升高,二者的共同作用导致了重度干旱下枝条导水损失率接近80%,但栓塞脆弱性改变和栓塞修复能力变弱在导致导水损失率升高中的作用有待进一步研究。前人的研究通常把导水损失率88%或90%作为阔叶树种的致死程度[14],在本研究中虽然未达到这一标准,但对植物的水分输导速率会产生较大影响,进而会影响到叶片水分状况和光合作用的进行。
干旱对植物光合作用的限制主要是因为两方面:一是水分限制造成的气孔导度下降,限制了二氧化碳进入叶片,进而限制光合速率;另一方面是干旱造成的水分匮乏对光合系统的直接损伤[15]。在本研究中,中度和重度干旱显著降低了叶片的光合速率,从水分利用效率分析,轻度和中度干旱的水分利用效率有一定的提升,这主要是因为在轻度和中度干旱下,水分匮乏对气孔的限制作用大于对光合系统的影响,因此水分利用效率有一定提升。重度干旱水分利用效率较轻度和中度干旱有一定程度的下降,这可能与重度干旱造成了光合系统本身的损伤有关。
3.2 不同干旱程度对枣树生长和生物量分配的影响
干旱影响了植物的水分状况,进而限制了光合碳摄取能力,进一步会限制植物的生长,并影响生物量分配模式[16]。在本研究中,随着干旱程度的增加,枣树的株高成下降趋势,但对基径的影响不大,这表明干旱更多地影响到了枣树地上部分的伸长生长,对增粗生长的限制较小。重度干旱限制了枣树的不同部位生长,但轻度干旱对生长的影响较小。同时干旱也降低了枣树的叶面积,这在一定程度上能够降低蒸腾面积,但也会降低光合面积,对枣树水分状况有利但对碳摄取不利。
本研究也发现重度干旱显著降低了所有部位的生物量,但轻度干旱和中度干旱对生物量的降低程度较小,因此轻度干旱和中度干旱的总生物量与正常供水处理没有显著性差异,重度干旱显著降低了总生物量。本研究也发现重度干旱显著降低了细根的生物量比例,这可能是光合产物分配不足造成的,根系距离光合器官最远,因此会同时影响到光合碳摄取能力和光合产物的韧皮部运输能力[17-18],因此两方面共同的结果导致了细根的生物量和细根生物量占比均有较大幅度的下降,这将进一步导致水分吸收能力受限。随着干旱程度的增加,叶质量占比也有小幅度的降低,但下降程度较根小,这可能与植物的碳素分配有关,叶的发生部位距离光合器官较近,因此与根相比不容易受到碳分配不足的影响。
3.3 不同干旱程度对枣树NSC 含量的影响
NSC 是植物碳摄取和利用间的缓冲,可以用于生长、储存等功能,也可以用于渗透调节、木质部导管气穴栓塞修复的生理功能,对在干旱下的维持有重要作用[19]。在本研究中,轻度和中度干旱促进了不同部位的可溶性糖含量增加,但NSC 含量没有显著变化。这主要是由于淀粉向可溶性糖转化造成,在干旱条件下需要更多的可溶性糖提供渗透调节、干旱损伤修复,导管气穴栓塞修复等生理功能,因此干旱会推动淀粉向可溶性糖转化,同时在干旱条件下对树木生长的限制作用也会造成NSC 的积累,对植物的抗旱和旱后恢复有利[20]。重度干旱不同部位的可溶性糖含量和NSC 含量均有一定程度的下降。尤其在枝条韧皮部、粗根韧皮部和细根,NSC 含量下降达到显著性水平。这表明严重干旱导致枣树NSC 含量降低,枣树受到一定程度的“碳限制”影响,尤其在根部“碳限制”体现得更为明显。这将在一定程度上影响其抗旱能力和干旱后的恢复。同时本研究也发现在重度干旱处理枝条和粗根韧皮部NSC 含量较正常供水处理有显著性降低,而同一部位木质部没有显著性变化,这可能是韧皮部NSC 向木质部运输有关,前人的研究也表明韧皮部NSC 向木质部运输,进而会驱动木质部气穴栓塞的修复[21],这可能是树木枝条、根木质部导管大量栓塞的主动响应。
根系的水平和垂直分布可能会对干旱胁迫有一定的响应,并能影响树木对干旱的适应性,在本研究中,盆栽处理不能体现树木根系分布的影响,同时苗木和成年树木的水分代谢、碳代谢间可能会有一定的差异。自然状态下成年枣树对不同程度干旱在水力结构和碳代谢上的响应和适应将是下一步研究的重点内容。
4 结 论
本研究测定了枣树苗木在不同干旱程度下的水分状况、光合作用、生长、生物量分配、不同部位的非结构性碳含量。结果表明:重度干旱条件下枣树苗木水分状况变差,木质部导管气穴栓塞导致的水分输导能力受限,并进一步限制了光合碳摄取能力;严重干旱限制了枣树苗木的生长,并降低了细根的生物量占比,严重干旱降低了枣树不同部位的NSC 含量,NSC 含量降低在根部表现更为明显,细根生物量降低和NSC 含量降低将进一步限制在干旱条件下的水分吸收能力。轻度和中度干旱对枣树苗木的水力结构和碳代谢影响较小。以上结果表明枣树对轻度和中度干旱有较强的适应能力,重度干旱导致的水分输导受限和根NSC 含量降低会影响到枣树水碳平衡,并影响到枣树的生长和之后的存活。
猜你喜欢
作文周刊·小学一年级版(2023年36期)2023-09-14 14:32:41
电脑报(2020年12期)2020-06-30 19:56:42
水土保持研究(2020年2期)2020-03-19 09:03:26
河北果树(2020年1期)2020-02-09 12:31:44
乡村科技(2019年21期)2019-02-21 09:19:29
生态学报(2018年17期)2018-10-18 02:26:48
四川农业大学学报(2018年3期)2018-09-04 08:38:36
癌症进展(2016年9期)2016-08-22 11:33:10
小说月刊(2015年10期)2015-04-23 08:51:44
声屏世界(2014年6期)2014-02-28 15:18:07
