
红海榄幼苗根系呼吸代谢对水淹胁迫的响应
2022-12-05 12:53程皓王友绍马晓宇
热带海洋学报 2022年6期
程皓, 王友绍, 马晓宇
1. 热带海洋环境国家重点实验室(中国科学院南海海洋研究所), 广东 广州 510301;
2. 中国科学院大亚湾海洋生物综合试验站, 广东 深圳 518121;
3. 南方海洋科学与工程广东省实验室(广州), 广东 广州 511458
红树林主要分布于热带和亚热带的河口、海湾和 潟湖中, 蕴藏着丰富的生物资源。红树林在潮间带有较为明显的成带现象, 随着高程的增加, 红树林群落从白骨壤等先锋物种, 逐渐过渡到以红树科植物为主的秋茄、木榄和红海榄等群落, 再到相对陆生的半红树植物群落(林鹏, 2001; 王友绍, 2021)。被誉为“海岸卫士”的红树林具有重要的生态价值,如生物固碳、水质净化以及防风固堤等(王友绍,2019; Mai et al, 2022)。作为海陆交错带的一类特殊生态系统, 红树林易受到全球气候变化和人类活动的双重威胁, 全球范围内红树林退化和消失的报道屡见不鲜(傅海峰 等, 2014; Lovelock et al, 2015)。为了扭转这一趋势, 我国自20 世纪90 年代以来, 相继在华南沿海开展了多项红树林恢复与重建计划, 并取得了巨大成效 (范航清 等, 2017)。
水分条件是影响红树林分布与生长的重要限制因子之一, 因为水淹会直接阻碍土壤和大气之间的气体交换, 导致植物根系缺氧和呼吸代谢的紊乱(Cheng et al, 2015)。三羧酸(tricarboxylic acid, TCA)循环有氧呼吸(简称TCA 呼吸)和无氧呼吸是植物呼吸代谢的两种常见的呼吸代谢途径(Armstrong et al,2019; Pan et al, 2021)。已有的研究结果表明, 缺氧条件会抑制黄瓜、桃树和大麦等植物根系细胞的TCA 循环效率, 导致三磷酸腺苷(adenosine triphosphate, ATP)生物合成受阻, 从而影响植物正常的生长, 甚至有可能引起植株死亡(He et al, 2015;Luan et al, 2018; Gu et al, 2021)。另外, 水淹厌氧胁迫下, 植物有可能启动无氧呼吸(如乙醇发酵和乳酸发酵), Zhang 等(2017)和Yin 等(2019)就曾报道水淹缺氧会诱导樱桃树和矮牵牛花无氧呼吸的上调。无氧呼吸可暂时减缓水淹胁迫所造成的ATP 匮乏,但无氧发酵代谢产物在根系细胞内聚集也可能对根系细胞产生毒害作用(Loreti et al, 2016; Pan et al,2019)。了解红树植物呼吸代谢对水淹胁迫的响应特征对于红树林的保护与造林有重要意义, 但目前有关水淹胁迫对植物呼吸代谢影响的相关研究主要集中在经济作物上(Loreti et al, 2016; 刘逸泠 等,2017), 仅有少数几篇文献探究了红树植物呼吸代谢对水淹胁迫的响应, 如Pezeshki 等(1997)和 Chen等(2005)曾报道水淹胁迫会诱导亮叶白骨壤、大红树和秋茄乙醇发酵的增强; 陈思婷(2021)则报道了围填海过程中高岭土淤积会导致白骨壤指状根气孔被堵塞, 从而使白骨壤遭受低氧胁迫, 诱导指状呼吸根乙醇发酵的增强以及细胞色素C 氧化酶活性上升。
红海榄广泛分布于华南沿海地区, 是我国红树林修复主要常见造林树种之一。为此, 本研究拟通过人工潮汐系统研究红海榄幼苗根系呼吸代谢对水淹胁迫的响应, 以期为红树林的保护与恢复提供基础数据和理论依据。
1 材料与方法
1.1 实验材料和水淹处理
实验所用的红海榄胚轴采摘于广东省湛江市高桥镇红树林保护区, 胚轴经高锰酸钾溶液消毒、清水冲洗后种植在干净河砂中, 以1/2 浓度的霍格兰营养液浇灌。待幼苗长出第三对叶后, 选择长势一致、无病虫害幼苗进行后续实验。
利用人工配置10‰的氯化钠溶液和人工潮汐系统装置, 模拟不同潮汐水淹环境。本实验共设4 个处理组(每天水淹0、6、12 和18h), 每个处理组有3盆, 每盆4 株植物, 其中同一盆中的4 株植物各生长或生理指标的平均值视为一个重复。实验持续时间为两个月, 温室玻璃房的温度为28±5℃, 光照条件约为300μmol·m–2·s–1。实验结束后, 收获植物并测定以下指标。
1.2 测定方法
1.2.1 植物生长指标测定
水淹处理两个月后, 收获植物, 放置于烘箱烘干, 分别测定地下根系生物量以及地上茎叶的生物量, 并计算地下部分/地上部分的生物量比值。
1.2.2 ATP 含量测定
ATP 含量测定参考前人的方法(Mohanty et al,1993)。己糖激酶催化葡萄糖和ATP 合成6-磷酸葡萄糖, 6-磷酸葡萄糖脱氢酶进一步催化6-磷酸葡萄糖生成还原态的烟酰胺腺嘌呤二核苷酸, 该物质在340nm 有特征吸收峰, 由此测定ATP 含量。利用试剂盒(BC0300, 索莱宝, 北京, 中国)测定红海榄幼苗根系中ATP 含量。
1.2.3 TCA 呼吸速率测定
TCA 呼吸速率的测定采用丙二酸抑制法(Bouma et al, 2001), 取0.05g 植物根系(直径1.5mm,长度2~3cm), 均匀切成2mm 根段后, 转移至离心管内后加入1.5mL 磷酸盐缓冲液(20mmol·L–1PBS, pH 6.8), 测量液体内单位时间氧浓度的变化, 作为总呼吸速率。配置0. 5mol·L–1的丙二酸-磷酸缓冲液溶液用来抑制TCA 循环, 将切好的根段转移至丙二酸-磷酸缓冲液溶液中, 测量溶液中氧浓度的变化, 作为剩余呼吸速率。TCA 呼吸速率计算公式如下:
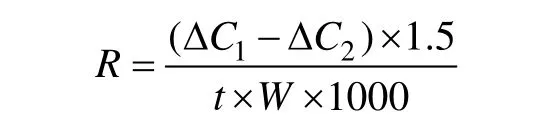
其中R是TCA 呼吸速率(μmol·min–1·g–1),1CΔ 和ΔC2是未加入和加入TCA 抑制剂时缓冲液中O2浓度的变化值,t是测试时间,W为根系的重量。
1.2.4 TCA 呼吸以及无氧呼吸关键酶活性测定
琥珀酸脱氢酶(succinate dehydrogenas, SDH)和苹果酸脱氢酶(malate dehydrogenase, MDH)是TCA循环的两个关键限速酶, 分别催化琥珀酸和苹果酸生成富马酸和草酰乙酸; 乙醇脱氢酶(alcohol dehydrogenase, ADH) 和乳酸脱氢酶(lactate dehydrogenase, LDH)所诱导的乙醇发酵和乳酸发酵则是植物无氧呼吸的主要方式, 为此, 本实验选择了上述4 个呼吸代谢关键酶, 利用分光光度计法,测定植物根系ADH、LDH、SDH 和MDH 的活性, 所用试剂盒均由索莱宝提供(BC1080、BC0680、BC0950、和BC1040, 索莱宝, 北京, 中国)(Li et al,2020; Luo et al, 2020, 2021)。
1.2.5 乙醇和乳酸含量测定
乙醇和乳酸的提取与测定参考前人的方法(Good et al, 1993), 称取0.5g 植株根系, 放入预冷的研钵中, 用预冷的1.6mL 0.6mol·L–1HClO4溶液提取,冰浴下研磨后静置30min, 在4 ℃ 10000×g下离心10min, 移液枪取出上清液, 用3mol·L–1KOH 调pH至4.5, 放置15min, 在4 ℃ 10000×g下离心10min,上清液即为提取待测液。乙醇测定反应混合液含有1mL 的 0.3mol·L–1K4P2O7(pH 9.0) 、 0.1mL 49mmol·L–1的 烟 酰 胺 腺 嘌 呤 二 核 苷 酸、0.02mL 75KU·L–1的乙醛脱氢酶、1.4mL 蒸馏水、0.1mL 酶液, 混合均匀, 测定340nm 波长的吸光值(A1), 用0.02mL 9000KU·L–1ADH 启动反应, 8min 后测定340nm 波长吸光值(A2), 以两次吸光值的差值(ΔA)计算乙醇含量。乳酸测定反应混合液含有0.67mL的 0.77mol·L–1谷 氨 酸 缓 冲 液(pH 8.9)、0.1mL 24mmol·L–1的烟酰胺腺嘌呤二核苷酸、0.05mL 780 KU·L–1的谷丙转氨酶、1.96mL 蒸馏水、0.2mL 酶液,混合均匀, 25℃下水浴保温10min, 测定340nm 波长的吸光值(A1), 用0.02mL 550KU·L–1LDH 启动反应,20min 后测定340nm 波长吸光值(A2), 以两次吸光值的差值(ΔA)带入公式计算乳酸含量。
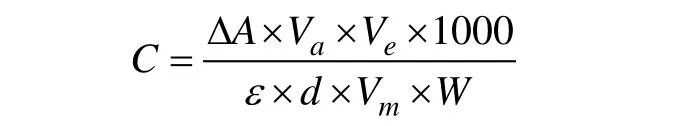
其中C为乙醇或乳酸的含量(μmol·g–1),AΔ 是吸光值的变化,Va为反应液的体积,Ve是总提取待测液的体积,ε是摩尔消光系数,d是比色杯量程,Vm是实验所用的提取液体积,W为根系的重量。
1.3 统计分析
使用 SPSS(statistical product and service solutions, SPSS)软件对实验数据进行统计分析, 实验结果表示为平均值±标准差, 通过单因素方差分析和最小显著性差异检验法比较样本的组间差异(P<0.05)。
2 结果与分析
2.1 红海榄对水淹胁迫的生长响应
红海榄对水淹胁迫的生长响应如表1 所示。与对照组相比, 适度的水淹(如每天水淹6h)不仅不会抑制红海榄幼苗的生长, 甚至还一定程度上促进了红海榄幼苗的生长。但是随着水淹时间的延长, 尤其是在每天水淹18h 处理组中, 红海榄生物量显著降低(P<0.05)。另外, 随着水淹时间的延长, 红海榄幼苗地下根系生物量的分配比例也显著下降。

表1 水淹胁迫下红海榄幼苗的生长以及生物量Tab. 1 Growth and biomass yield of R. stylosa subjected to waterlogging, different letters indicated significant variations between groups at P<0.05
2.2 水淹胁迫对红海榄根系ATP 含量的影响
红海榄根系ATP 含量对随水淹时间变化情况如图1 所示。随着水淹时间的延长, 红海榄根系ATP含量出现明显的下降趋势(P<0.05), 并在水淹18h 处理组中达到最低值。

图1 水淹胁迫对红海榄根系ATP 含量的影响不同字母代表组间存在显著性差异P<0.05Fig. 1 Effect of waterlogging on ATP content in the roots of R.stylosa, different letters indicated significant variations between groups at P<0.05
2.3 水淹胁迫对红海榄根系TCA 呼吸速率的影响
红海榄幼苗根系TCA 呼吸速率对水淹胁迫的响应如图2 所示。随水淹时间的延长, 红海榄幼苗根系TCA 呼吸速率呈明显的下降趋势, 且TCA 呼吸速率的最小值出现在每天水淹18h 处理组中。
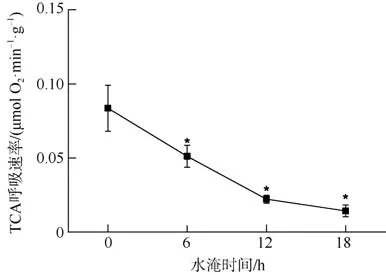
图2 水淹胁迫对红海榄根系TCA 呼吸速率的影响*代表与对照组有显著性差异(P<0.05)Fig. 2 Effect of waterlogging on TCA respiration rate in the roots of R. stylosa. * indicated significant differences with control at P<0.05
2.4 水淹胁迫对红海榄根系乙醇和乳酸含量的影响
水淹胁迫对红海榄根系乙醇和乳酸含量的影响如图3a、b 所示。每天水淹6h 胁迫下红海榄根系乙醇和乳酸含量与对照组无显著性差异, 但随着水淹时间的延长, 红海榄根系乙醇和乳酸含量均显著增加(P<0.05)。
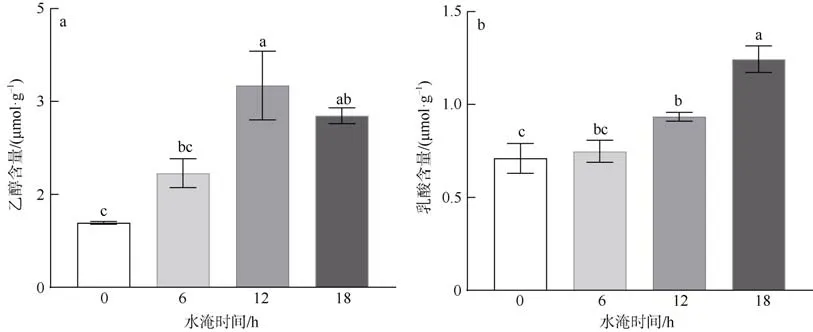
图3 水淹胁迫对红海榄根系无氧呼吸代谢产物的影响不同字母代表组间存在显著性差异(P<0.05)Fig. 3 Effect of waterlogging on anaerobic respiratory metabolites in the roots of R. stylosa, different letters indicated significant differences between groups at P<0.05
2.5 水淹胁迫对红海榄根系呼吸代谢关键酶活性的影响
红海榄根系TCA 呼吸关键限速酶随水淹时间变化情况如图4a、b 所示。结果表明, 水淹胁迫显著降低了MDH 酶活性, 即便低强度的水淹(每天水淹6h)也会显著降低红海榄幼苗根系中MDH 酶活性。低强度的水淹(每天水淹6h)对SDH 活性无显著影响, 但随着水淹时间的延长, 红海榄幼苗根系中SDH 活性显著降低。
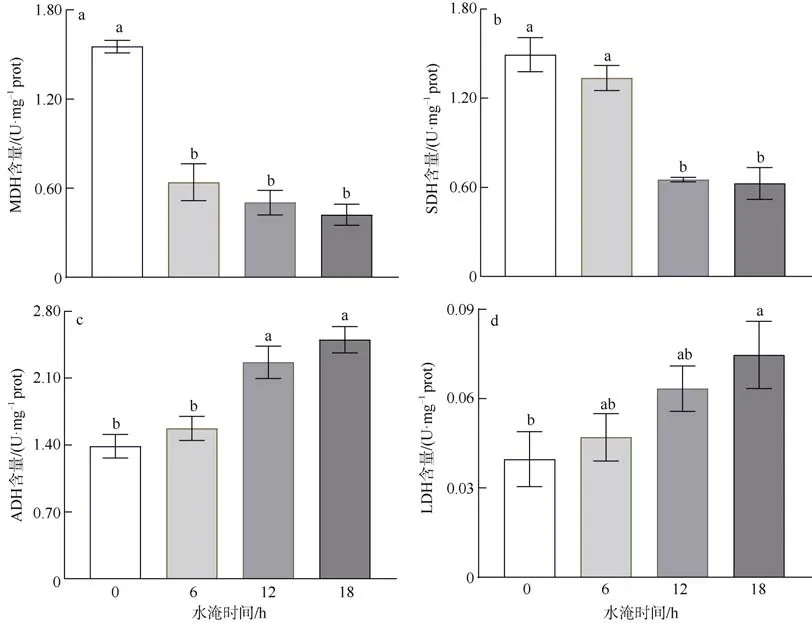
图4 水淹胁迫对红海榄根系TCA 循环和无氧呼吸关键酶的影响误差棒上不同字母代表组间存在显著性差异(P<0.05)Fig. 4 Effect of waterlogging on TCA and anaerobic respiration related enzymes in the roots of R. stylosa, different letters indicated significant variations between groups at P<0.05
红海榄根系无氧呼吸酶ADH 和LDH 活性对水淹胁迫的响应如图4c、d 所示。水淹胁迫, 尤其是长时间的水淹胁迫, 显著增加了红海榄幼苗根系中ADH 和LDH 活性。另外还发现, ADH 的活性要远远高于LDH 的活性。
3 讨论
尽管红树植物具备一定的耐淹能力, 但本研究的结果表明过度的水淹仍会直接抑制红海榄幼苗的生长。本研究中红海榄幼苗生物量最大值出现在每天水淹6h 处理组中, 其结果表明适宜的潮汐生境对红海榄幼苗的生长至关重要, 至于红海榄最优水淹时间则还需更进一步的研究。随着水淹时间的延长,红海榄幼苗的生长明显受到抑制(表1)。已有的研究结果(叶勇 等, 2007; 廖宝文 等, 2010; 代捷 等,2020)表明, 红树植物并不能忍受持续长时间的水淹,过度的水淹同样会直接抑制白骨壤、秋茄和木榄等红树植物正常的生长。
氧气在水中的扩散速率约为在空气中的万分之一, 因此潮汐淹水会阻碍土壤和大气之间的气体交换, 导致红树林底泥的缺氧(Bailey-Serres et al, 2012;Cheng et al, 2020a, b)。关于植物对水淹胁迫的适应机制, 目前有生活史策略与能量分配、结构适应以及代谢适应三种主流的观点。在长期的进化过程中,一些植物可以通过调节生命周期的长短来避免水淹伤害, 这类植物的特点是生命周期短, 在夏季洪水来临之前就完成了生殖生长, 以种子的形式埋藏在土壤中以躲避水淹(Warwick et al, 2003)。减少地下根系生物量分配比例也是植物对水淹胁迫的一种适应策略(刘逸泠 等, 2017; Fujita et al, 2021), 本研究也发现水淹胁迫会降低红海榄幼苗地下根系生物量/地上茎叶生物量的比值(表1)。地下根系生物量分配比例的降低将有助于减少根部的需氧和呼吸消耗,而地上部分分配比例的增加将扩大与大气的接触面积, 所有这些都能一定程度上帮助植物适应水淹缺氧生境(Cheng et al, 2015)。结构适应则主要强调通气组织的作用, 通气组织是指皮层细胞凋亡后所形成的空腔, 普遍存在于湿地植物根系中, 其形成有助于氧气向根系的传输, 缓解水淹厌氧伤害(Armstrong et al, 2019; Cheng et al, 2020b; Pedersen et al, 2021)。
本研究则首次研究了红海榄幼苗根系呼吸代谢对水淹胁迫的响应。我们发现红海榄根系TCA 呼吸速率以及ATP 产能效率对水淹缺氧十分敏感, 即便是每天水淹6h 也会显著降低红海榄根系TCA 呼吸速率和ATP 产能效率(图1、2)。有趣的是, 每天水淹6h 处理对红海榄幼苗的生长基本无负面影响, 其原因可能是因为, 非水淹周期内红海榄幼苗的生理代谢可能会部分恢复, 以维持植物正常的生长(Shingaki-Wells et al, 2014; Borella et al, 2019)。但随着水淹时间的延长, 红海榄TCA 呼吸和ATP 生成进一步受阻, 这将会抑制根系正常的生理活动以及对养分和水分吸收, 导致植物的生长和生物量的显著降低(Wang et al, 2019; Cheng et al, 2020a; Fujita et al,2021)。另外, 我们还发现红海榄幼苗根系SDH 和MDH 活性在水淹胁迫下呈明显的下降趋势(图4)。TCA 循环是碳水化合物、脂肪和蛋白质分解代谢的共同通路, 是ATP 合成的主要途径, 其中SDH 和MDH 是TCA 循环过程中两个关键限速酶(Miro and Ismail, 2013; Jiang et al, 2020)。SDH 是TCA 循环过程中唯一的一个位于线粒体内膜上的酶, 是氧化磷酸化和电子传递的枢纽之一, 已有的研究结果表明,厌氧环境不利于SDH 催化琥珀酸生成富马酸, 另外,糖酵解代谢产物丙酮酸的积累还会进一步抑制SDH与琥珀酸的结合, 导致TCA 循环效率的下降(Li et al, 2020)。MDH 则催化了TCA 循环过程中的最后一次脱氢反应, 伴随着水淹时间的延长以及MDH 活性的下降, 这将会阻碍草酰乙酸的再生, 迫使TCA循环中断, 抑制线粒体ATP 合成(Bailey-Serres et al,2012; Pan et al, 2021)。
相反, 红海榄幼苗根系细胞无氧呼吸关键酶(ADH 和LDH)的活性则在水淹胁迫下呈上升的趋势(图4)。张艳婷 等(2016)也曾报道水淹胁迫会显著增加“中山杉118”根系ADH 和LDH 酶的活性。无氧呼吸的增强可能是植物对水淹厌氧胁迫的一种适应策略, 在TCA 循环受阻的情况下维持糖酵解的继续进行, 为植物提供暂时和短期的ATP 能量供给(Pan et al, 2021)。已有的研究结果表明水稻根系在水淹胁迫下的替代呼吸途径有丙酮酸-乙醇、丙酮酸-乳酸及丙氨酸发酵途径, 其中通过糖酵解产生的丙酮酸中约有92%被用于丙酮酸-乙醇途径, 仅7%与1%的丙酮酸被分别用于丙酮酸-乳酸与丙氨酸发酵途径(Miro et al, 2013)。本实验中, 红海榄幼苗根系内ADH 活性要显著高于LDH 活性(图4), 表明红海榄受到水淹胁迫时同样主要以丙酮酸—乙醇途径作为主要的无氧呼吸替代途径。值得注意的是, 无氧呼吸的ATP 产能效率远不如TCA 呼吸(Loreti et al,2016; Jiang et al, 2017), 不能完全补偿TCA 呼吸受阻所导致的ATP 产能效率的下降。另外, 无氧呼吸的增加会导致无氧发酵产物在植物体的积累。从图3 可以发现, 过度的水淹会显著增加红海榄幼苗根系中乙醇和乳酸的含量。乙醇和乳酸均有一定的毒性, 能损伤线粒体结构、破坏蛋白结构并加剧膜脂伤害等(Tamang et al, 2014; Xu et al, 2016; Wang et al,2019)。但目前尚未开展乙醇和乳酸对红树植物毒害作用的相关研究, 另外, 随着现代分子生物技术的发展, 一些红树植物已完成了全基因组测序, 红树植物对水淹胁迫的代谢响应及其分子调控机制也值得进一步研究(王友绍, 2019; He et al, 2019)。
4 结论
本实验通过人工模拟潮水系统, 首次探究了水淹胁迫对红海榄幼苗根系呼吸代谢的影响。与对照相比, 每天淹水6h 对红海榄幼苗的生长影响不大,但随着水淹时间的延长, 红海榄幼苗的生物量显著降低。我们还发现水淹胁迫会显著抑制红海榄幼苗根系琥珀酸脱氢酶和苹果酸脱氢酶活性, 导致三羧酸循环有氧呼吸速率降低。无氧呼吸关键酶(乙醇脱氢酶和乳酸脱氢酶)活性以及无氧发酵代谢产物(乙醇和乳酸)含量在水淹胁迫下则呈上升趋势, 但是无氧呼吸的增强尚不足以补偿三羧酸循环受阻所导致的三磷酸腺苷产能效率下降。研究结果还进一步表明, 水淹缺氧条件下丙酮酸—乙醇途径是红海榄主要的无氧呼吸替代途径。本研究将有助于更好地了解红树植物对水淹胁迫的响应和适应特征, 为红树林的保护与修复提供基础数据和理论依据。
猜你喜欢
汽车实用技术(2022年16期)2022-09-03
儿童故事画报·自然探秘(2022年3期)2022-04-27
幼儿园(2021年4期)2021-07-28
儿童时代·幸福宝宝(2020年3期)2020-04-07
青少年科技博览(中学版)(2019年12期)2019-04-10
小哥白尼(军事科学)(2018年3期)2018-06-15
歌海(2018年4期)2018-05-14
小学科学(学生版)(2017年11期)2017-12-20
小学科学(2017年11期)2017-11-23
天然气勘探与开发(2012年2期)2012-03-25
