
不同耕作模式对稻田土壤真菌丰度及群落结构的影响
2022-12-03 06:26:48徐慧芳万子维盛荣魏文学张文钊黄国勤
华中农业大学学报(自然科学版) 2022年6期
徐慧芳,万子维,盛荣,魏文学,张文钊,黄国勤
1.江西农业大学生态科学研究中心,南昌 330045;2.中国科学院亚热带农业生态研究所桃源农业生态试验站/亚热带农业生态过程重点实验室,长沙 410125
土壤微生物群落多样性作为评价土壤质量的关键指标,已越来越引起人们的广泛关注[1]。微生物在土壤生物化学循环中扮演着重要角色,研究表明,不同的农业管理措施将导致土壤微生物群落结构及其多样性的变化[2],进而影响农业的可持续发展。因此,研究稻田长期不同耕作模式下土壤微生物群落结构及多样性的变化,可为制定合理的耕作措施及改善土壤质量提供重要依据。
真菌在土壤中的数量仅次于细菌和放线菌,其在有机质分解[2]、土壤养分固定[3]、促进植物养分吸收[4]及增强土壤抗盐碱能力[5]等过程中均发挥着重要作用,是维持生态系统健康的关键指示物。人为干扰、轮作方式及土地管理措施均会影响土壤真菌群落组成[1,3,6]。目前国内外针对轮作方式对真菌群落结构的影响已开展了一些研究。例如,Behnke等[7]研究发现,与玉米单作相比,玉米-大豆轮作(旱旱轮作)处理显著增加了真菌的数量。阳祥等[8]研究表明水旱轮作模式增加了土壤真菌多样性指数。苏婷等[9]研究发现,相较双季玉米和单季水稻种植方式,玉米-水稻的水旱轮作显著增加了土壤真菌的数量。
双季稻在我国水稻种植历史中占据重要的地位,对于保障我国粮食安全意义重大。然而,近年来由于双季稻种植的经济效益逐渐下滑,加之农村劳动力大量转移,这些因素促使农民将部分长期淹水的双季稻田转化为单季稻或者水旱轮作等种植体系。研究表明,水旱轮作引起了土壤养分元素形态和有效性的变化[10],但养分元素的变化是否会导致土壤真菌丰度及群落结构变化的研究较少。
因此,本研究通过实时荧光定量PCR 和高通量测序技术研究我国典型稻田耕作模式(双季稻,中稻,中稻-油菜,中稻-小白菜-油菜)对土壤真菌的丰度及群落组成的影响,旨在为稻田生态系统可持续发展等相关措施的制定提供重要的科学依据。
1 材料与方法
1.1 试验样地及土壤样品采集
试验样地位于湖南省桃源县中国科学院桃源农业生态试验站水田长期耕作模式试验田(2012 年至今),东经111°27΄,北纬28°55΄。该地区属于我国典型的中亚热带向北亚热带过渡的季风湿润气候区,年均温16.5 ℃,年降雨1 447.9 mm,年平均日照1 531.4 h。供试的土壤为第四纪红色黏土发育的红壤,试验前为早稻-晚稻种植模式。试验前耕层土壤(0~15 cm)的有机质含量26.08 g/kg,全氮1.28 g/kg,全磷0.54 g/kg,全钾12.79 g/kg,pH 4.84。
试验田设置4 个处理,分别为:中稻、双季稻(早稻-晚稻)、中稻-油菜、中稻-小白菜-油菜,记为MR、DR、MROR、MPOR,每个处理均设置3个野外重复。不同处理之间的施肥种类和施肥用量详见表1。

表1 不同耕作模式下稻田施肥状况Table 1 Amount of fertilizer applied of four farming patterns kg/m2
MR 模式下,中稻的氮肥按照基肥∶分蘖肥∶穗肥=4∶5∶1 施用,磷钾肥作基肥一次施入。有机质含量为27.34 g/kg,全氮1.58 g/kg,全磷0.64 g/kg,全钾12.11 g/kg,pH 5.13,水分含量28.66%,铵态氮含量2.53 g/kg,硝态氮含量0.51 g/kg,可溶性有机碳
84 .57 mg/kg。
DR 模式下,早稻的氮肥按照基肥∶分蘖肥=4∶6 施用,磷钾肥作基肥一次施入,而晚稻采用氮肥按照基肥∶分蘖肥∶穗肥=4∶5∶1 施用,磷钾肥作基肥一次施入。有机质含量为34.38 g/kg,全氮1.60 g/kg,全磷0.6 g/kg,全钾12.44 g/kg,pH 5.17,水分含量31.33%,铵态氮含量2.78 g/kg,硝态氮含量0.61 g/kg,可溶性有机碳83.22 mg/kg。
MROR 模式下,复合肥和有机肥均作为基肥一次施用。有机质含量为26.23 g/kg,全氮1.47 g/kg,全磷0.57 g/kg,全钾13.58 g/kg,pH 5.22,水分含量26.33%,铵态氮含量1.92 g/kg,硝态氮含量1.19 g/kg,可溶性有机碳85.46 mg/kg。
MRPOR 模式下,有机肥均作为基肥一次施用,小白菜移栽40 d 后收获,移栽油菜。有机质含量为26.04 g/kg,全 氮1.36 g/kg,全 磷0.60 g/kg,全 钾13.00 g/kg,pH 5.38,水分含量25.33%,铵态氮含量1.87 g/kg,硝态氮含量1.04 g/kg,可溶性有机碳78.76 mg/kg。
2017 年晚稻收获后于11 月3 日采用随机多点对不同耕作模式的0~15 cm 的耕作层土壤样品进行取样,去除明显的杂质和根系后,充分混匀后分为两部分,一部分约200 g 土壤样品,用锡箔纸包装后,装入灭菌且写有标签的布袋并置于液氮罐中,于−80 ℃冰箱保存,用于真菌丰度及群落结构分析;另一部分新鲜土壤样品放入冰盒后于4 ℃冰箱保存,用于土壤理化性质的测定。
1.2 土壤理化性质的测定
土壤理化性质采用常规分析方法[11]。土壤全氮用H2SO4消化后采用流动注射分析仪进行测定(FI⁃Astar5000,FOSS,Sweden);全磷经过高氯酸-硫酸法熔融后采用钼锑抗比色法测定;全钾用氢氧化钠灼烧法后采用火焰光度法测定;土壤pH 采用水土比1∶1,用pH 计 测 定(FE-20,METTLER TOLE⁃DO,China);土壤有机质通过重铬酸钾加热后,采用硫酸亚铁铵进行滴定测定;土壤硝态氮和铵态氮用2 mol/L KCl浸提后,采用连续流动分析仪测定(Flastar 5000 Analyzer)测定;土壤可溶性有机碳(DOC)采用0.5 mol/L 的K2SO4浸提后测定其含量;采用烘干法测定土壤质量含水量。
1.3 土壤总DNA提取
土壤微生物总DNA 的提取主要采用Fast DNA SPIN 提取土壤DNA 试剂盒(MP Biomedicals,San⁃ta Ana,CA)。提取的土壤DNA 用50 μL 无菌水进行稀释,然后用1%的琼脂糖凝胶检测其质量,最后采用Nanodrop ND-1000UV-Vis 分光光度计测定DNA 的浓度及质量系数(Nanodrop Technologies,Wilmington,DE,USA)。凝胶电泳检测图显示所提取的土壤总DNA 条带未发生弥散,完整度较高,可直接用于后续的PCR扩增。
1.4 荧光定量PCR扩增及高通量测序
采 用 Nu-ssu-0817F (5′- TTAGCATG⁃GAATAATRRAATAGGA-3΄) 和 Nu-ssu-1196R(5΄-TTAGCATGGAATAATRRAATAGGA-3΄)对18S rRNA 进行扩增。扩增体系为10 μL,包括上游引物(10 μmol/L)0.2 μL,下游引物(10 μmol/L)0.2 μL,5 μL 的SYBR GreenⅡ(TaKaRa),0.2 μL 的Rox(TaKaRa),DNA 为1 μL(5 ng),补无酶水至10 μL。荧光定量PCR 扩增程序为:95 °C 30 s,40 个循环的95°C 5 s,60°C 30 s。
由于18S rRNA 在大多数生物中趋于保守,生物之间的基因组序列变化不大,而其内转录间隔区(ITS)作为非编码区相对变化较大,并且在物种注释分析时可以提供更加详细的物种信息,因此,我们采用ITS 区域鉴定真菌的物种组成[12]。运用ITS1F(5΄ -CTTGGTCATTTAGAGGAAGTAA-3΄) 和ITS2R(5΄-GCTGCGTTCTTCATCGATGC-3΄)[13]对ITS 序列进行PCR 扩增。扩增体系为Mix(TaKa⁃Ra)12.5 μL,上游引物(10 μmol/L)7.5 μL,下游引物(10 μmol/L)7.5 μL,DNA 为2 μL(40 ng),补无酶水至25 μL。PCR扩增程序为:95 ℃10 min,94 ℃30 s,55 ℃30 s,72 ℃1 min,40 个循环;74 ℃9 min。PCR产物经琼脂糖凝胶电泳,使用胶回收试剂盒纯化后送到上海美吉生物医药科技有限公司进行高通量测序,基于Illumina Miseq PE300 测序平台进行双端测序。
1.5 高通量测序数据OTU聚类及群落组成分析
高通量测序得到的双端序列数据,根据序列首尾两端barcode 和引物序列区分样品得到有效序列。过滤去杂的数据采用97%进行OTU(operational taxonomic units)聚类。为了得到每个OTU 对应的物种分类信息,采用RDP classifier 贝叶斯算法对各个基因的OTU 代表序列进行分类学分析,设置分类水平默认置信度阈值为0.8,并在phylum(门)水平上统计各个样本的群落组成。真菌ITS 数据库为UNITE 8.0(http://unite.ut.ee/index.php)[14]。真菌ITS 的高通量测序数据已经上传至NCBI 数据库(No.PRJNA836033)。
真菌在不同耕作模式之间的群落组成差异分析采用基于欧式距离的PCA 主成分分析(principal component analysis)。Venn 图用于分析不同耕作模式下真菌群落的结构相似性。采用CANOCO 5.0的db-RDA(distance-based redundancy analysis)分析环境因子与真菌群落结构的关系,其中采取前置选择法(forward selection)确定每个环境因子对真菌群落结构的相对贡献率。
1.6 数据处理
使用SPSS 25.0 软件对数据进行统计分析。采用单因素方差分析(one-way ANOVA)比较不同耕作模式下真菌18S rRNA 丰度的差异性;数据结果以“平均值±标准差”表示,n=3;采用LSD 多重比较对数据进行差异显著性检验(α=0.05);采用Origin 2021和R软件进行绘图。
2 结果与分析
2.1 不同耕作模式下稻田土壤的真菌丰度
从图1可知,不同耕作模式之间真菌丰度存在显著差异。真菌丰度在MRPOR 处理(4.15 × 109copies/g)中显著高于MROR(2.05 × 109copies/g)与DR(3.6×109copies/g)(P<0.05)。MR(2.86×109copies/g)的真菌丰度显著高于MROR(P<0.05),但DR 与MRPOR、MR 与MRPOR 之间的真菌数量差异不显著(P>0.05)。
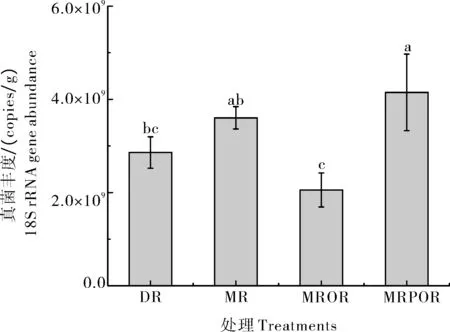
图1 不同耕作模式土壤真菌丰度的变化Fig.1 The abundance of soil fungi under different farming patterns
2.2 不同耕作模式下稻田土壤的真菌群落组成
4 种典型的稻田耕作模式对真菌群落结构的影响见PCA 分析图(图2)。由图2 可见,PCA 分析共提取了2 个主成分PCA1 和PCA2,二者方差累计贡献率为46.84%。代表不同处理的点距离越远,表明真菌群落结构差异性越大。DR 与MR 环境样本点的真菌群落结构单独聚在一起,而MROR 与MRPOR环境样点的真菌群落结构没有明显分离,说明双季稻(DR)转为单季稻和水旱轮作耕作模式后显著改变了真菌的β-多样性。
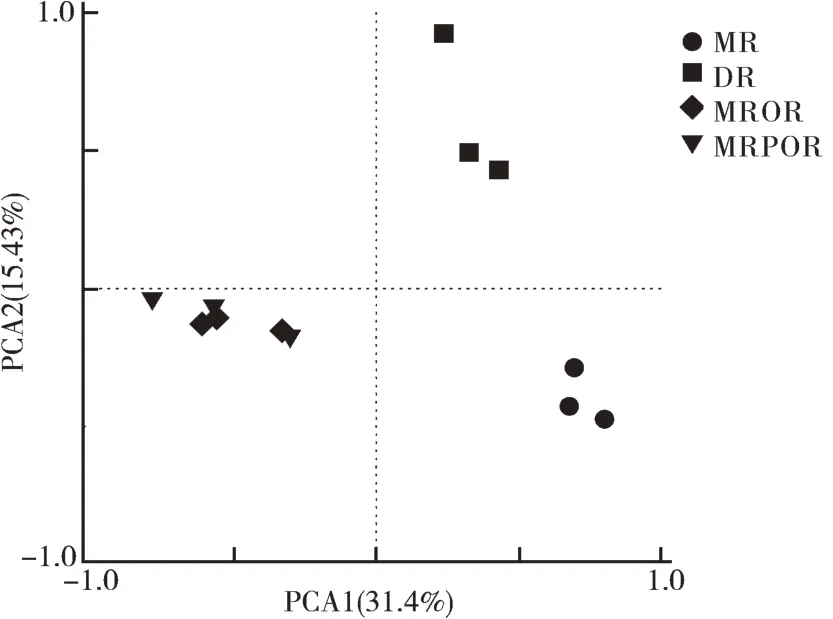
图2 不同耕作模式土壤真菌的主成分分析PCAFig.2 PCA analysis for soil fungi under different farming patterns
2.3 不同耕作模式下稻田土壤的真菌物种组成
如图3 所示,不同耕作模式对土壤真菌OTU 有明显影响。4 个处理共产生3 990 个OTU,其中共有的OTU 有358 个,占总OTU 数量的8.97%。DR、MR、MROR 及MRPOR 分别具有1 354、1 625、1 515及1 782 个OTU。MRPOR 独有的OTU 数量最多(826),而MROR 独有的OTU 数量最少(590)。MR与MRPOR 共有OTU 最多(659),DR 与MROR 共有的OTU 最少(500),结果表明,不同耕作模式下稻田土壤真菌组成存在差异,MR、MROR 及MRPOR均增加了土壤真菌类群。

图3 不同耕作模式土壤真菌群落Venn图Fig.3 Venn of soil fungi under different farming patterns
由图4 可见,4 个处理检测出的真菌门主要包括子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、壶菌门(Chytridiomycota)、球囊菌门(Glomeromyco⁃ta)、接合菌门(Zygomycota)及分类地位不确定真菌(Unclassified fungi)。其中,子囊菌门(24.12% ~31.77%)与担子菌门(7.07%~24.71%)为优势菌门,均为常见门类。与DR 相比,MR、MROR、MRPOR 子囊菌门与接合菌门的相对丰度均增加,壶菌门的相对丰度均降低;担子菌门的相对丰度在MRPOR 处 理 中 增 加,而 在MR 及MROR 处 理 中减少。
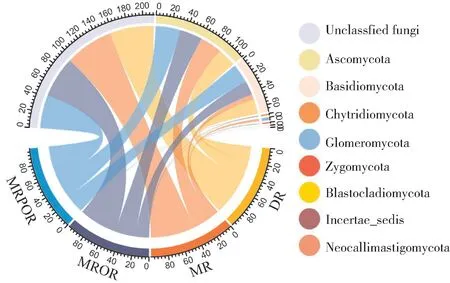
图4 不同耕作模式土壤真菌门水平群落组成Fig.4 Composition of soil fungi phylum under different farming patterns
2.4 不同耕作模式下稻田土壤的真菌群落多样性
不同耕作模式下稻田土壤真菌群落的Alpha 多样性指数如表2 所示。其中Chao1 和ACE 表示菌群丰度,Shannon 和Simpson 表示菌群多样性,PD whole tree 表示系统发育多样性,Coverage 表示观察到的物种覆盖度。由表2可知,各处理覆盖度均达到99%以上,说明各样本测序深度合理。MROR 与MRPOR 的Shannon 指 数 显 著 高 于MR 与DR,而Simpson 指数正好相反。MRPOR 的真菌丰富度及系统发育多样性均显著高于其他3个处理。

表2 不同耕作模式下土壤真菌群落的Alpha多样性指数Table 2 The fungi Alpha diversities of different genes among four farming patterns
2.5 不同耕作模式下稻田土壤的真菌丰度及群落结构与环境因子的相关性
对真菌丰度与环境因子进行Pearson 相关性分析,结果表明真菌数量与TK、pH、NO3−-N、水分含量的相关系数分别为0.505、0.707、0.744、0.123,其中真菌丰度与pH 及NO3−-N 呈现显著正相关(P<0.05);真菌数量与SOM、TN、TP、NH4+-N及作物产量的相关系数分别为−0.341、−0.644、−0.388、−0.633 及−0.123,其中真菌丰度与TN 及NH4+-N 呈现极显著的负相关(P<0.01)。
由图5可见,环境因子对土壤真菌群落结构的解释量前2 个排序轴分别为43.59%和20.98%。从箭头长度可见,TN、NH4+-N、土壤含水量及作物产量与真菌群落结构分布相关性较好且均达到显著性水平(P<0.05)。其中土壤含水量对于不同耕作模式下真菌群落结构分布差异的解释率最高,为39.5%,而TK解释率最低,为10.4%。
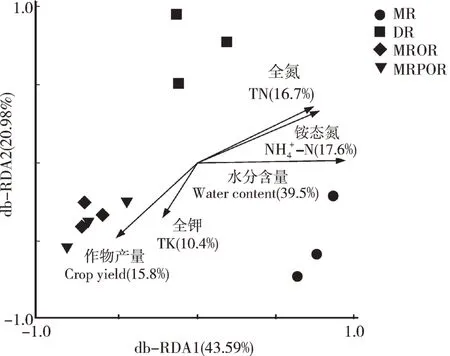
图5 不同耕作模式土壤真菌群落结构的db-RDA图Fig.5 db-RDA of soil fungi under different farming patterns
3 讨 论
土壤微生物的群落结构是土壤生态环境的重要生物指标。本研究结果表明,双季稻转变为中稻-小白菜-油菜后土壤真菌的数量显著增加。通常pH[15]、温度[16]、养分有效性[4]等被认为是影响真菌数量的关键环境因子。在本研究中,虽然pH 与真菌丰度存在极显著的正相关关系,但4种稻田轮作模式之间pH 值无明显差异(5.13~5.38)。因此,pH 可能不是导致中稻-小白菜-油菜模式下真菌数量增加的因素。而土壤硝态氮也与真菌丰度呈显著正相关关系。造成这种现象的原因可能有以下两方面:其一,相比双季稻,在长期施肥的情况下中稻-小白菜-油菜的土壤硝态氮含量比双季稻增加了近1倍,促进了微生物的代谢过程,这与周玉杰等[17]认为土壤养分是影响土壤真菌群落分布的重要因素的结论相一致;其二,相较双季稻,中稻-小白菜-油菜的水旱轮作处理下作物会分泌大量的单糖、多糖、氨基酸和有机酸等碳水化合物,为根际土壤真菌提供了碳源,促进了土壤真菌的繁殖[18],特别是水旱轮作条件下,土壤处于好氧和厌氧活动反复交替状态,厌氧条件下有利于形成腐殖质,好氧条件下有利于产生CO2,进一步有利于形成团粒结构的土壤,而团粒结构有助于真菌类群的栖息繁殖[19]。但在本研究中双季稻转为中稻-油菜后,其土壤真菌数量却显著下降。原因是虽然中稻-小白菜-油菜与中稻-油菜均为水旱轮作处理,但是中稻-油菜比中稻-小白菜-油菜减少了1 茬旱作,导致中稻-油菜植物残体数量低于中稻-小白菜-油菜。同时有研究表明,绝大多的真菌为腐生性微生物,具有分解多种植物纤维素、木质素等难降解有机质的能力[20]。而植物残体作为腐生微生物主要营养物质,其数量的减少会直接导致真菌丰度的降低[21]。此外,本研究的物种注释结果也显示,子囊菌门(24.12%~31.77%)与 担 子 菌 门(7.07%~24.71%)为不同耕作模式下土壤中的主要真菌门类,二者均为腐生营养为主[6,22],也印证了我们的猜测。
本研究中,不同轮作模式显著改变了真菌的群落结构,这与张慧等[22]的研究结果相似。此外,双季稻转变为中稻及水旱轮作模式后真菌的Shannon 多样性指数显著增加。双季稻转变为水旱轮作模式后,通常会使土壤变得疏松,从而增加了土壤的透气性能,有利于真菌的多样性增加[23]。这表明,水旱轮作模式在提升土壤真菌群落多样性方面具有一定的优越性,这与阳祥等[8]对不同轮作模式下土壤真菌群落结构及功能特征的分析结果相一致,即不同耕作模式会引起土壤pH、速效养分等土壤理化指标的改变,而这些土壤理化因子的改变又与真菌的群落组成密切相关。本研究表明土壤含水量是影响土壤真菌群落结构分布差异关键的环境因素,这与其他学者[24-26]探讨水分对真菌群落结构影响的结果一致,均认为水分含量是影响真菌的群落组成的关键因子。此外,子囊菌门(Basidiomycota)是不同耕作模式下的优势种群,其作为典型的陆生型真菌们更加适应土壤水分低及通气条件好的环境。土壤TN、NH4+-N 及作物产量也是影响真菌群落结构分布差异的关键影响因子。本研究表明调控不同耕作模式真菌群落结构的因素复杂多样,并不是由单一的环境因素决定真菌在不同耕作模式下的分布差异,而是各因素共同调控的结果。
不同耕作模式下稻田土壤理化性质的差异导致了土壤真菌丰度及群落结构的差异。其中双季稻转变为中稻-小白菜-油菜种植模式后增加了土壤的硝态氮含量,进而提高了真菌的丰度;而由于双季稻转变为单季稻或者水旱轮作种植模式后改变了土壤的水分含量,间接影响了真菌对土壤养分的利用,导致真菌群落结构发生改变。双季稻转换为单季稻或者水旱轮作过程中改善了土壤的养分状况,提高了真菌的丰度及多样性,这些结果有助于更好地选择科学的轮作模式以改善土壤微生物群落结构并提高土壤生产力。
猜你喜欢
华南农业大学学报(2024年6期)2024-01-01 00:00:00
今日农业(2021年21期)2022-01-12 06:31:28
农村实用技术(2021年9期)2021-10-30 02:11:26
现代农业科技(2017年5期)2017-04-19 20:11:53
湖南农业(2017年1期)2017-03-20 14:04:47
现代农业科技(2016年21期)2017-03-06 11:12:47
中国稻米(2016年1期)2016-06-29 07:59:53
中国稻米(2016年1期)2016-06-29 07:59:38
湖南农业(2016年3期)2016-06-05 09:37:36
湖南农业(2015年5期)2015-02-26 07:32:28
