
暖温带马尾松茎干径向变化特征及其对环境因子的响应
2022-11-29 13:13:42师志强章异平罗鹏飞翟乐鑫侯佳玉徐军亮
西北林学院学报 2022年6期
师志强,章异平,罗鹏飞,竹 磊,翟乐鑫,侯佳玉,徐军亮
(河南科技大学 园艺与植物保护学院,河南 洛阳 471000)
树木生长除受自身基因型[1]和生理过程[2]等内源因素的影响外,光照、水分等外源环境条件对树木生长亦有重要影响[3]。研究环境因子和树木生长之间的关系,有利于探讨气候变化对树木生长过程的影响。以往对于树木径向生长的研究主要通过树轮年代学来实现,树木年轮包含了树木每年的气候变化信息,成为研究树木对气候变化响应的重要材料[4-5]。这部分研究通过长时间序列(百年或千年)的树木的生长规律来反映气候变化。然而,树木生长并不匀速,因此与气候因子并非简单的线性关系,而是受生长阶段影响的非线性关系[6]。因此,仅从长时间年际序列来理解环境因子对树木生长的影响存在一定的局限性,有必要从短时间序列来探索树木径向生长与气候之间的相关关系,加深气候变化对树木生长影响的理解。
径向生长测量仪(DENDROMETER)是一种高精度的,高分辨率的,能够自动连续监测树木径向生长的仪器[7-8]。可有效地从树木生理生态学的角度,来分析树木径向生长与环境因子之间的响应,因此近年来被广泛应用[9-10]。21世纪以来,众多国内外学者使用径向生长测量仪来探究树木径向生长与环境因子之间的关系。A.Deslauriers等[11]对加拿大魁北克北方森林的胶冷杉(Abiesbalsamea)径向生长的研究表明,降水是影响树木径向生长的主要环境因子。A.Gruber等[12]对阿尔卑斯山内树种的树木径向变化的研究发现,温度是影响树干径向日变化量的主要环境因子。在芦芽山地区,董满宇等[13]研究表明,土壤温度是影响华北落叶松(Larixprincipis-rupprechtii)径向生长的主要影响因子。同一地区白杄[14](Piceameyeri)在茎干快速生长期,径向生长也受土壤温度的影响。但是在茎干脱水收缩期,土壤含水量对白杄累积茎干径向变化的影响增加。在处于干旱区的祁连山地区,水分是限制树木径向生长的主要因素。但也有研究认为,热带地区的苦楝[15](Meliaazedarach)径向生长同样受水分的限制。由此可知,树木生长与环境因子的关系存在较大的区域差异性。目前相关研究主要集中在热带地区和高山林线地区,对暖温带地区的研究甚少。
马尾松(Pinusmassoniana)是我国南方荒山造林的主要树种之一[16],是中国亚热带地区植被恢复的先锋树种[17],也是亚热带东部地区的乡土树种[18]。由于马尾松年轮界线清晰,它也是我国亚热带地区树轮气候学的主要树种之一[19]。已有研究表明,不同区域条件下的水热组合使得马尾松径向生长对气候的响应存在区域差异。在马尾松分布的南界,侯爱敏等[17]研究表明,夏季高温成为马尾松径向生长的主要限制因子。在分布中心地区[20],生长季早期的温度升高对马尾松径向生长有利,而后期受温度和降水的综合影响。而在北界信阳地区[21],温度不再制约马尾松的生长,水分是影响马尾松径向生长的主要环境因子。这些研究结果均以马尾松轮宽为指标开展的年际间研究,而对马尾松在短时间尺度的年内研究还相对较少[22],且研究区域主要位于亚热带地区。对于年内马尾松和环境因子之间的关系尚不清楚。因此,本研究在马尾松分布北界的暖温带地区(河南洛阳),使用径向生长测量仪对马尾松年内径向生长动态进行了1 a的连续监测。研究马尾松茎干径向变化动态,分析不同时间尺度上(日、月、季节)树木茎干径向变化特征。探讨马尾松径向变化与环境因子的相关关系,旨在为马尾松林的经营管理制定合理措施提供科学依据。
1 材料与方法
1.1 研究区概况
本研究地点位于河南省洛阳市河南科技大学校园内西北角(34°36′N,112°24′E),属于暖温带大陆性季风气候,夏季高温多雨,冬季寒冷干燥,春秋季节较短。选取距离样地最近的国家气象站孟津站多年气象数据(1960-2018)(图1)可知,研究区年均总日照时数为2 198.3 h,年均温14.2 ℃,月最高空气温度26.4 ℃,出现在7月,月最低空气温度0 ℃,出现在1月。年均总降水量620.4 mm,全年63%的降水主要集中在6-9月,其中7月份降水最多,达到140.9 mm。样树周围有悬铃木(Platanusacerifolia)、紫叶李(Prunuscerasifera)、大叶黄杨(Euonymusjaponicus)、圆柏(Sabinachinensis)和石楠(Photiniaserratifolia)等树种。
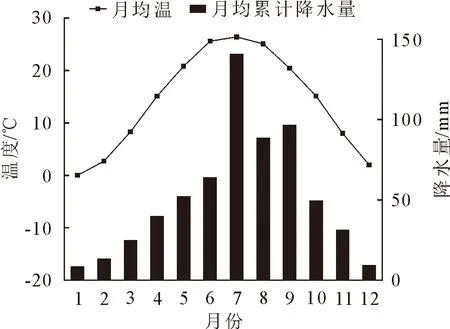
图1 孟津气象站1960-2018年月均温和月均累积降水量
1.2 数据采集和处理方法
1.2.1 马尾松径向生长数据采集处理 选取长势良好,树高和胸径相近的3棵马尾松作为试验对象(表1)。在马尾松树高1.3 m处安装点触式树木径向生长测量仪(Point DENDROMETER,Ecomatik,Germany),安装时应首先剥去树干外部的死树皮,以减少树皮水分变化对树干径向生长监测数据的影响[23]。在2019年6月-2020年5月,持续监测马尾松树干径向变化,以10 min为间隔时间自动记录并保存数据。

表1 马尾松样树基本特征
通常情况下,径向变化测量仪监测树木的径向生长信息包含2部分:一部分是由树木茎干水分波动引起的可逆的收缩与膨胀变化,另一部分是树木茎干形成层细胞分裂与木质部生长引起的不可逆的变化[10]。采用A.Deslaurier等[11]的方法,根据连续5 d的树木茎干径向日变化量的均值与零值的差异来确定生长季的开始日期和结束日期。若两者之间存在显著性差异(独立样本T检验),则确定连续5 d的首日为生长开始日。同理,若两者之间不存在显著性差异,则以连续5 d的首日为生长结束日。同时,依据O.Bouriaud等[24]提出的最大值法,提取茎干日变化量,具体做法为从每日获取的144个树木茎干原始值中,提取最大值,将当日最大值与后一日最大值之差作为当日树木茎干的日变化量,在此基础上加和得到树木累积茎干径向变化量,具体公式如下[25]。
每日茎干径向变化量(ΔR,mm)公式为:
ΔRi=Rmax(i+1)-Rmax(i)
(1)
式中:ΔRi表示每日茎干径向变化量,Rmax(i+1)表示第i+1天的茎干半径的最大值,Rmax(i)表示第i天的茎干半径最大值。
累积茎干径向变化量(Rcum,mm)公式为:
(2)
式中:Rcum表示累积茎干径向变化量,ΔRi表示第i天的茎干径向变化量。
1.2.2 气象数据采集及处理 在样地内的空地上安装小型HOBO-U30自动气象站(ONSET,美国)同步监测气象数据,气象因子包括10 cm和20 cm土壤体积含水量(soil water content,SWC,m3/m3)、10 cm和20 cm土壤温度(soil temperature,ST,℃)、空气温度(temperature,T,℃)、空气相对湿度(relative humidity,RH,%)、太阳辐射(solar radiation,SR,W/m2)等,记录时间间隔与DENDROMETER监测树木径向变化一致均为10 min。降水量(precipitation,P,mm)数据来自中国气象数据网的孟津气象站,网址为:http://data.cma.cn/site/index.html。饱和水汽压差(vapor pressure deficit,VPD,kPa)根据经验公式[26]获得,计算公式如下:
(3)
式中:VPD表示饱和水汽压差,RH表示样地相对空气湿度,T表示样地平均空气温度。
1.2.3 数据处理 首先对取得的实验数据进行初步检验,剔除异常值,为进一步分析数据提供保障。获取日尺度上的土壤体积含水量、土壤温度、空气相对湿度、空气温度和降水量等环境因子,因仪器故障,10 cm土壤体积含水量数据存在部分缺失。采用Pearson相关分析的方法分析马尾松茎干径向变化量与环境因子的相关性。文中数据统计分析利用SPSS 25.0和OFFICE 2019软件完成,图表制作用ORIGIN 2018完成。
2 结果与分析
2.1 不同月份马尾松茎干径向日变化规律
由图2可以看出,在日尺度上马尾松茎干径向日变化规律为0:00-8:00茎干半径持续上升,8:00-16:00持续下降,16:00-次日0时持续上升。树木茎干半径最大值通常出现在8:00,最小值出现在16:00,不同月份之间稍有差异。3-10月,随着树木茎干径向日变化的开始,树木茎干最大值出现的时间逐渐提前,最早出现在6:00。11月至次年2月,最大值出现的时间逐渐推迟,最晚出现在9:00。
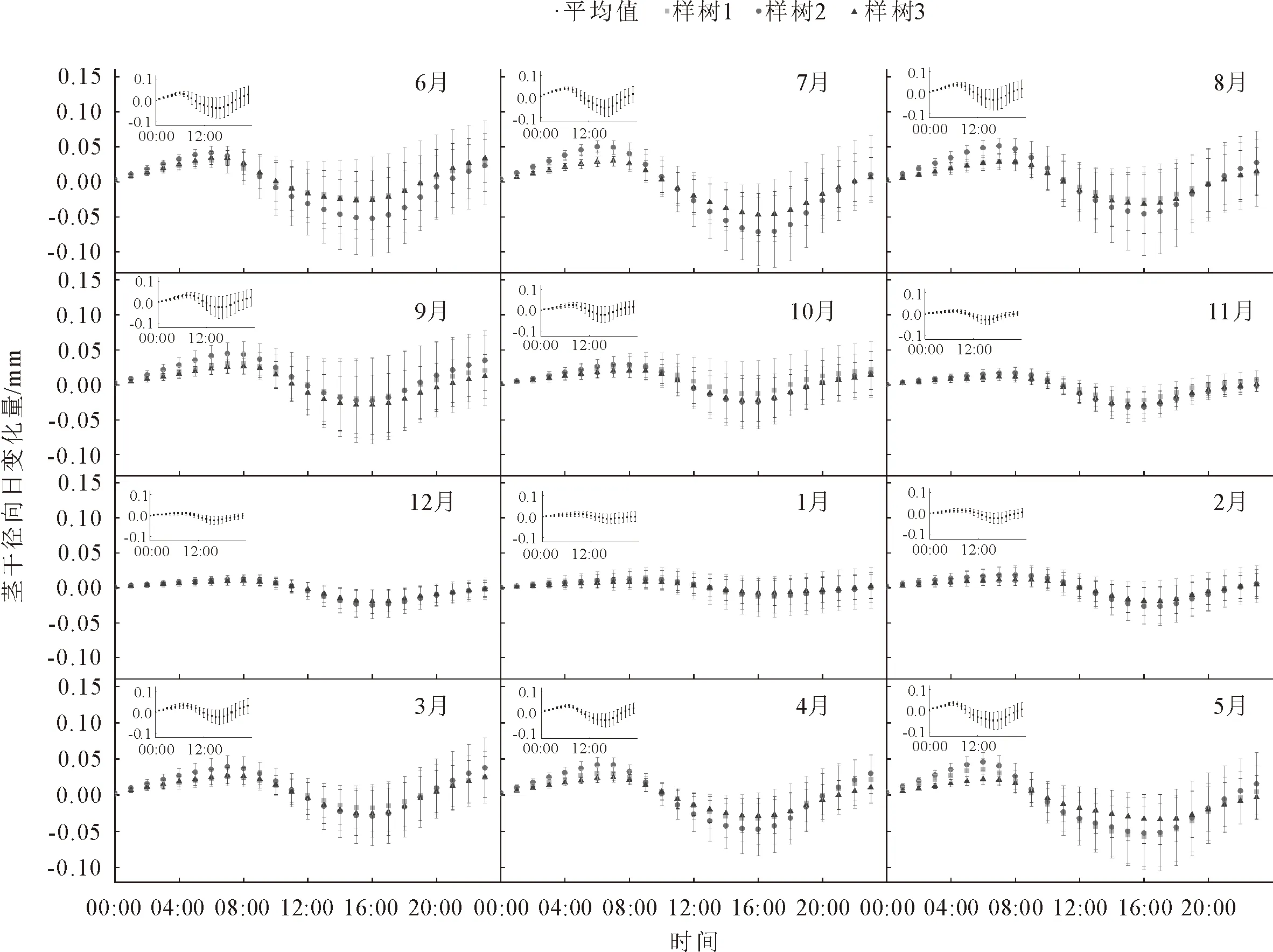
注:左上方小图代表相应月份3棵样树径向日变化均值。
不同月份,马尾松年内茎干径向日变化的振幅在21.9~92.4 μm波动,其中最大振幅出现在7月,最小振幅出现在1月。3-10月径向日变化振幅都较为明显,相比而言,11月至次年2月径向日变化趋于稳定,振幅较小。
2.2 马尾松月累积茎干径向变化量
根据连续5 d马尾松径向生长结果显示(图3),马尾松径向生长开始和结束时间分别为3月4日和11月5日。进一步结合马尾松月累积茎干径向变化量(图4)来看,马尾松年内径向变化特征可以分为3个阶段。第1阶段:春季茎干复水期,3-5月,随着温度的升高,马尾松月累积茎干径向变化量开始明显增加,3月和4月马尾松茎干径向变化量分别达到977.3 μm和722.1 μm,5月,在水热条件的影响下,茎干径向变化量为301.5 μm。第2阶段:夏秋茎干快速增长期。6-10月,马尾松月累积茎干径向变化量的范围为517.7~943.3 μm,累积茎干径向变化量最大的月份为6月,最小的月份为7月。第3阶段:冬季茎干稳定期。11月至次年2月,马尾松累积茎干径向变化量在-23.6~199.9 μm起伏。2月树木茎干会出现迅速变粗,从而导致树干径向变化量出现迅速增加。

注:↓表示马尾松径向生长结束(左)和开始(右)。

注:左上方小图代表相应月份3棵样树的累积径向变化量均值。
2.3 气象因子对马尾松茎干径向变化量的影响
从图5可知,研究区样地年均空气温度为16.5 ℃,月最高空气温度出现在7月为28.5 ℃,月最低空气温度出现在1月为2.8 ℃。年总降水量为668.4 mm,降水最多的月份为8月,达到192.7 mm。总的来说,10 cm和20 cm的土壤温度相差不大,其年均值分别为16.7 ℃和16.8 ℃,但在2019年6-8月和2020年3-5月,土壤温度10 cm大于20 cm。与土壤温度一致,10 cm土壤含水量总体上高于20 cm,其年均值分别为17%和14%,但在6月21-27日,土壤含水量10 cm小于20 cm。2020年5月,日最高空气温度达到28.6 ℃,平均温度24.1 ℃,土壤含水量10 cm和20 cm为整个生长季最低,分别为16%和13%,VPD达到1.20 kPa。
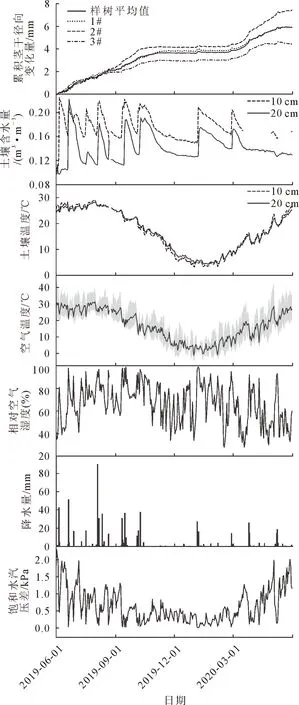
图5 马尾松累积茎干径向变化量及同期环境因子特征
将2019年6月至2020年5月日尺度上马尾松茎干径向变化量与同期环境因子做Pearson相关分析(表2),结果表明,3-5月,土壤温度、空气温度和VPD与马尾松茎干径向变化量均为极显著正相关(P<0.01),而土壤含水量与马尾松茎干径向变化量存在极显著负相关(P<0.01)。6-10月,相关分析结果相反,即土壤温度、空气温度和VPD与马尾松茎干径向变化量的关系由正相关变为负相关(P<0.01),而土壤含水量与马尾松茎干径向变化量的关系由负相关变为正相关(P<0.05)。这说明,生长季初期较高的温度有利于马尾松径向变化量的增加,而过高的温度反而抑制马尾松径向变化量的积累。11月至次年2月,土壤含水量、相对湿度与马尾松径向变化量为极显著正相关(P<0.01),降水与马尾松径向变化量亦有显著正相关(P<0.05),而VPD则与马尾松茎干径向变化量呈现出极显著负相关的关系(P<0.01)。

表2 马尾松日尺度茎干径向变化量与同期环境因子的Pearson相关关系
3 结论与讨论
3.1 结论
利用DENDROMETER监测仪连续监测了分布在暖温带马尾松的茎干径向生长动态。在马尾松分布的北界,马尾松茎干径向日变化规律为昼缩夜胀,8:00为径向变化极大值,16:00为径向变化极小值。马尾松年内茎干径向变化量分为3个阶段:春季茎干复水期,夏秋茎干快速增长期,冬季茎干稳定期。在不同阶段,影响茎干径向变化量的主要环境因子不同。温度是生长季(3-10月)中控制马尾松茎干日变化量的主导因子。
3.2 讨论
3.2.1 不同时间尺度马尾松茎干径向变化量特征 从日尺度来看,马尾松茎干径向日变化量和水分有紧密关系。马尾松茎干径向日变化表现为昼缩夜胀的规律,即在0:00-8:00增大,在8:00达到极大值,之后茎干开始收缩,在16:00缩至当天极小值,之后茎干又继续增大。在生长季,树木根部夜晚开始吸收水分,早上随着太阳升起,树木开始进行蒸腾作用,水分随着茎干运输到树冠部分。此时,蒸腾作用消耗的水分小于树木根部吸收的水分,水分被存储在茎干部分,使得树干半径增加[27]。随着蒸腾作用的增强,运输到树冠部分的水分增加,蒸腾耗水大于根部吸水,树木半径逐渐减小。到了傍晚,蒸腾作用减弱,茎干水分得以恢复,树木半径再次增加。这种树木茎干昼缩夜胀的日变化规律,不仅出现在马尾松,在瑞士阿尔卑斯山中部的欧洲落叶松(Larixdecidua)和挪威云杉[28](Piceaexcelsa);在祁连山地区的华北落叶松[29](L.principis-rupprechtii),祁连圆柏[29](Juniperusprzewalskii)等都有发现。但也有研究表明,树木茎干径向日变化也可能呈现昼胀夜缩的规律,这与树种、海拔、纬度、气温和降水等综合因素有关[30]。
从季节尺度来看(图4),在生长季,马尾松月累积茎干径向变化量存在“双峰”现象,这与刘亚静[31]、李颖俊[32]的研究结果一致。本研究显示,春季(3-5月),随着温度的回升,马尾松径向生长启动,月累积茎干径向变化量发生明显变化。夏秋季节,在6月和9月出现2个峰值,分别为943.3 μm和823.6 μm。冬季,温度降低,树木月累积茎干径向变化量基本为0。2月,随着温度升高,降水增加,月累积茎干径向变化量增加,树干出现迅速增粗的假象[33-34],这一假象很容易被误认为是树木生长[11,35]。事实上,这是由于根部吸水所带来的茎干细胞中水分的变化,而并非生长所带来的茎干细胞生长。这一现象并不是马尾松独有的,在樟子松[30](Pinussylvestris)、红松[36](Pinuskoraiensis)、青杄[23](Piceawilsonii)和落叶松[13](Larixgmelini)等树种的研究中,均有发现。
3.2.2 马尾松茎干日变化量与环境因子的关系 树木生长受环境因子、树木生理特性等多因素的共同影响[37]。因此,树木径向生长对气候变化的响应在不同生长阶段并不相同[38]。在生长季,温度是影响马尾松茎干日变化量的主要因子。在非生长季,水分是控制马尾松茎干日变化量的主要因子(表2)。这可能是因为,在非生长季,温度降低,树木径向生长基本停止,此时,环境中水分的变化直接影响着树木茎干中水分的膨胀收缩。降水一方面会导致空气湿度的上升,使树木茎干吸水膨胀;另一方面通过增加土壤含水量来影响树木茎干的径向生长。
值得注意的是,温度在生长季不同阶段的影响亦并不相同。在生长季初期(3-5月),温度升高可以促进马尾松径向生长。但在生长季中后期(6-10月),过高的温度反而起抑制作用(表2)。3月月均气温为13.1 ℃,10 cm和20 cm土壤含水量分别为18%和15%。此时,空气温度适宜,土壤含水量相对较高。马尾松茎干径向生长开始启动,植物光合作用也开始加强,树干水分变化波动幅度大,马尾松月累积茎干径向变化量有了明显的增大,达到977.3 μm。随着径向生长速度的加快,累积径向变化量在6月达到第1个高峰。之后,过高的空气温度和较低的土壤含水量可能对马尾松径向生长产生抑制作用,使得径向生长在7-8月出现暂时性的生长变缓,累积径向变化量在7-8月下降到577 μm。当空气温度下降至20 ℃左右时,土壤含水量略有回升,马尾松再次进入快速生长期,在9月径向变化量达到第2个高峰。马尾松的“双峰”现象在闽北地区也有发现[31],其双峰值出现在5月和9月。但在湖北省秭归县地区的研究中[39],却并未发现马尾松双峰这一生长特征。由此可见,马尾松双峰生长特征可能具有区域性,后期有必要在大区域尺度开展马尾松生长监测,这将有助于加深触发对马尾松双峰生长所需要的水热特征及其相关环境阈值的认识和理解。
猜你喜欢
南京林业大学学报(自然科学版)(2022年1期)2022-11-29 10:10:16
中学生数理化·高一版(2021年3期)2021-06-09 06:10:20
数学物理学报(2021年1期)2021-03-29 03:14:18
重型机械(2020年3期)2020-08-24 08:31:40
农业机械学报(2020年3期)2020-04-01 08:11:10
绿色科技(2019年5期)2019-11-29 13:17:37
数学年刊A辑(中文版)(2019年3期)2019-10-08 07:34:44
种子(2019年5期)2019-07-02 11:37:44
天然产物研究与开发(2016年11期)2016-06-15 20:29:15
广西林业科学(2016年1期)2016-03-20 05:33:00
