
circUTRN24通过肝星状细胞自噬介导胆道闭锁肝纤维化
2022-12-02 15:13:32刘冬高家辉张曦耘梅倩倩吴宙光
岭南现代临床外科 2022年5期
刘冬,高家辉,张曦耘,梅倩倩,吴宙光
胆道闭锁(biliary atresia,BA)是一种发生于肝内外胆管的以炎性细胞浸润和纤维化闭塞为特征的新生儿肝脏疾病[1]。若不能及时地发现并加以干预,大多数患儿将在生后数周内迅速地发展为肝纤维化,甚至导致肝硬化肝衰竭的严重后果,这使其成为了小儿肝移植最常见的原因之一[2]。尽管肝门空肠吻合术(Kasai术)[3]使多数患儿实现了胆汁的再次引流,但仍未能改变肝纤维化肝硬化的结局,最终仍需接受肝移植治疗[4]。不良的预后和巨额的费用,使家庭承受了沉痛的打击并给社会带来了极大的负担。然而胆道闭锁肝纤维化的机制目前尚不清楚,这给临床诊断和救治带来了巨大的挑战。
环状RNA(circular RNA,circRNA)是一种具有特殊闭合环状结构的非编码RNA,是近些年的研究热点之一。它的5′和3′末端以非共价连接形成闭合环状结构区别于以往的线性RNA。circRNA较早被发现在RNA病毒中[5],但由于受早期检测技术所限而被人们所忽视。随着基因测序技术的发展,circRNA被证实普遍的存在于真核生物细胞中,其多样的生物学功能逐渐为人们所认识;越来越多的circRNA被发现参与了机体的代谢、自噬、凋亡等过程,并在各种人类疾病中扮演着重要的角色[6,7]。本课题组在之前的研究中通过高通量测序发现[8],胆道闭锁患儿肝组织中circRNA表达与对照组存在明显差异,其中circUTRN24的表达明显高于对照组并可能与自噬途径相关。目前研究发现细胞自噬可以参与肝星状细胞(hepatic stellate cell,HSC)的活化,而HSC的活化被公认为是造成肝纤维化的核心环节[9]。然而circUTRN24与自噬的关联以及是否参与胆道闭锁肝纤维化的过程,目前尚无报道。
本文旨在探究胆道闭锁中circUTRN24与自噬途径的关联,以及是否通过自噬参与胆道闭锁的肝纤维化过程,为胆道闭锁的抗肝纤维化治疗开拓新的方向。
1 资料与方法
1.1 一般资料
1.1.1 肝组织样本 本研究纳入2021年7月至2021年12月期间的10例胆道闭锁患儿和10例肝功能正常的胆总管囊肿(choledochal cyst,CC)患儿作为对照。所有患儿均由深圳市儿童医院同一外科团队完成腹腔镜胆道手术。BA组患儿平均日龄为60.3 d(34~81 d),CC组患儿平均日龄为251.3 d(15~511 d),两组中男女比例均为2∶3。肝组织标本离体后立即置于-80℃冰箱内保存,并在3个月内完成RNA及蛋白质的提取。本研究由深圳市儿童医院伦理委员会审核批准,标本采集均征得患儿家属同意并已签署知情同意书。
1.1.2 细胞系及培养 肝星状细胞(HSC)的激活被证明参与了肝纤维化的进展,我们的研究使用了LX⁃2细胞,即具有肌成纤维细胞样表型的永生化人HSC,细胞系从ATCC细胞库购得。所有细胞均培养于含10%胎牛血清的DMEM培养液中,细胞约2 d可生长至培养皿的80%。
1.2 实验方法
1.2.1 实时荧光定量PCR(RT⁃qPCR)检测肝组织中circRNA的表达水平 取1 cm×1 cm×1 cm大小的肝组织用TRIzol试剂提取总RNA,按照反转录试剂盒说明,将1μg的总RNA加入最终体积为10μL的混合试剂中进行反转录。在ABI 7500实时PCR系统上使用SYBR Green Mix试剂盒进行实时定量PCR。通过分析熔解曲线以验证PCR的可靠性。甘油醛⁃3⁃磷酸脱氢酶(glyceraldehyde⁃3⁃phosphate dehydrogenase,GAPDH)被用作内参。按照MIQE指南[10]的要求进行RT⁃qPCR实验,通过2⁃ΔΔCt法计算circRNA的相对表达量,每个样本重复3次独立实验结果取均值。引物序列如下:circ⁃UTRN24上游序列为(5′⁃3′):ATCTTCTTGGCCTT⁃CACCCC;下游序列为(5′⁃3′):CAGTTCAACAGC⁃TCTGCACTTA。GAPDH上游序列为(5′⁃3′):GGT⁃GAAGGTCGGAGTCAACG,下游序列为(5′⁃3′):CCATGTAGTTGAGGTCAATGAAG。
1.2.2 蛋白免疫印迹(Western blot,WB)法检测组织和细胞中的蛋白表达水平 提取各组肝组织和细胞中的总蛋白并通过BCA法测定蛋白浓度。将含50μg总蛋白的裂解产物进行SDS⁃PAGE电泳,100 V电泳至Marker前缘接近胶底,适时停止电泳。湿式电转仪100V稳压转印2.5 h至PVDF膜上,含5%脱脂奶粉的TBST(Tris Buffered Saline with Tween)室温封闭2 h。分别加入对应的Beclin⁃1、LC3B、α⁃SMA或COL⁃Ⅰ抗体4℃孵育过夜,TBST洗膜3次,每次10 min。辣根过氧化物酶标记的二抗室温孵育2 h后经ECL显色。采用Image J软件对条带的灰度值进行分析,以GAPDH为内参计算各组目标蛋白的相对表达量。
1.2.3 慢病毒转染 将circUTRN24编码序列通过酶切连接插入到circRNA表达载体pLV⁃circ⁃puro,将空载体pLV⁃circ⁃puro及circRNA表达载体pLV⁃circUTRN24分别包装为慢病毒。感染前,将LX⁃2细胞按约5×105个/孔的密度接种于6孔培养板中并加入2 mL DMEM培养24 h。将表达circUTRN24的慢病毒转染LX⁃2细胞,并以空白载体作为对照组,继续培养48 h后裂解细胞并提取总蛋白。
1.3 统计学处理
应用GraphPad Prism 9.0软件进行绘图及统计分析。计量数据以均数±标准差()表示,采用独立样本t检验进行两组间比较,相关性分析采用pearson检验,r为相关系数。以P<0.05为差异具有统计学意义,所有检验P值均为双侧。
2 结果
2.1 胆道闭锁肝组织中circUTRN24及自噬相关蛋白的表达
通过RT⁃qPCR实验分别检测10例BA组和10例CC组的肝组织中circUTRN24的表达。结果显示circUTRN24在BA患儿的肝组织中表达水平明显较高为13.59±6.74,而CC组仅为1.16±0.66,两组相差约12倍,P<0.0001具有显著的统计学差异(t=5.512,见图1A)。同时我们还对两组的肝组织中自噬相关蛋白Beclin⁃1的表达情况进行了Western blot检测。结果如图一B、C所示,自噬相关蛋白Beclin⁃1在BA组肝组织中的相对表达水平为1.06±0.19高于CC组0.64±0.20的1.6倍,且具有统计学差异(t=4.517,P=0.0003)。
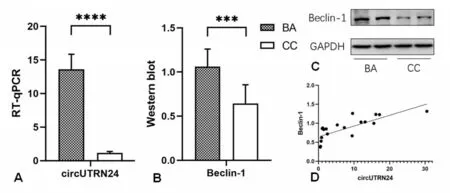
图1 circUTRN24及自噬相关蛋白Beclin⁃1在BA肝组织及对照组中的表达及相关性分析A:circUTRN24的RT⁃qPCR结果,B、C:Beclin⁃1的WB结果,D:二者线性相关分析
2.2 胆道闭锁肝组织中circUTRN24与自噬相关蛋白的表达呈正相关关系
我们将从BA组和CC组的肝组织中获得的circUTRN24与自噬相关蛋白Beclin⁃1的相对表达水平分别进行pearson线性相关分析(见图1D)。结果发现circUTRN24的表达水平与Beclin⁃1呈正相关关系(P<0.0001,r=0.7767)。因此我们推测在胆道闭锁肝组织中circUTRN24表达的升高可能通过自噬途径参与肝纤维化的进展。
2.3 circUTRN24转染LX⁃2细胞促进了自噬相关蛋白及HSC活化相关蛋白的表达
通过慢病毒感染使LX⁃2细胞过表达circU⁃TRN24并与空载体组作对照,过表达circUTRN24的LX⁃2细胞中自噬相关蛋白Beclin⁃1和LC3⁃Ⅱ的表达明显升高,同时HSC活化相关蛋白α⁃SMA和COL⁃Ⅰ的表达水平也明显上调。其中Beclin⁃1、α⁃SMA和COL⁃Ⅰ分别升高了约1.9、1.4和1.6倍;另外实验组LC3⁃Ⅱ表达高于对照组的1.6倍,并且实验组中LC3⁃Ⅱ/LC3⁃Ⅰ的比值约为3倍明显高于对照组的1.4倍(见图2)。这提示circUTRN24的表达可能促进了LX⁃2细胞的自噬过程,并可能通过自噬途径促进HSC的活化参与了胆道闭锁的肝纤维化进展。
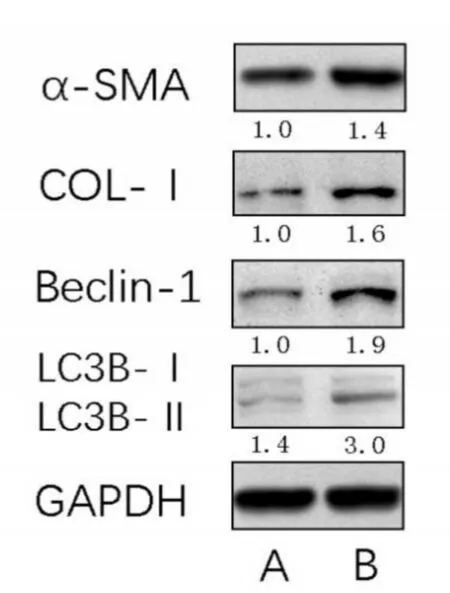
图2 circUTRN24转染LX⁃2细胞后自噬相关蛋白及HSC活化相关蛋白的表达
3 讨论
持续进展的肝纤维化成为胆道闭锁临床诊疗一道无法解决的难题。Kasai术给胆道闭锁的治疗带来了希望,但尽管使胆汁复流也仍然没能阻止大多数患儿肝纤维化的进程,最终仍需接受肝移植治疗[11]。肝纤维化是由各类肝脏细胞、细胞因子和调控介子等经过复杂的相互作用而产生的后果。随着近年来人们对自噬的关注发现其与肝纤维化的进展关系密切,有研究证明自噬可以通过脂质动员和线粒体氧化等方式为HSC的活化提供能量从而促进纤维化的发生[12]。细胞自噬是由自噬相关基因(包括LC3、Beclin⁃1、PI3K等超过30个基因)及其编码的蛋白分子家族所介导的,通过降解细胞内蛋白及细胞器维持细胞内环境稳态的代谢过程,并受到多种因素的影响和调节[13]。
circRNA是近年来备受瞩目的一类RNA家族的新成员,特殊的闭合环状结构决定了其具有较高的稳定性和组织特异性。同时circRNA拥有较长的碱基序列,富含多个功能区域,这赋予了其多样的生物学功能[14]。circRNA曾一度被当作没有功能的转录噪声为人们所屏蔽,随着高通量RNA测序技术的进步,人们发现circRNA在各种疾病和生物学过程中发挥着重要的调控作用。最近的一篇综述报道了,circRNA可以通过转录及转录后水平参与细胞自噬多个阶段的调控[15]。还有报道发现circRNA⁃012091/PPP1R13B可通过自噬介导硅肺病的肺纤维化反应[16],circMTO1可以通过调控miR⁃17⁃5p/smad7抑制肝纤维化[17],另外人们还发现在CCl4小鼠肝纤维化模型中circRNA表达谱存在着明显的变化[18]。这些提示我们circRNA与自噬和肝纤维化可能存在着复杂的关联。本研究组根据前期对胆道闭锁肝组织进行的circRNA高通量测序结果[8],筛选出可能与自噬通路相关的在BA中特异性表达的circUTRN24。故本研究进一步通过对人肝组织研究与体外人HSC细胞培养,探究circUTRN24与细胞自噬和BA肝纤维化的关联及作用机制。
在本研究中,我们首先通过RT⁃qPCR验证了circUTRN24在BA肝组织中高表达,并且应用Western blot检测发现BA肝组织中自噬相关蛋白Beclin⁃1的表达水平亦明显增高。与我们结果相似的是,有研究发现与胆汁淤积症患儿相比,胆道闭锁肝组织中自噬相关蛋白ATG5、ATG7、LC3、PHB2和SQSTM1的水平均有升高[19]。这提示自噬过程的活化可能在BA肝纤维化机制中起到了关键的作用。我们进一步通过线性相关分析,发现Beclin⁃1表达与circUTRN24呈正相关关系。这说明circUTRN24可能参与了自噬过程的活化,为进一步验证这一结果我们进行了后续的细胞水平研究。
肝星状细胞(HSC)的活化目前被认为是肝纤维化的关键节点。HSC是一类间充质细胞多处于静息状态,当受到信号刺激而被激活后,HSC可以表达α⁃平滑肌肌动蛋白(α⁃SMA)以及Ⅰ、Ⅲ型胶原纤维(COLⅠ/Ⅲ),使肝细胞外基质过度堆积导致纤维化的发生。本研究为探讨肝纤维化的机制,特选择具有肌成纤维细胞样表型的永生化人HSC细胞系LX⁃2作为研究载体。基于肝组织水平的研究结果,我们通过慢病毒转染使LX⁃2细胞中过表达circUTRN24,以探索circUTRN24对细胞自噬状态及纤维化指标的影响。研究结果显示,circUTRN24的表达上调了细胞中Beclin⁃1的表达,这与BA肝组织中的结果相一致。同时circU⁃TRN24的表达还增加了细胞中LC3⁃Ⅱ与LC3⁃Ⅰ的比值,这提示了细胞自噬水平的升高,但在BA肝组织中LC3B的比例变化无明显的统计学差异,这可能与肝组织细胞成分复杂,不同细胞间自噬水平差异较大有关。另外对HSC活化指标的检测结果显示,α⁃SMA和COL⁃Ⅰ的表达水平也随circU⁃TRN24的表达而升高,这反映了上调的circU⁃TRN24可以促进LX⁃2的活化。
综合以上结果我们得出推论,circUTRN24可能通过调控HSC的自噬状态而使其活化,最终促进了胆道闭锁肝纤维化的进程,这为胆道闭锁肝纤维化的治疗开辟了新的思路。
猜你喜欢
材料与冶金学报(2022年2期)2022-08-10 09:15:38
传染病信息(2022年3期)2022-07-15 08:24:28
云南医药(2021年3期)2021-07-21 05:40:30
肝博士(2021年1期)2021-03-29 02:32:16
作文成功之路·小学版(2020年6期)2020-07-27 01:48:28
医学研究杂志(2015年6期)2015-07-01 17:40:08
肝胆胰外科杂志(2015年5期)2015-02-27 11:12:44
西南军医(2014年5期)2014-04-25 07:42:38
中国中医药现代远程教育(2014年13期)2014-03-01 04:26:57
无机化学学报(2014年10期)2014-02-28 17:33:13
