
水热条件及坡向对长白山区蒙古栎林土壤呼吸的影响1)
2022-11-28 09:23孙忠林华明阳尹智博李岩冯颖姜云天
东北林业大学学报 2022年11期
孙忠林 华明阳 尹智博 李岩 冯颖 姜云天
(内蒙古民族大学,内蒙古·通辽,028043) (通化师范学院)
土壤呼吸作用在全球碳循环中具有重要地位,是除植被光合作用外,陆地生态系统第二大碳通量,也是植被光合固定的CO2返还大气的第一途径[1]。温带森林中近70%的生态系统呼吸来源于土壤[2],准确量化森林的土壤呼吸是陆地碳循环领域的重要内容,是准确评估特定生态系统碳源、碳汇功能的基础,是全球各个国家履行《联合国气候变化框架公约》与《京都协议书的》的依据[3]。土壤呼吸作用是生物地球化学循环模型中的重要组成部分,但模型的整合结果还存在很大的不确定性,定量评价水热因子的偶联和去偶联机制是准确评估土壤呼吸的重要前提[4-5]。对森林生态系统的长期研究,有助于深入理解和评估土壤呼吸对全球变化的影响[6]。土壤呼吸主要是土壤中生物代谢过程的结果,因此直接受土壤中生物因子(微生物、根系等)的影响,也间接受土壤中水热因子[7]、土壤因子(土壤养分、质地、pH等)和地表植被的影响[8-9],但各因子间存在很强的耦联关系。因此,分析水热因子的交互作用和互逆关系并确定主导因子,是准确评估土壤呼吸作用的关键;明确非生物因子对土壤呼吸的影响机制,有助于提高土壤呼吸模型空间尺度上推过程中的预测精度。
蒙古栎(QuercusmongolicaFisch. ex Ledeb.)在我国主要分布于东北地区和华北地区,是我国温带地区落叶阔叶林和针阔混交林的主要树种。蒙古栎林是我国东北林区分布面积最广的天然林型,随着全球气候变化,蒙古栎林地理分布范围极有可能继续扩大,成为东北林区的最主要林型[10]。蒙古栎林多分布于坡度较大、地形较为复杂的山坡和山顶,土壤保水能力相对较弱,季节性干旱是其分布区的典型特征。本研究以长白山区阴坡、阴坡蒙古栎天然次生林土壤为研究对象,以生长季(4—10月份)为研究时段,在蒙古栎林地阳坡和阴坡各设置3块样地(20 m×30 m),每个样地随机布置土壤聚氯乙烯(PVC)环9个(上、中、下每个坡位各3个),测定土壤呼吸速率、同步测量土壤环周围5 cm深度土壤温度和土壤体积含水量;以土壤温度、土壤体积含水量、呼吸速率、呼吸通量为评价指标,应用指数函数、线性函数,分析两个坡向蒙古栎林土壤呼吸速率的差异、水热因子对不同坡向蒙古栎林土壤呼吸的影响、生长季土壤呼吸通量的月动态变化。旨在为区域尺度上提高长白山区蒙古栎天然次生林土壤呼吸的预测精度提供参考。
1 研究区域概况
研究样地位于长白山脉的西南部,吉林省通化市境内(125°59′~126°E,41°44′~41°45′N)。该区域属中温带湿润气候区,年平均气温5.5 ℃,1月份平均气温最低(-14 ℃左右),极端最低气温达-33 ℃;7月份平均气温最高(22 ℃左右),极端最高气温36 ℃。通化地区年平均降水量870 mm,主要集中在6—8月份,降水量占年总降水量的60%以上,年日照时间2 200 h。该区域地带性土壤为暗棕壤,植被为伐后形成的天然次生林,主要包括蒙古栎林、落叶松(Larixgmelinii)林及红松(Pinuskoraiensis)人工林。
研究区域为蒙古栎天然次生林,林龄25 a(平均树龄,生长锥法测定)。蒙古栎的胸高断面积比例超过90%,伴生树种有紫椴(TiliaamurensisRupr.)和色木槭(AcermonoMaxim.);乔木密度为1 333株/hm2,胸高断面积22.30 m2/hm2,胸径(4.34±0.31)cm,树高(8.21±0.49)m。由于蒙古栎多分布于坡度较大、土壤水分渗透能力极强的生境下,所以成为该区域的优势树种。
2 研究方法
2.1 土壤呼吸速率的测定
分别在蒙古栎林阳坡、阴坡各设置3块样地(20 m×30 m),阴阳坡样地最小直线距离约50 m。每个样地随机布置土壤聚氯乙烯(PVC)环9个(上、中、下每个坡位各3个,环高5.0 cm、内径19.5 cm),把土壤环一端处理成楔形,以便压入土壤并减少对土壤的扰动。土壤环测量端露出地面3 cm,在整个生长季测定过程中土壤环的位置固定。应用LGR便携式温室气体分析仪(915-0011,Los Gatos Research Inc.,San Jose,USA)配备土壤呼吸气室(PS3000,Lica United,Beijing,China)测定土壤呼吸速率,使用该仪器自带温湿度探头就近同步测量土壤环周围5 cm深度土壤温度和体积含水量。测量时段为2017年4月12日—10月28日,每次测量时间间隔7 d,共观测29次。在试验样地外500 m左右,设置自动气象站(CR1000,CSI Campbell,USA),用于自动测定,空气和土壤温、湿度,太阳净辐射、光合有效辐射和降水等环境因子。
2.2 数据处理
对于单次观测结果而言,阳坡和阴坡土壤呼吸速率、土壤温度和体积含水量数据为同一天的测定结果(2个坡向观测间隔<3 h);假设以上3个指标在观测期间无明显的时间动态[11],通过配对数据T检验2个坡向温度、水分和土壤呼吸速率的差异,利用皮尔逊(Pearson)相关系数分析2个坡向土壤温度、体积含水量和土壤呼吸速率的变化趋势的一致性。
利用指数函数拟合时间尺度上土壤呼吸与土壤温度的关系,应用土壤呼吸温度敏感性(Q10)表示土壤呼吸的温度敏感系数,即温度升高10 ℃时土壤呼吸速率升高的倍数[12]。Rs=α×eβt;Q10=e10β;式中的Rs为土壤呼吸速率(单位为μmol·m-2·s-1),t为5 cm土壤温度(单位为℃),α为0 ℃土壤呼吸速率,β为温度反应系数。
蒙古栎林分布的土壤环境比较干旱、贫瘠,考虑到土壤温度与土壤湿度的交互作用可能对土壤呼吸速率产生一定的影响,本研究应用模型Rs=exp(a+b×t+c×W+d×W×t)拟合Rs与t、W之间的关系[13];式中的t为5 cm土壤温度(单位为℃),W为土壤5 cm处土壤体积含水量(用百分比计),a、b、c、d为方程系数。建模前把Rs对数转换,以满足线性和方差齐性;建模时选择向后剔除法,先建立全指标模型,若模型中与评价指标具有最小偏相关的影响因素符合设定标准(P>0.05),则最先从模型中剔出;然后,再根据设定的判定依据重复上述步骤,直到回归方程中不再含有不符合判据的影响因素为止。
由以上方差分析可知,在影响草果脱水量的因素中,微波作用时间和微波功率影响显著,物料量影响不显著。三因素及其交互作用对脱水量的影响见图4~图6。
应用指数函数分析土壤温度均值与月土壤呼吸通量之间的关系,应用线性函数分析月降水量与土壤呼吸均值之间的关系,应用协方差分析(ANCOVA)检验不同回归方程的显著差异(非线性关系转化为线性关系再比较)。
3 结果与分析
3.1 土壤温度、土壤体积含水量和土壤呼吸速率的时间动态
观测期间,蒙古栎林土壤温度在时间尺度上呈现单峰曲线形式(见图1a),阳坡的多数观测值大于阴坡,2个坡向土壤温度的变异(标准误)在生长季初和生长季末相对较大,生长季盛期相对较小,整体上温度变化趋势相似(R2=0.85,P<0.01)。阳坡温度均值18.37 ℃,阴坡温度均值17.66 ℃,2个坡向的温度峰值均出现于7月8日。阳坡土壤温度显著高于阴坡(n=29,t=1.85,P<0.05),但2个坡向温度的变异系数不存在显著差异,说明2个坡向样地尺度上土壤温度的差异较大。
土壤体积含水量在观测期间呈现不规则变动(见图1b),但绝大多数时间阳坡观测数据低于阴坡。阳坡土壤体积含水量均值14.17%,阴坡土壤土壤体积含水量均值18.92%,阴坡显著高于阳坡(n=29,t=6.87,P<0.01),且2个坡向相关显著(R2=0.60,P<0.01)。
在生长季内,2个坡向土壤呼吸速率均呈现双峰曲线形式(见图1c),变化趋势相似(R2=0.86,P<0.01),4—7月份为土壤呼吸速率的上升期,9—10月份为呼吸速率的下降期,峰值出现在7月份或8月份,但最低值都出现在生长季初期。阳坡和阴坡土壤呼吸速率平均值分别为1.90、2.20 μmol·m-2·s-1,阴坡比阳坡表现出更高的土壤呼吸强度(n=29,t=2.42,P=0.01),这种差异性表现出明显的季节特征,4—6月份2坡向土壤呼吸差异大于7—10月份。阳坡土壤呼吸的空间变异略高于阴坡,但没有显著差异。仅从土壤呼吸和土壤温、体积含水量的观测值在时间尺度上的分布看,2个坡向蒙古栎林土壤呼吸速率和土壤温度存在相关性。
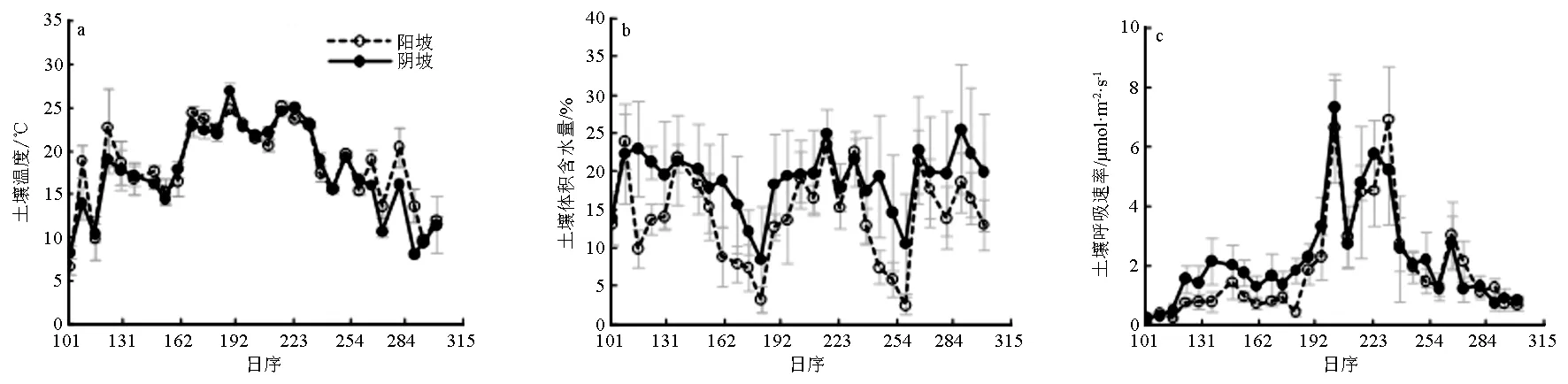
图1 土壤温度、土壤体积含水量和土壤呼吸速率的时间动态
3.2 土壤呼吸速率与土壤温度、土壤体积含水量的关系
由图2a可见:2个坡向土壤呼吸速率(Rs),随土壤温度(t)的增加均呈非线性增加趋势,呈显著的指数函数关系(n=29,P<0.01)。在阴坡时土壤温度对土壤呼吸速率影响的决定系数(R2)为0.580,阳坡时土壤温度对土壤呼吸速率影响的决定系数(R2)为0.295。从土壤呼吸程度看,阳坡和阴坡土壤呼吸速率的温度敏感性(Q10)分别为2.66、3.03,与阳坡相比,阴坡的土壤呼吸速率对温度的变化更为敏感,同等温度变化幅度条件下,阴坡土壤呼吸速率变化幅度更大。
2个坡向土壤呼吸速率与土壤体积含水量呈线性关系(见图2b),但不同坡向的土壤呼吸速率与土壤体积含水量相关性(R2)与增长趋势(方程斜率)均不同。阳坡方面,土壤呼吸速率与土壤体积含水量间的相关显著(P<0.05,R2=0.199),阴坡土壤呼吸速率与土壤体积含水量间相关不显著(R2=0.016)。从线性回归方程的斜率看,阳坡土壤呼吸速率对土壤体积含水量的变化更为敏感。
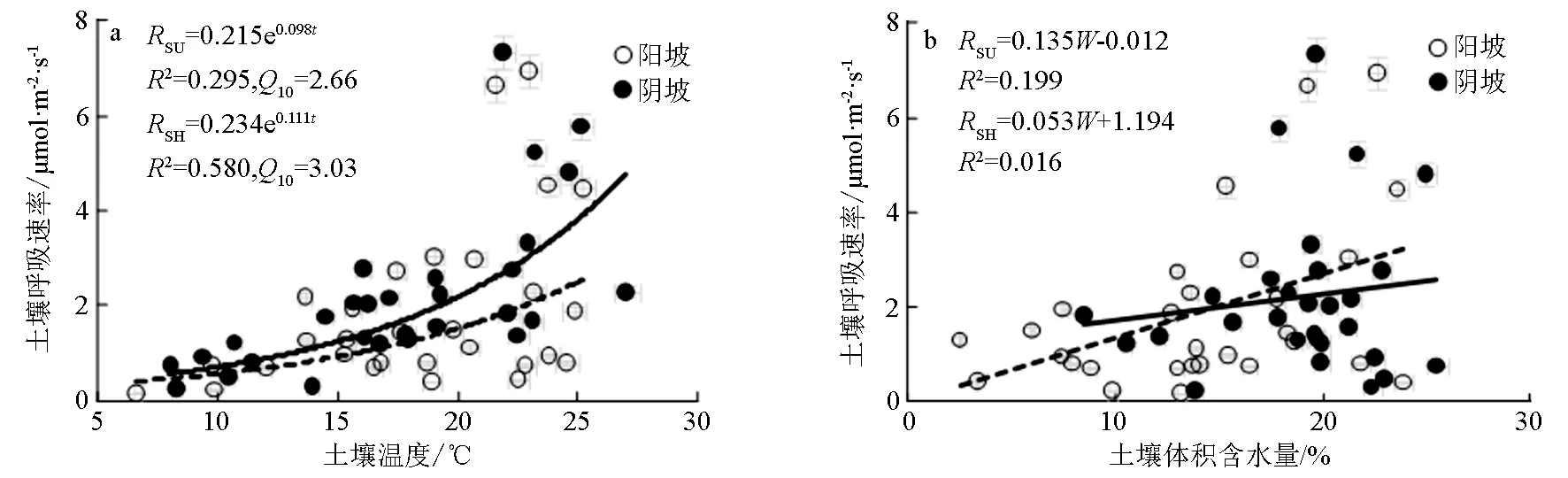
RSU为阳坡土壤呼吸速率;RSH为阴坡土壤呼吸速率;t为土壤温度;W为土壤体积含水量;R2为决定系数。图2 2个坡向土壤呼吸速率随土壤温度、土壤体积含水量增加的变化
多元回归结果表明,土壤呼吸速率与土壤温度、土壤体积含水量拟合的较好(n=29,P<0.01,见表1),但对于不同坡向蒙古栎林土壤呼吸速率的拟合结果中,回归方程的土壤温度值、土壤体积含水量和参数差别较大。从回归方程影响因素的拟合参数看,阳坡的土壤呼吸速率主要受土壤体积含水量(负相关)、土壤温度和土壤体积含水量的交互作用(正相关)影响显著,而阴坡的土壤呼吸速率主要受土壤温度(正相关)、土壤体积含水量(正相关)的影响显著。

表1 蒙古栎林2个坡向土壤呼吸速率与土壤温度、体积含水量的回归模型回归结果
3.3 土壤呼吸通量的月动态
月土壤呼吸通量(F)与月土壤平均温度(ta)呈指数关系(n=7,P<0.05,见图3a,土壤呼吸月通量与月土壤平均体积含水量关系不显著,但与月降水量(H)有较好的线性关系(n=7,P<0.01,见图3b)。土壤平均温度对土壤呼吸月通量影响显著(阳坡R2=0.614、阴坡R2=0.807),月降水量对土壤呼吸月通量影响显著(阳坡R2=0.866、阴坡R2=0.829)。曲线的拟合结果表明,随土壤温度和降水量的增加,2个坡向蒙古栎林土壤呼吸月通量呈现显著的增加趋势,但相同的月土壤平均温度或月降水量时,阳坡的月土壤呼吸通量低于阴坡。通过F-ta曲线推算,月土壤呼吸通量温度敏感性(Q10)分别为阳坡6.05、阴坡4.52,阳坡比阴坡高33.6%,说明同等条件时,阳坡土壤呼吸月通量对土壤平均温度变化的反应更加敏感。协方差分析结果表明,2个坡向土壤呼吸月通量随降水量增加的趋势相同(P=0.45),阴坡土壤呼吸的初始值显著高于阳坡(P<0.01);2个坡向土壤呼吸月通量随土壤温度增加的趋势和初始值,均存在显著差异(P<0.01),即转化后的线性方程斜率和截距均不同。
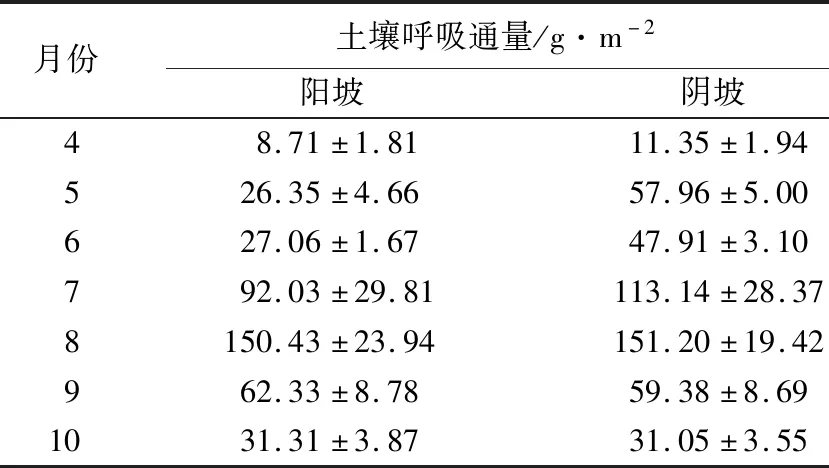
表2 2个坡向生长季内各月份土壤呼吸通量
4 讨论
4.1 土壤呼吸速率的月动态
土壤呼吸速率主要有土壤异养呼吸和根际呼吸两方面的贡献,不同的组分代表不同的生物过程[14],土壤呼吸组分受控因素和影响机制各异,因此土壤呼吸存在明显季节动态和年季差异[15-17]。温带森林的土壤呼吸研究结果表明,土壤呼吸速率大体上呈单峰曲线形式[18-20]。7、8月份土壤呼吸出现的峰值,这与水热联合驱动的土壤微生物的旺盛代谢活动和生长季细根生物量的季节动态贡献的根际呼吸量有很大关系[12]。本研究中,由于土壤呼吸的测定时间步长为1周,土壤呼吸在峰值期间出现的波动情况,这与降水在时间尺度上分布不均,导致土壤呼吸受水分胁迫有关。蒙古栎林通常分布于坡度大且干旱的生境,土壤水分对土壤呼吸速率产生了重要影响。
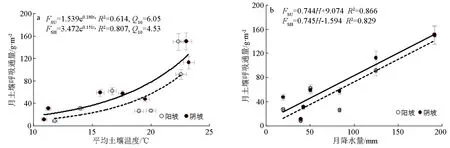
FSU为阳坡月土壤呼吸通量;FSH为阴坡月土壤呼吸通量;ta为土壤温度均值;H为月降水量;R2为决定系数。图3 2个坡向月土壤呼吸通量随着土壤温度均值、月降水量增加的变化
4.2 坡向对土壤呼吸速率主要影响因素的间接影响
本研究在日尺度上,2个坡向土壤呼吸温度敏感性(Q10)分别为2.66、3.03(见图2),位于温带森林土壤呼吸温度敏感性(Q10=1.8~4.1)[21]的中等水平,与大尺度研究结果的均值(3.0)[22]接近。阳坡土壤呼吸的温度敏感系数小于阴坡,说明干旱胁迫导致了土壤呼吸的温度敏感系数降低,土壤呼吸的敏感程度与水分的胁迫程度呈负相关[23]。
本研究的二元回归结果表明,2个坡向土壤呼吸速率均受水热因子的显著影响,说明水热条件共同对土壤呼吸起作用。阳坡土壤呼吸速率回归模型剔除了影响因素中的土壤温度,说明较为严重的干旱胁迫掩盖了土壤温度对土壤呼吸速率的影响;阴坡的土壤呼吸速率回归模型剔除了影响因素中的土壤温度和土壤体积含水量交互作用,说明干旱胁迫相对较轻,凸显了土壤温度对土壤呼吸的影响。由于水热条件的交互作用和互逆关系[24],土壤水分对土壤呼吸的影响容易被土壤温度效应所掩盖,因此要客观评价水热条件对土壤呼吸的影响,综合考虑土壤温度和土壤湿度2个影响因素是很有必要的。
4.3 坡向对土壤呼吸通量的影响
土壤呼吸速率受土壤温度和土壤湿度的综合影响,但影响程度和影响机制并不尽相同,这与底物有效性、植被特征、土壤理化性质、土壤微生物群落组成等多生态因子有关。本研究中,林分条件近乎相同的2个坡向蒙古栎林土壤呼吸速率存在显著差异,说明蒙古栎林土壤呼吸模型在空间尺度上推时,坡向是重要的影响因素。利用阳坡土壤呼吸数据推算的年通量将低估8.37%,利用阴坡数据则会出现同等水平的高估情况。阳坡由于受到严重干旱的胁迫,尽管土壤温度显著高于阴坡,其土壤呼吸通量却显著低于阴坡;阴坡受到土壤干旱胁迫相对较小,其土壤呼吸温度敏感性(Q10)更高。地形条件复杂的森林生态系统,土壤呼吸速率与通量的空间变异不但存在于林型之间[25],也存在于林型内部。
月土壤呼吸通量可以理解为更大时间尺度土壤呼吸速率。本研究中,2个坡向月土壤呼吸通量温度敏感性(Q10)高于观测尺度为7 d的土壤呼吸通量温度敏感性(见图2a,图3a),且7 d观测结果拟合得到的2个坡向土壤呼吸通量温度敏感性(Q10)为阳坡的小于阴坡的,而月土壤呼吸通量拟合的结果则为阳坡的大于阴坡的。说明土壤呼吸温度敏感性存在普遍的时空变异性[24,26],不同时间尺度拟合得到的土壤温度敏感性(Q10)反应不同地下生态过程对温度变化的敏感程度;季节尺度上,土壤水分、底物质量、微生物数量、微生物多样性、植物根系的物候变化等因素的影响,均是土壤温度敏感性(Q10)的变异来源;因此,不同时间尺度估测土壤呼吸通量,得出的结论也存在一定的差异。Boone et al.[12]研究表明,温带森林根际呼吸比异养呼吸的温度敏感性高将近1倍(84%),本研究中,土壤呼吸温度敏感性(Q10)的时间尺度上的变异,与根系的物候变化(根际呼吸的贡献)密不可分,具体机理有待进一步研究。
对于分布范围较广的森林生态系统,坡向是重要的地形因子。作为重要的环境因素,坡向通过改变特定生态系统的水热收支过程,影响生物化学进程。本研究中,土壤呼吸无论在7 d观测尺度,还是月尺度,2个坡向蒙古栎林土壤呼吸均存在显著差异。2个坡向土壤呼吸月通量均与降水量极显著线性相关(图3b),这在一定程度上体现了降雨激发土壤呼吸的“Birch效应”[27-28],说明干旱生境的降水是土壤呼吸预测的重要因素[29]。月降水量与月土壤呼吸通量较好的拟合,为大尺度森林生态系统土壤呼吸通量的推算提供了新的思路,但其适用的时空尺度和降水范围有待于进一步深入的研究。
5 结论
坡向对蒙古栎林土壤呼吸速率有显著影响,蒙古栎林阳坡和阴坡的土壤呼吸通量分别为398.22、470.99 g·m-2,阴坡比阳坡高18.27%。
土壤体积含水量、土壤温度、土壤体积含水量与土壤温度的耦合作用,均对土壤呼吸过程有不同影响。
坡向是复杂地形蒙古栎林生态系统重要的环境因素。坡向通过影响水热因子的分配,影响土壤的物理性质,进而影响土壤中的生物化学过程,在利用微气象学方法推算区域尺度的土壤呼吸通量时,坡向是主要指标之一。
不同时间尺度计算的土壤呼吸温度敏感性,反应不同的生物学过程,因此在模型整合时应严格区分观测尺度。利用降水量推算土壤呼吸的月排放通量,能在一定程度上提高蒙古栎林碳循环模型的预测精度。
猜你喜欢
种子科技(2021年17期)2021-10-29
思维与智慧·下半月(2021年7期)2021-08-13
现代农村科技(2018年11期)2018-11-15
安徽农学通报(2017年11期)2017-06-23
课程教育研究(2017年11期)2017-04-17
绿色科技(2017年1期)2017-03-01
湖北农业科学(2014年16期)2014-10-20
现代农业科技(2009年1期)2009-04-09
