
太行山区栓皮栎、刺槐和侧柏对干旱的适应策略研究
2022-11-29 07:07:06牛存洋寿文凯杨喜田张志华乔永胜
林业科学研究 2022年6期
牛存洋,寿文凯*,杨喜田,张志华,乔永胜
(1.河南农业大学林学院,河南 郑州 450002;2.国有济源市南山林场,河南 济源 459000)
水分短缺是太行山低山丘陵区的典型特征。干旱缺水问题严峻,使得当地植被生长缓慢,稳定性差,甚至发生衰退死亡事件[1-2]。研究表明,干旱条件下,木质部水分传输失败(简称水力失败)是造成树木衰退或死亡的重要原因。尽管已有大量关于太行山区不同树种对干旱胁迫适应策略的研究[3-6],但是从水力结构角度研究不同造林树种对干旱的适应策略较少,而且干旱诱导的不同造林树种气穴化栓塞(cavitation and embolism)的水分生理机制尚未明晰。
植物水力结构特征可以反映植物对干旱胁迫的适应策略,通常用植物木质部导水率(Kh)、木质部比导率(Ks)、叶比导率(Kl)、胡伯尔值(HV)等功能性状来表示植物的水力结构特征。研究表明,Ks与导管直径的4次方成正比,这说明植物木质部导管越粗的树种输水效率高于具有较细导管的树种[7];同时,木质部导管直径越粗,越容易发生气穴化栓塞。因此,植物木质部水分传输效率和抵抗气穴化栓塞的能力之间存在权衡关系[8]。根据气种假说,干旱胁迫下木质部气穴化发生程度主要与木质部导管或管胞壁上的纹孔解剖结构有关[9-10]。常绿针叶树种纹孔膜结构由纹孔塞和纹孔缘2部分组成:纹孔塞位于纹孔膜中心位置,密度较大,孔隙度较小;而纹孔缘是环绕纹孔塞的多孔性膜,孔隙度较大,对树液流动的阻力较小[11]。落叶阔叶树种纹孔膜密度较均一,没有塞、缘结构,不同树种的孔隙度存在差异。其次,不同的落叶阔叶树种或常绿阔叶树种,其木质部纹孔开口面积、纹孔开口比例、纹孔密度以及单位面积导管或管胞壁上纹孔面积越大,干旱胁迫下越容易产生气穴化栓塞[12-14];另外,不同的落叶或常绿阔叶树种木质部气穴化发生程度与纹孔膜厚度、弹性以及纹孔膜上的孔隙度也有关系,纹孔膜的厚度越大,越难被拉伸,气穴化导管内的气体进入相邻导管突破纹孔膜遇到的阻力越大;纹孔膜上的孔隙越小,气体越不易进入邻近管腔[15]。
刺槐(Robinia pseudoacaciaL.)、栓皮栎(Quercus variabilisBlume)和侧柏(Platycladus orientalisL.Franco)是太行山地区主要的造林树种。近年来,栓皮栎、侧柏长势良好,而刺槐常在春、夏两季出现落叶、枯梢现象[4,16]。王林等[1]和靳欣等[4]利用控制实验探讨了不同梯度的干旱对刺槐、栓皮栎和侧柏幼苗水力结构和生长的影响,表明严重干旱条件下,水力失败和碳饥饿交互作用加重了干旱对刺槐幼苗的影响,刺槐在水分充足时迅速进行光合积累,而干旱时,通过气孔关闭甚至落叶来避免过度失水。王谦等[2]从太阳辐射、气温、土壤湿度和风速4因子的气候空间角度,解释了高温干旱胁迫下栓皮栎幼苗死亡的机制。赵勇等[17]探讨了太行山区不同造林树种光合水分利用特征,表明栓皮栎是属于高蒸腾高水分利用类型,光合速率日进程属于单峰型;刺槐和侧柏属于双峰型。虽然不同学者从不同角度探讨了不同造林树种的适应策略,但是大多研究倾向于幼苗水平的控制实验,无法反映自然干旱状况下成龄树种对干旱的适应策略,而且对不同造林树种抵抗干旱诱导气穴化的生理学机制尚缺乏解释。本研究以太行山区成龄树种栓皮栎、刺槐和侧柏为研究对象,从水力结构角度探讨不同树种水分传输效率、干旱诱导的气穴化栓塞以及木质部导管和纹孔解剖结构特征等功能性状,探讨不同树种抵抗干旱诱导气穴化栓塞的策略,揭示不同树种抵抗干旱诱导气穴化的机制。
1 材料与方法
1.1 研究区概况
研究地位于太行山南麓小浪底森林生态系统定位研究站附近,35°01′ N,112°28′ E,海拔约400 m,气候属暖温带大陆性季风气候,年平均降水量617.23 mm,7—9月降雨量占全年降雨量的60%左右。年平均温度11.36 ℃,7月平均气温26.2 ℃。1月平均气温-0.5 ℃。日均温≥10 ℃的积温为3 600~4 400 ℃。全年无霜期平均215 d。冬季寒冷干燥,夏季温热少雨。土壤多属碳酸盐褐土和红黏土,土壤有机质含量在0.9%以下,供氮能力低,速效磷缺乏,特别是红黏土,质地黏重,土壤通气性不良。土壤主要由花岗片麻岩等土壤母质发育而成,砾石含量大,保水能力较差。植被区系为暖温带阔叶林区,垂直地带特征不明显。由于天然植被退化严重,此地区多为人工林树种和灌木组成的人工林生态系统,森林主要是刺槐、栓皮栎和侧柏形成的人工纯林。
1.2 试验材料
本试验在太行山南麓南山森林公园开展,于2020年7月选取树龄为30~42 a左右的成龄树种刺槐、栓皮栎和侧柏为研究对象,各个树种的基本信息见表1。此试验为消除立地条件差异引起的试验误差,在海拔、坡度、坡向和坡位尽量一致,各树种生长地靠近的区域设置样地;其次,选取由各个树种组成的纯林为试验样地,每个树种的纯林密度相似(约2 000 株·hm-2),从而消除不同密度栽植的树木给试验数据带来的不确定性。每个树种选择6棵树作为重复,于凌晨4:00—5:00(太阳升起之前),在5~6 m左右的高度,用高枝剪剪取1个长1 m,直径6~8 mm的阳生枝条作为试验材料,随后立即用装满清水的喷壶将枝条的叶片和枝干喷湿减少蒸腾,并放入黑色塑料袋保持湿润,立即带回实验室处理。在水下用枝剪在枝条中间茎段剪取1段长20 cm,直径6 mm左右茎段作为导水率测定的材料,剪取后浸没在水中。将剪取茎段后剩下的枝条末端的叶片和茎干留下,作为茎木质部解剖特征和叶片面积测定的材料。
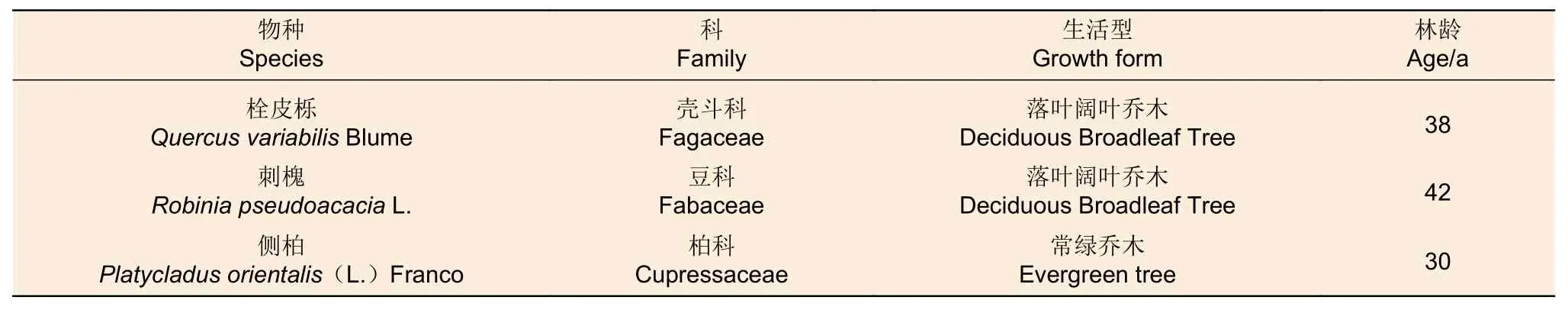
表1 3个树种的基本信息。Table 1 Information of the three tree species used in the present study.
1.3 木质部水分传导率的测定
将取下的长20 cm的茎段用来测定木质部水分传导率。配置浓度为20 mmol·L-1的KCl溶液,将KCl溶液液面和实验平台之间设置50 cm高度差(产生0.005 MPa压力),KCl溶液通过枝条木质部导管流入移液管,通过记录移液管内液体通过的刻度和时间,可以得到溶液通过枝条木质部的流速(单位时间内流过液体的体积,Jv),枝条木质部导水率(Kh/(kg·m·s-1·MPa-1))的计算方法[18]如下:
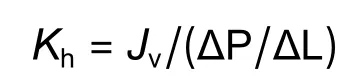
式中:Jv为通过该段枝条的水流速度;∆P为50 cm高的水柱所提供的压力;∆L为该枝条的长度。
利用冲洗法[18]获取栓皮栎和侧柏木质部最大导水率。对测完初始导水率的枝条加压(0.1 MPa),使KCl溶液通过枝条木质部快速流出,冲洗15~20 min,再一次测定木质部导水率(方法同上),即为枝条木质导管的最大导水率(Khmax/(kg·m·s-1·MPa-1))。将测定完初始导水率的侧柏茎段浸没在装满蒸馏水的容器中,置于真空罐中保持真空状态,利用真空方法[19]去除茎段气穴化管胞内的气体,24 h后取出所测导水率即为侧柏最大导水率。
干旱诱导的气穴化栓塞以木质部导水率丧失百分比(PLC/%)反映,计算公式如下:
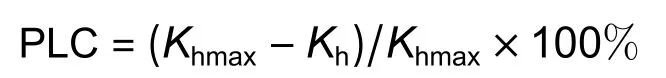
测定完木质部导水率后的茎段用来测定边材面积。用游标卡尺测定茎段两端直径和木材髓心直径,根据圆面积公式计算茎段横截面面积和髓心面积,然后相减得到茎段木质部边材面积(SA/m2)。木质部比导率(Ks/(kg·m-1·s-1·MPa-1))计算公式为:
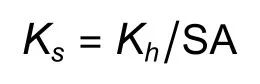
木质部最大比导率(Ksmax/(kg·m-1·s-1·MPa-1))计算公式为:

利用扫描仪扫描枝条末端所有的叶片,并用Image-J计算叶面积大小,得到枝条末端叶片面积和(Total Leaf Area/m2),叶比导率(Kl/(kg·m-1·s-1·MPa-1))计算公式为:
胡伯尔值(HV)计算公式为:

1.4 木质部解剖结构特征和木材密度的测定
将测定木质部水分传导率的茎段用来做木质部解剖。用枝剪将茎段剪成2~3 cm长的茎段,剥去树皮,每个树种6个重复。利用切片机(YD-202)将茎段做横切,切取厚度为22 μm的薄片,利用甲基苯胺蓝(1%)染色。放置在10倍或40倍光学显微镜(Leica ICC50,Wetzlar,Germany)下,拍摄木质部边材最近3年生长年轮,确保视野内至少有100个导管或管胞,便于计算导管或管胞的直径和密度。用Image-J图像处理软件计算得到单个导管或管胞的面积,将每个导管或管胞看作近似圆形,从而计算出导管或管胞的半径(r/μm),导管平均直径(Dv/μm),平均水力直径(Dh/μm)的计算公式[20]如下:
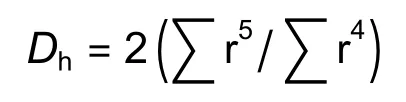
导管密度(VD/(no.·mm-2))为单位视野面积内导管的数量。
切取横切片后的茎段继续做茎段纵切,以获取导管或管胞壁纹孔数量、纹孔开口大小、开口形状等性状。利用切片机(YD-202)切取20 μm厚的纵切片,然后依次放入配置好的浓度为30%、50%、70%、80%、90%、100%的酒精溶液中浸泡10 min,取出晾干8 h确保样品完全干燥,带到中心实验室,将导电胶粘贴在样品桩上,喷金后带到电镜室进行观察。观测时,放大倍数约为7000~10000倍。每个树种选6个茎段作为重复,每个茎段切取1~2片,每个树种至少拍摄15~20张照片,计算纹孔密度(Np),选择至少30个纹孔个体计算单个纹孔膜面积(Ap)、单个纹孔开口面积(Aap)、纹孔开口比例(Fap=Aap/Ap)、纹孔开口形状参数(APf,纹孔口最短轴比最长轴)、纹孔密度(Np,单位面积导管壁纹孔数量)、纹孔比例(Fa,单位面积导管壁总纹孔面积比例)等性状[12]。
每个树种选取6个长1~2 cm茎段泡入水中,剥去树皮,将髓心去除。利用排水法测定湿润边材的体积V,然后将枝条放入烘箱内60 °C烘干72 h至恒质量,用万分之一电子天平(CPA225D,Sartorius Inc.,Göttingen,Germany)称量记录为M,最后得到木材密度(ρwood/(g·cm-3)),计算公式为:
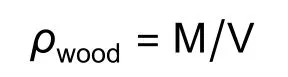
1.5 数据分析
利用Sigmaplot(10.0)软件作图,利用SPSS(20.0)软件进行数据统计分析,利用单因素方差分析(one-way ANOVA)不同树种之间功能性状差异,利用LSD进行多重比较。
2 结果与分析
由图1可以看出,栓皮栎、刺槐和侧柏木质部水力结构存在显著差异。栓皮栎和刺槐的木质部初始比导率和最大比导率均显著高于侧柏(p<0.05),刺槐和栓皮栎之间无显著差异,表明栓皮栎和刺槐水分传输效率高于侧柏。栓皮栎叶比导率显著高于侧柏,刺槐叶比导率和其它2个树种差异不显著。侧柏胡伯尔值(HV)(2.15×10-4)显著高于栓皮栎(1.10×10-4)和刺槐(8.34×10-5),表明侧柏维持每单位叶面积水分供给所需的茎干组织较粗,而栓皮栎和刺槐维持单位叶面积水分供给所需的茎干组织较细。由图2可知:刺槐木质部导管平均直径(Dv)和导管水力直径(Dh)均显著高于栓皮栎和侧柏,栓皮栎Dv和Dh显著高于侧柏(图2a、b,p<0.05)。3个树种Ks的变化趋势和Dv、Dh的变化趋势一致,表明导管直径越宽,Ks越大。木质部导管密度(VD)(图2c)侧柏(3 656.03)>栓皮栎(47.05)>刺槐(17.21),3树种导管密度之间差异显著(p<0.05),且3树种的VD变化趋势和Dv、Dh的变化趋势相反,表明导管直径越宽,导管密度越低。3树种木材密度之间差异不显著(图2d)。
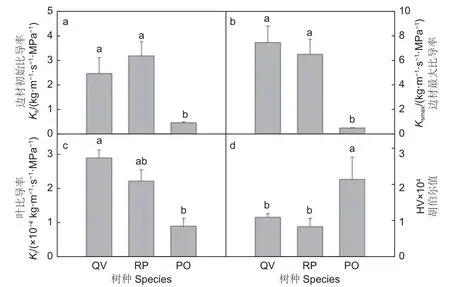
图1 3个树种的枝条木质部水力结构Fig.1 Branch xylem hydraulic architecture of three tree species
由表2和图3可以看出:3树种木质部导管或管胞壁纹孔形态和结构性状特征存在明显差异。形态方面,刺槐纹孔室和纹孔口具有点状和齿状附物,栓皮栎和侧柏纹孔室没有附物,侧柏纹孔膜中央显示加厚纹孔塞结构(图3)。纹孔膜面积比较,刺槐>侧柏>栓皮栎,刺槐纹孔膜面积是栓皮栎的1.70倍,是侧柏的1.001倍。纹孔开口面积刺槐>栓皮栎>侧柏,刺槐纹孔开口面积显著高于栓皮栎和侧柏,刺槐纹孔开口面积是栓皮栎的1.92倍,是侧柏的2.12倍。纹孔开口比例刺槐>栓皮栎>侧柏,3树种纹孔开口比例均差异显著。纹孔口形状参数侧柏>栓皮栎>刺槐,侧柏纹孔开口形状接近圆形。纹孔密度刺槐>栓皮栎>侧柏,3树种纹孔密度差异显著,刺槐纹孔密度是栓皮栎的1.24倍,是侧柏的3.65倍。木质部单位面积导管或管胞壁上所有纹孔的面积刺槐>栓皮栎>侧柏,3树种之间差异显著(p<0.05),刺槐纹孔面积占比是栓皮栎的2.19倍,是侧柏纹孔面积占比的4.28倍(表2)。刺槐的纹孔面积等性状显著高于栓皮栎和侧柏,刺槐的Dv和Dh也显著高于栓皮栎和侧柏(图2),表明导管直径越宽,导管壁上的纹孔膜面积、纹孔开口面积、纹孔密度等越大。此外,3树种木质部导水率丧失百分比(PLC)栓皮栎(67.03%)>刺槐(50.90%)>侧柏(5.12%)(图4),3树种木质部气穴化程度存在显著差异(p<0.05),表明干旱条件下,栓皮栎更容易发生气穴化栓塞,刺槐次之,侧柏抵抗干旱诱导气穴化栓塞的能力最强。
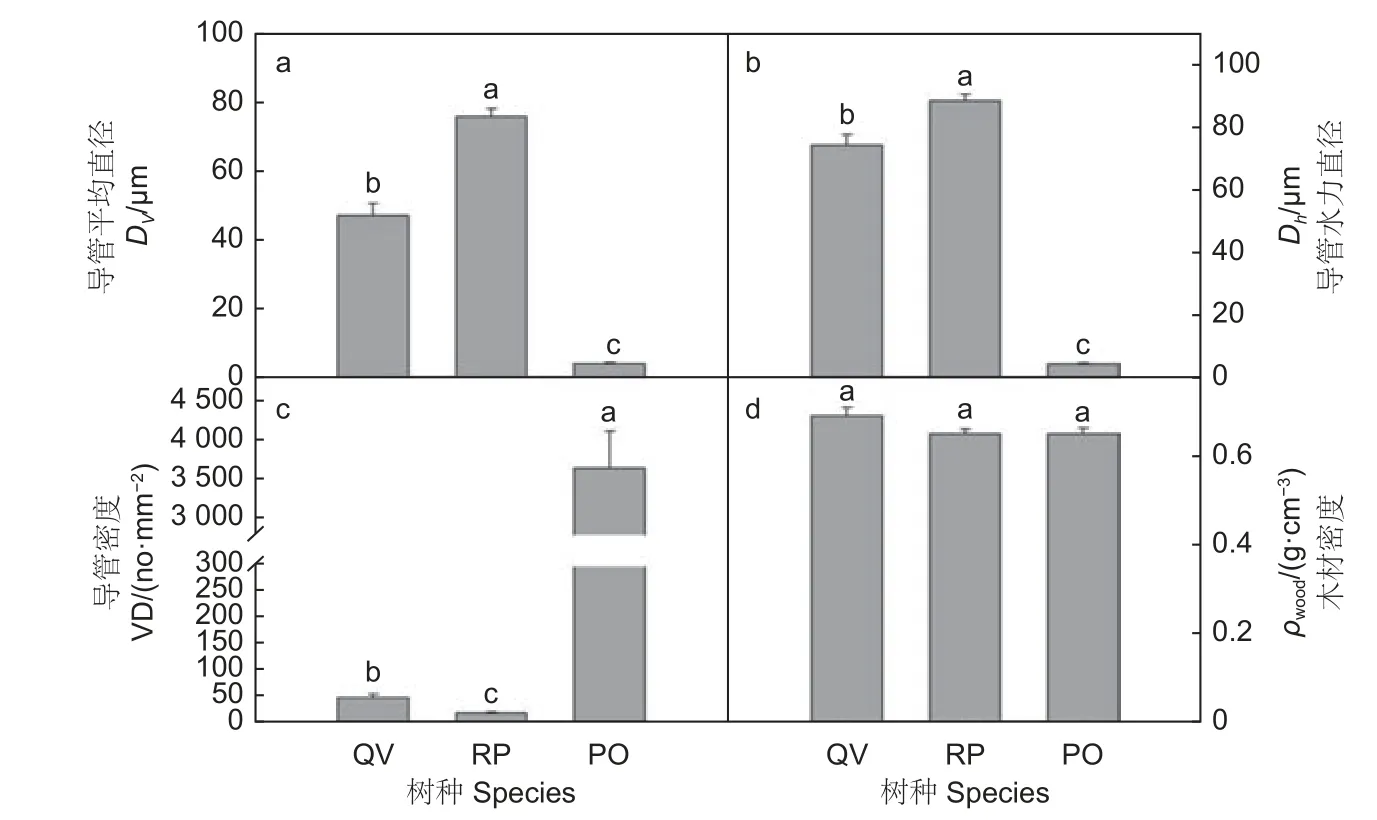
图2 3个树种茎段木质部解剖结构Fig.2 Stem xylem anatomy of three tree species
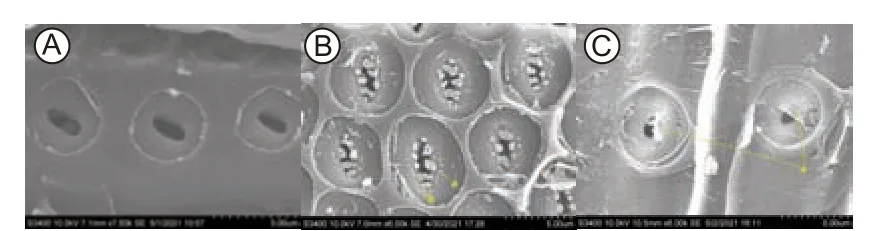
图3 3个树种木质部扫描电镜图像Fig.3 Scanning electron anatomical pictures of three tree species
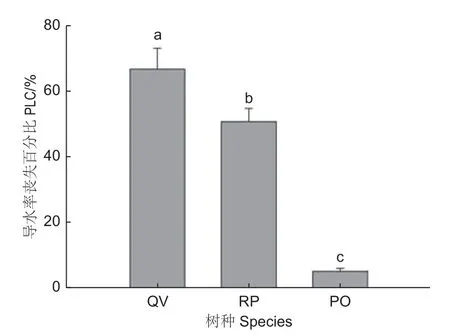
图4 3个树种茎段木质部导水率丧失百分比Fig.4 Stem xylem percent loss of hydraulic conductivity of three tree species

表2 3个树种木质部纹孔解剖结构特征Table 2 Pits functional traits of three tree species
3 讨论
参考前人的研究,本课题组提出以下假设:(1)与侧柏相比,栓皮栎和刺槐木质部具有较宽导管和较高的枝条比导率和叶比导率以及较大的纹孔开口面积,干旱胁迫下木质部导管内产生大量的气穴化栓塞,且栓塞程度高于侧柏;与栓皮栎相比,刺槐可能具有更粗的导管或更大的纹孔开口面积等性状,所以干旱胁迫下刺槐木质部产生更多的气穴化栓塞。(2)3个树种木质部较宽的导管或管胞可维持较高的水分传输效率,但是干旱会诱导发生更多的气穴化栓塞,因此,体现枝条木质部水分传输的有效性和抵抗干旱诱导气穴化能力之间的权衡关系;不同树种具有不同的水力结构,表现出不同的适应干旱的策略。
太行山区3种造林树种木质部水力结构存在显著差异。3个树种枝条木质部比导水率(Ks)(图1a)和Dv、Dh(图2a、b)变化趋势一致,表明导管或管胞直径越大,木质部水分传输效率越高,这与Hacke等[21]的研究结果相符。因为根据Hagen-Poiseuille方程,导水率与导管直径的4 次方成正比[21]。3个树种PLC排序栓皮栎>刺槐>侧柏(图4),即干旱胁迫下,栓皮栎和刺槐木质部抵抗气穴化的栓塞能力比侧柏更弱,这支持了本研究第一个假说。栓皮栎和刺槐的Dv和Dh也显著高于侧柏,表明木质部导管直径越宽,其对干旱诱导气穴化的抵抗能力越弱,这与前人的研究一致,不管是在树种个体水平,还是器官水平上,具有较大导管或管胞直径的树种或器官比具有较小导管或管胞直径的树种或器官更容易发生气穴化栓塞[12,22]。木质部较宽的输水管道可以支持较高的水分传输效率,但却增加了干旱胁迫下发生气穴化栓塞的风险,表明木质部水分传输效率与抵抗气穴化栓塞能力之间存在权衡关系[23]。
其次,3个树种多个纹孔性状和导管性状大小排列趋势一致(图2,表2),且导管和纹孔开口较大的刺槐和栓皮栎PLC显著高于侧柏,表明木质部导管或管胞性状与纹孔结构性状具有协同关系,协同影响木质部水分传输效率和安全性[24]。这与其他学者对杨属的研究结论一致,在组织水平上,杨属较宽的导管次生细胞壁上纹孔面积占据很大比例,这也使得出现罕见的比较大的纹孔的概率增加,进而增加了发生气穴化栓塞的风险[25-26]。此外,侧柏木质部导管壁APf最大(表2),侧柏PLC最小,栓皮栎和刺槐APf较小,但PLC均较高(图4),表明纹孔开口短轴越短,长轴越长,其抵抗气穴化的能力越弱。这与Scholz等[27]的研究一致,其研究结果表明10种李属树种木质部APf越大,抵抗干旱诱导气穴化栓塞能力越强。刺槐纹孔膜面积、纹孔开口面积、纹孔密度及纹孔比例等性状均显著高于栓皮栎,但刺槐PLC略低于栓皮栎(表2,图4),这一结果不支持水分传输有效性和安全性之间存在权衡,且与“气种假说”理论相矛盾,干旱条件下,木质部导管或管胞内气穴化的形成主要取决于相邻管壁连接处纹孔的超微解剖结构,纹孔开口比例或纹孔密度越大,越容易使气泡扩散至相邻管腔形成栓塞[28]。刺槐PLC低于栓皮栎,这可能与刺槐纹孔室和纹孔口存在的点状和齿状附物有关,因为研究表明附物例如螺旋形加厚的沟槽能增加导管壁的表面积,增强水分与细胞壁结合,增加水柱张力而不破损,增加了其对干旱胁迫诱导气穴化的抵抗能力[24,29]。也有学者认为,附物纹孔的主要化学成分是木质素和多糖,木质素能提供负压下纹孔膜拉伸和偏转的机械缓冲作用,多糖具有水分聚合力,二者均能减弱干旱状况下气穴化发生的风险[30-31]。通过扫描电镜发现,刺槐纹孔属于附物纹孔,其纹孔室发现点状附物,纹孔口发现齿状附物(图3),这些不同形态的附物可能一定程度上缓冲了干旱胁迫下纹孔膜的拉伸,增加了细胞壁表面积,进而削弱了气穴化导管气体向相邻导管扩散,增强刺槐抵抗干旱诱导气穴化的能力,未来更多的研究应该尝试研究更多具有附物纹孔的树种,并量化纹孔附物的形态、分布及密度和化学组成成分等,从而揭示附物纹孔树种抵抗干旱诱导气穴化的机制。
栓皮栎、刺槐和侧柏共存同一地区,表明其具有不同的生态位,对干旱环境有不同的适应策略。栓皮栎、刺槐和侧柏由于木质部解剖结构的差异,导致其水分传输效率和抵抗干旱诱导气穴化能力产生差异。侧柏管胞直径较小(图2a、b),纹孔膜厚度不均匀,中间具有加厚的纹孔塞结构(图3),水分充足条件下,允许水分经过纹孔膜孔隙流通,但在干旱胁迫发生时,纹孔对纹孔膜两侧压力差增大,导致纹孔膜发生拉伸和偏转[32],纹孔塞覆盖纹孔口,可有效阻止更多的空气向相邻管胞的扩散,因此,侧柏抵抗干旱诱导气穴化能力较强(图4);但侧柏较安全的纹孔结构同时也限制了木质部的水分传输效率,侧柏水分传输效率显著低于栓皮栎和刺槐(图1),表明侧柏具有更保守的资源获取和利用策略。栓皮栎和刺槐其木质部导管直径较宽,纹孔开口面积、纹孔开口比例和纹孔比例等都较大(表2),干旱胁迫下,纹孔对两侧压力差增加造成纹孔膜发生偏转后,纹孔膜孔隙被拉伸变大,空气更容易向相邻导管内扩散,因此,栓皮栎和刺槐抵抗干旱诱导气穴化能力较弱。但栓皮栎和刺槐较宽的导管和较大纹孔膜及纹孔口等结构特征支持更高的水分传输效率(图1),可以在生长季快速获取资源实现快速生长,具有快速生长的策略。另有研究表明,木质部水分传输效率和叶片光合速率之间存在显著协同关系[33],因此,侧柏与栓皮栎和刺槐对干旱环境不同的适应策略也可从叶片光合水分利用特征上得到验证。如赵勇等[17]研究表明,栓皮栎和刺槐叶片水分利用效率和光合速率明显高于侧柏,支持侧柏具有保守资源获取与利用策略,栓皮栎和刺槐具有较快的资源获取能力支持快速生长。
关于植物木质部抵抗干旱诱导气穴化栓塞的能力与抗旱性之间的关系仍存在争议,作者倾向于支持木质部抵抗气穴化能力与抗旱性之间关系并不明显这一观点[34]。本研究结果显示,干旱环境下,栓皮栎对干旱诱导气穴化抵抗能力最弱,刺槐次之,侧柏最强,但是栓皮栎和侧柏生长较好,刺槐经常发生枝条枯梢的现象,表明干旱诱导的气穴化栓塞程度高低并不能准确反映不同树种的抗旱能力。不同树种抗旱能力是根、茎、叶等各个器官或组织综合表现的结果,不能仅仅利用枝条水平的气穴化程度来评价某个树种是否更耐旱,根系分布、叶片气体交换能力等都对植物抗旱能力有影响。研究表明,刺槐叶片水分调节对策属于等水调节,干旱胁迫下其通过关闭叶片气孔使木质部水势保持在稳定水平,刺槐春夏落叶或枯梢是与叶片的气孔调节策略有关,为了整株树存活采取的舍弃部分叶片或枝干的策略[35]。栓皮栎叶片调节对策属于非等水调节,其叶片气孔对环境变化不敏感,木质部水势随环境而变化[36]。水力安全边界是评估植物抵抗气穴化栓塞能力的重要性状,通常用枝条或叶片正午最低水势和P50(导水率丧失50%对应的枝条或叶片水势)之间的差值表示,其差值正值越大,抵抗气穴化能力越强。在干旱胁迫下,关闭气孔避免气穴化;而差值是较小的正值或趋于负值的树种,干旱胁迫下不完全关闭气孔,以维持长时间的气体交换,但其气穴化程度较高,而其栓塞恢复力也更强[37]。刺槐水力安全边界值可能是较大的正值,抵抗气穴化能力强,但栓塞恢复能力较差,而栓皮栎虽然气穴化程度较高,但恢复能力较强,因此,干旱环境中,栓皮栎长势比刺槐好,也更抗旱。也有学者通过对刺槐幼苗的干旱复水实验,结果表明,刺槐在反复干旱复水后叶片气孔蒸腾作用变化微弱,水分传输恢复缓慢,导致光合作用速率恢复缓慢,产生碳饥饿,增加了死亡风险[38],而栓皮栎反复干旱以及复水后,能快速恢复水分传输和叶片光合碳同化能力[39],也验证了栓皮栎更抗旱这一观点。
4 结论
(1)与侧柏相比,栓皮栎、刺槐枝条木质部具有较宽的导管和较高的木质部水分传输效率,但是后者产生大量的木质部气穴化栓塞,即栓皮栎和刺槐抵抗干旱诱导气穴化的能力较弱。
(2)虽然刺槐导管直径和纹孔开口、纹孔开口比例等性状均显著高于栓皮栎,但是刺槐PLC显著低于栓皮栎,这可能与刺槐纹孔室和外纹孔口存在点状和齿状附物有关,不同形状附物可增加导管壁表面积,增加水分与细胞壁结合,防止水柱断裂。另外,附物的化学组成成分多为木质素和多糖,可增加水分聚合力,缓冲干旱诱导下纹孔对纹孔膜机械拉伸作用,从而减小气体从气穴化导管向相邻导管腔扩散的可能性,增强刺槐抵抗干旱诱导气穴化的能力。
(3)栓皮栎和刺槐生长季利用较高的水分传输效率快速生长,具有快速生长的策略。侧柏较窄的管胞和纹孔膜的塞缘结构使得侧柏木质部水分传输效率较低,具有比较保守的资源利用策略。木质部导管与纹孔性状协同决定木质部水分传输效率和干旱诱导的气穴化。
致谢感谢河南黄河小浪底关键带国家野外科学观测研究站
猜你喜欢
花卉(2024年1期)2024-01-16 11:33:50
机床与液压(2023年18期)2023-10-12 01:36:42
林业科学研究(2023年4期)2023-04-29 00:44:03
新农民(2022年22期)2022-11-07 23:38:06
机床与液压(2022年13期)2022-09-15 05:12:56
星星·诗歌原创(2021年3期)2021-06-01 09:13:23
农业机械学报(2020年9期)2020-10-10 07:08:02
南方农业·下旬(2020年1期)2020-06-01 07:47:32
现代园艺(2019年24期)2019-01-07 01:19:36
现代农业科技(2009年21期)2018-09-03 00:00:00
