
高原鼠兔对高寒草甸植物物种多样性和功能多样性关系的影响
2022-11-23 07:38段媛媛张静王玲玲刘彩凤王乙茉周俗郭正刚
草业学报 2022年11期
段媛媛,张静,王玲玲,刘彩凤,王乙茉,周俗,郭正刚*
(1.兰州大学草地农业生态系统国家重点实验室,兰州大学农业农村部草牧业创新重点实验室,兰州大学草地农业教育部工程研究中心,兰州大学草地农业科技学院,甘肃 兰州 730020;2.四川省草原科学研究院,四川 成都 611731)
植物物种多样性主要表现草地资源的丰富性[1],而植物功能多样性则主要强调草地植物群落内不同植物物种间功能性状的总体差异,以及植物性状对环境变化的响应[2-3]。无论是植物物种多样性,还是植物功能多样性,两者均对草地生态系统功能的变化具有决定作用[3]。植物物种多样性和植物功能多样性关系,不仅是揭示植物群落内物种共存的重要途径之一[4],而且是草地生态系统稳定的基础[5],更是制定区域内生物多样性保护策略和草地资源合理利用的关键[6]。当草地环境改变时势必引起植物物种多样性和功能多样性的联动变化,进而影响植物物种多样性和功能多样性间的关系[3-4,7-8],以及生态系统的稳定性和物种共存的机制。草地环境的改变,要么是生物因子变化所致[1,9],要么是非生物因子变化所致[3-4]。已有研究表明,当草地刈割强度增加时,植物物种多样性和功能多样性的关系从正线性相关变为不相关,氮肥添加量却不会影响两者的正线性关系[7]。当水域生态系统的盐分浓度从低增高时,植物物种多样性和功能多样性间因相关系数变小而导致关联度降低[10],这说明非生物和生物干扰确实能够明显改变植物物种多样性和功能多样性间的关系。小型啮齿类动物是草地的主要组分,也是草地生态系统的重要干扰因子[11-12]。小型啮齿类动物往往镶嵌分布于天然草地,其活动会改变高寒草甸的水热过程[9,11]和养分异质性[9,13],但其是否影响植物物种多样性和功能多样性的关系,尚需要科学的试验提供证据,从而有助于解释小型啮齿类动物干扰影响草地生态系统的稳定性和物种共存机制。
高原鼠兔(Ochotona curzoniae)是青藏高原特有的小型啮齿类动物,主要分布于青藏高原高寒草甸,不仅是高寒草甸的生态工程师[11],而且是主要生物干扰因子之一[13-17]。高原鼠兔通过刈割、掘洞、覆土、排泄等[14],会通过增加生境的异质性而增加植物物种多样性[13,18-19]、杂草类功能群丰富度指数[20],而且会降低植物均匀度指数[15]、禾草科和莎草科功能群丰富度指数[20]。高原鼠兔分布区内植物覆盖度和土壤养分的变化,必然会改变植物利用资源的空间维度,从而影响植物功能多样性[9,21-24]。不同植物从生存环境中获取利用资源的方式不同[6],物种间资源利用的互补性会将植物物种多样性和功能多样性有机耦联[25],这说明高原鼠兔干扰可能会影响高寒草甸植物物种多样性和功能多样性的关系。
植物物种多样性通常采用物种丰富度指数、物种多样性指数、物种均匀度指数测度[20]。虽然测度植物功能多样性的指数较多,但常用指数主要是功能丰富度指数、功能均匀度指数和功能离散度指数[25]。目前已有研究分析植物物种多样性指数和功能多样性指数间的关系时,均采用一个植物物种多样性指数对应多个植物功能多样性指数,而不是一一对应关系[3,8,10]。因此本研究在青藏高原的甘肃碌曲、青海祁连和共和县3个地点同步开展野外调查,采用3个地点的数据,首先分析高原鼠兔干扰下植物物种多样性和功能多样性常用指标的变化特征,然后阐明高原鼠兔干扰对植物物种多样性和功能多样性关系的影响,以期为理解高原鼠兔干扰下高寒草甸植物物种多样性和功能多样性的维持机制提供基础信息,为全面解读高原鼠兔在高寒草甸生态系统中的作用提供独特视角。
1 材料与方法
1.1 研究区概况
高原鼠兔分布广,栖息地气候、地形、海拔差异大[11,13]。若采用单一地点作为研究对象,其结果可能是高原鼠兔干扰所致,也可能是高原鼠兔干扰与环境因子叠加所致。因此,本研究选择碌曲县尕海镇(102°22′12″E,34°15′51″N)、祁连县默勒镇(100°34′48″E,37°43′26″N)和共和县黑马河乡(99°47′11″E,36°43′48″N)做研究地点,同步分析高原鼠兔干扰对高寒草甸植物物种多样性和功能多样性关系的影响,以期发现高原鼠兔干扰对高寒草甸植物物种多样性和功能多样性关系影响的普遍特征。3个研究地点的气候均为高原大陆性气候,具有高寒阴湿特征,土壤类型为高山草甸土[26],其中尕海镇、默勒镇和黑马河乡的海拔分别为3550、3468、3194 m,年均温分别为4.9、2.1、5.6℃,年均降水量分别为653、420、400 mm。虽然3个地点的高寒草甸均以莎草科植物为优势种,但不同地点高寒草甸的优势种存在分异,其中尕海镇高寒草甸优势种为线叶嵩草(Kobresia capillifolia),主要伴生种为草地早熟禾(Poa pratensis)、垂穗披碱草(Elymus nutans)、莓叶委陵菜(Potentilla fragarioides);默勒镇高寒草甸的优势种为高山嵩草(Kobresia pygmaea),主要伴生种为矮生嵩草(Kobresia humilis)、美丽风毛菊(Saussurea pulchra)、莓叶委陵菜;黑马河乡高寒草甸的优势种为高山嵩草,主要伴生种为矮生嵩草、垂穗披碱草、草地早熟禾、莓叶委陵菜、二裂委陵菜(Potentilla bifurca)、乳白香青(Anaphalis lactea)。3个地点的高寒草甸均被牧户承包,每家牧户承包的高寒草甸被分为冷季牧场和暖季牧场,有些暖季牧场为多个家庭共用,没有明显的围栏边界,冷季牧场为每户单独所有,具有明显的围栏边界。
1.2 研究方法
1.2.1 植物功能性状指标的选取与测定 植物功能多样性指数往往通过植物功能性状指标计算所得。测度植物功能性状的指标很多,但不同研究所选择的指标差异很大[3,8,10],然而任何研究选择植物功能性状指标时,均需要考虑植物形态特征、生殖特征和生理特征[3]。本研究选择植物功能性状指标时,既考虑植物的形态特征、生殖特征和生理特征,又考虑了选择指标数据获取的可操作性和易测性。因此,选择植物高度、盖度和密度作为植物形态特征的测度指标,传粉方式、繁殖类型和生活史作为植物生殖特征的测度指标,固氮能力作为植物生理特征的测度指标,共计7个功能性状指标。采用样方法收集植物高度、盖度及密度的数据信息,采用野外形态观察和文献相结合的方法获取植物传粉方式、生活史、固氮类型、繁殖类型的信息[27]。采用赋值方式量化植物传粉方式、生活史、固氮类型、繁殖类型的功能特征,其中植物传粉方式为风媒传粉时赋值为0、虫媒传粉时则赋值为1、风媒传粉和虫媒传粉两者兼有时赋值为0.5;生活史为一年生植物时赋值为0、多年生植物时赋值为1、一年生或多年生植物兼有时赋值为0.5;固氮方式为自身固氮时赋值为0、非自身固氮时赋值为1;繁殖方式为营养繁殖时赋值为0、有性繁殖时赋值为1、营养繁殖和有性繁殖兼备则赋值为0.5。
1.2.2 试验设计 在每个研究地点,选择地势相对平坦的冷季放牧地作为调查取样区。调查区内植物生长季不放牧,可排除植物生长季的放牧影响,以期最大限度反映高原鼠兔干扰对高寒草甸植物物种多样性和功能多样性关系的影响。高原鼠兔是地面群居性小型啮齿类动物[16],喜欢栖息于低矮开阔的生境[9,18],因此高原鼠兔栖息地呈片状分布状态。虽然有些高寒草甸处于低矮开阔生境,但目前没有被高原鼠兔占据,是高原鼠兔潜在的分布区,是因为高原鼠兔扩散是一个逐步的过程[28],因此可以将高原鼠兔潜在分布区作为对照区[29],实现分析高原鼠兔干扰对植物物种多样性和功能多样性关系影响的目标。
采用随机分层配对设计的方法,确定每个地点调查取样区的样地。高原鼠兔巢域的平均面积为1262.5 m2[12],因此样地面积设置为35 m×35 m,最大可能地接近巢域平均面积。针对一个研究地点,首先根据高原鼠兔出现和有效洞穴存在与否,确定第一个干扰样地,然后寻找第二个干扰样地,以此类推,每个地点选择10个干扰样地,干扰样地之间的距离大于5 km,避免因高原鼠兔移动而带来的影响。其次,针对每个干扰样地,设置一个未干扰样地,设置未干扰样地的基准为其地貌、植物群落和土壤类型与干扰样地基本一致。未干扰样地与干扰样地之间的距离为0.5~1.0 km,若距离过大,则很难保证未干扰样地与干扰样地的地貌、植物群落和土壤类型基本一致,距离过小,则高原鼠兔的活动会影响到未干扰样地的植物群落和土壤。每个研究地点选择10组配对样地,共计60个试验样地,包括30个干扰样地和30个未干扰样地。
1.2.3 野外调查与取样 高原鼠兔一年内繁殖具有多崽多胎特征[12],每年8月种群密度达到峰值[14,29],对高寒草甸的干扰程度最明显。因此野外调查与取样于2020年8月进行。每个样地内(干扰样地与非干扰样地)布设5个1 m×1 m(1 m2)的植物群落调查样方,样方设置为“W”型布局,样方之间的距离为8 m。针对干扰样地,若样方恰好落在裸斑时,则需要小幅度移动样方位置,避开裸斑。共计调查样方300个,其中每个研究地点100个,其中干扰样地和未干扰样地分别为50个。调查时,首先记录每个样方中出现的植物种类,然后测定每种植物的高度、盖度和密度,其中植物高度为随机5株的平均高度,若植物出现次数小于5次,则采用所有出现次数的平均值作为该植物的高度[10];每种植物盖度通过“针刺法”获得[1,20];植物密度测定分双子叶植物和单子叶植物,双子叶植物以株为基准,而单子叶植物以样方内的一级分枝数为基准[7]。实验室内根据文献,判别每种植物的传粉方式、生活史、固氮类型和繁殖类型[27]。
1.2.4 植物物种多样性和功能多样性指数计算方法 植物物种多样性采用物种丰富度指数Patrick(S)、物种多样性指数Shannon(H)、物种均匀度指数Pielou(J)测度。利用物种高度、盖度、密度计算每个物种的重要值(important value,IV),公式为:

式中:IV代表物种重要值,A代表植物相对高度,为植物5个高度的平均值与样方内所有植物平均高度总和的比值,B代表植物相对盖度,为植物盖度与样方内所有植物盖度总和的比值,C代表植物相对密度,为植物密度与样方内所有植物密度总和的比值。
物种丰富度指数Patrick(S):

物种多样性指数Shannon(H):

式中:S是样方内的物种数,Pi是种i的相对重要值。
物种均匀度指数Pielou(J):

式中:S是样方内的物种数,H是物种多样性指数。
植物功能多样性指数采用功能丰富度指数(functional richness index,FAD2)、功能均匀度指数(functional evenness index,FEve)和功能离散度指数(functional divergence index,FDis)测度[21],计算过程包括4个步骤:获得功能性状矩阵、将性状矩阵转化为距离矩阵、将距离矩阵聚类生成树状分类图、计算树状分类图的分枝总长度。
功能丰富度指数的计算公式:

式中:S是样方内出现的物种总数,dxy表示物种x和y之间的欧几里得距离,mtx和mty分别表示物种x和y之间第t个功能性状的观测值,t=1、2、3……T,T为性状数量。
功能均匀度指数:


式中:PEWl表示局部加权平均均匀度,EWl表示加权平均的均匀度,Px和Py分别表示物种x和y在群落中的相对密度。
功能离散度指数:

式中:c表示加权重心,Py表示物种y在群落中的相对密度,Zy表示物种y与重心c的加权距离,mxk表示物种x的第k个性状值。
1.3 数据分析
首先用夏皮罗-威尔克检验(Shapiro-Wilk test)检测3个地点整体数据和各个地点数据的方差齐性,若方差不齐性则采用Log或者平方转化,使其满足正态分布。其次采用Two-Way ANOVA分析方法,将地点和高原鼠兔干扰作为固定因子,3个植物物种多样性指数和3个植物功能多样性指数作为因变量,分析地点、高原鼠兔干扰及两者互作是否显著影响测度植物物种多样性的3个指数和植物功能多样性的3个指数;若高原鼠兔干扰对某个指数有显著影响,则用One-Way ANOVA方法分析每个地点高原鼠兔干扰对该指数的影响,以验证高原鼠兔对高寒草甸植物物种多样性和功能多样性的影响是否具有普遍特征;若高原鼠兔干扰对某个指数影响不显著,则不分析每个地点高原鼠兔干扰对该指数的影响。最后用Person相关分析法在0.05和0.01水平上评价植物物种多样性的3个指数与植物功能多样性的3个指数间是否存在显著关联性,若两个指数间关系显著,则采用线性、对数、多项式、指数等函数拟合两者关系,根据决定系数R2最大原则,遴选两个指数关系的最优方程;若两个指数间关系不显著,则不做遴选。上述数据分析和方程拟合均采用SPSS 24.0统计分析,采用Origin 2019制图。
2 结果与分析
2.1 高原鼠兔对高寒草甸植物物种多样性的影响
无论是3个地点数据整体分析,还是单个地点数据单独分析,高原鼠兔干扰均显著增加了植物物种丰富度指数和植物物种多样性指数(P<0.01),但没有显著影响植物物种均匀度指数(图1)。

图1 高原鼠兔对植物物种丰富度指数、植物物种多样性指数和植物物种均匀度指数的影响Fig.1 Effect of plateau pika on plant species richness index,plant species diversity index and plant species evenness index
2.2 高原鼠兔对高寒草甸植物功能多样性的影响
3个地点数据整体分析结果表明,高原鼠兔干扰显著增加了植物功能丰富度指数(P<0.01),但没有显著影响植物功能均匀度指数和植物功能离散度指数(图2)。每个地点数据单独分析时,高原鼠兔干扰对植物功能丰富度指数、植物功能均匀度指数和植物功能离散度指数的影响与3个地点统一分析时一致。
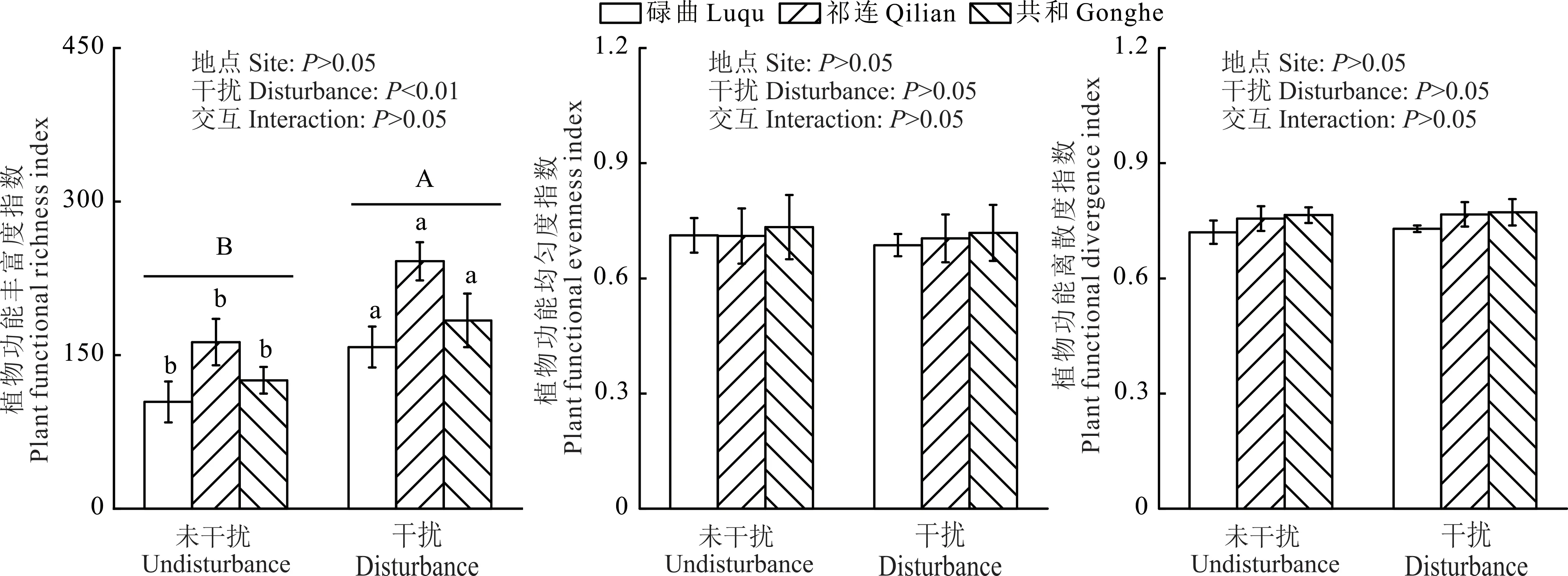
图2 高原鼠兔对植物功能丰富度指数、植物功能均匀度指数和植物功能离散度指数的影响Fig.2 Effect of plateau pika on plant functional richness index,plant functional evenness index and plant functional divergence index
2.3 高原鼠兔对高寒草甸植物物种多样性和功能多样性关系的影响
高原鼠兔干扰对高寒草甸植物物种多样性和功能多样性关系的影响随着指标变化而出现分异(表1)。植物功能丰富度指数与植物物种丰富度指数间关系在未干扰生境内最优拟合方程为对数函数y=0.7449ln(x)+0.1245,R2为0.9378,而在高原鼠兔干扰生境内则变为二次函数y=-0.0001x2+0.0346x+1.6999,R2为0.8800,虽然两者关系在未干扰和干扰生境内均呈显著正相关(P<0.01),但两者的相关系数从0.959变为0.938,说明关联性具有减弱趋势。虽然植物功能丰富度指数与植物物种多样性指数间的关系在未干扰和干扰生境下均为正相关二次函数,但相关系数从0.926变为0.913,说明两者间的正相关关系呈现减弱趋势(P<0.01),而植物功能离散度指数与植物物种多样性指数关系在未干扰和干扰生境内最优拟合方程均为二次函数,相关系数从-0.458变为-0.502,说明高原鼠兔干扰增强了两者的负相关关系(P<0.01)。植物功能离散度指数与植物物种丰富度指数的关系在高原鼠兔干扰和未干扰生境内最优拟合方程均为线性函数,相关系数由-0.580变为-0.520。高原鼠兔干扰让植物功能均匀度指数与植物物种均匀度指数关系从未干扰生境内的不显著相关,变为干扰生境内的显著正相关(P<0.01)。高原鼠兔干扰没有改变植物功能丰富度指数与植物物种均匀度指数、植物功能均匀度指数与植物物种丰富度和植物物种多样性指数,以及植物功能离散度指数与植物物种均匀度指数间的关系。3个研究地点单独分析时,其结果基本趋同于整体分析结果。

表1 高原鼠兔对植物物种多样性与功能多样性关系的影响Table 1 The relationship between plant species diversity and functional diversity with plateau pika
3 讨论
草地植物物种多样性和功能多样性间的关系变化,取决于植物物种多样性、植物功能多样性,或者两者的同时改变[4-6,10]。本研究表明,高原鼠兔干扰通过同时改变植物物种丰富度指数和植物功能丰富度指数,而减弱了两者间的正相关关系,说明高原鼠兔干扰下植物物种丰富度指数表征植物功能丰富度指数的程度有所降低,这主要与高原鼠兔干扰产生大量星罗棋布的裸斑有关。高原鼠兔干扰增加植物物种多样性的途径主要包括:首先,高
原鼠兔干扰产生大量裸斑,而裸斑的存在会增加降水下渗率,减少地表径流的活动,这会增加高寒草甸的生境异质性[4,16],从而为多种植物共存提供了更好的环境[4,13];其次,高原鼠兔栖息地因裸斑土壤内速效磷、氨态氮和硝态氮较高[30],这有助于更多嗜氮的禾草植物定居[18];第三,高寒草甸因地表存在致密的草毡层,一定程度上会隔离植物种子和土壤的接触,迫使部分植物种子虽然能够萌发,但不能定居,而裸斑则为植物种子和土壤的充分接触创造了更好的条件和机会[29],有助于萌发种子成功定居[13]。高原鼠兔干扰不仅会增加新物种[13,18],而且会引起肥岛效应[31-32],这必然会改变高寒草甸原有植物群落内植物物种间的关系,促进植物各自适应新的土壤生长环境和竞争关系[33],从而引起植物群落中植物利用和获取资源的途径更加多样化[1,7],导致植物个体功能呈现多样化趋势,占据更多生态空间,增加植物群落的功能性状特征和植物群落功能丰富度。虽然高原鼠兔存在会增加高寒草地生态系统内整体接纳降水的能力,强化水源涵养功能[10,17],但高原鼠兔栖息区内大量裸斑会加速土壤水分从表层向深层渗透[17],同时裸斑因无植物覆盖,地表水分蒸发量增加,从而导致高原鼠兔栖息区内土壤表层含水量降低[20,34]。而植物根系主要生长在土壤表层,因此,针对植物生长而言高原鼠兔栖息区的生境具有旱化特性[20],这势必诱导植物群落组分发生一定的改变,湿生植物具有逐渐减少乃至退出的趋势,而中生和旱生植物则具有逐渐增多的趋势[13,18],植物群落内组分的变化和环境压力增加时植物强烈的趋同选择会限制植物功能多样性的变化[4,7-8],客观上形成了高原鼠兔干扰会减弱植物物种丰富度和植物功能丰富度间关联程度的现象。
高原鼠兔干扰促进植物功能丰富度指数与植物物种丰富度指数的关系从对数函数变为二次函数,说明植物功能丰富度随物种丰富度增加的变化过程发生了明显的变化。高寒草甸没有高原鼠兔栖息的生境内,植物群落内物种丰富度增加时,植物功能丰富度的增长率一直处于减小状态[8],说明植物物种丰富度增加到一定程度时,植物功能性状会达到饱和状态,此时植物群落功能冗余达到最高[7],此后植物功能丰富度随植物物种丰富度增加几乎不会发生较大变化[8],此过程趋同于种群增长的S型曲线。高寒草甸栖息高原鼠兔后,植物功能丰富度指数与植物物种丰富度指数的关系呈现为开口向下的二次函数,说明植物功能丰富度随植物物种丰富度增加表现为先增加后降低的过程,这是因为随植物物种丰富度增加,植物群落功能性状存在一个阈值[8],在阈值内适应性强的植物由于较高的生存率和建立率存活下来并且保留功能性状[20,35],而随着植物物种丰富度继续增加,部分适应性弱的植物会慢慢退出或消失[13]。虽然高原鼠兔干扰生境内会出现新物种[18,29],但新增加物种的植物功能性状不足以弥补消失植物的功能性状[20],植物功能丰富度则反而降低,两者关联强度减弱。
植物功能均匀度指数与植物物种均匀度指数的关系从高原鼠兔未干扰生境下的不显著相关,转变为高原鼠兔干扰生境下的显著正相关,说明高原鼠兔干扰增强了植物功能均匀度和植物物种均匀度间的关联性。高原鼠兔没有栖息的高寒草甸,其植物群落内各个物种间的关系已经通过种内和种间竞争,形成了以莎草科植物为优势的稳定结构[14,20],群落内植物功能特征差异相对较大[29],此时植物群落功能冗余的保险效应[14,20],导致植物功能均匀度指数与植物物种均匀度指数间关联性不高[36];而高原鼠兔栖息高寒草甸后,则会因创建开阔视野而刈割高大植物[9,11],弱化高大植物的竞争优势[14,18],从而增加了高寒草甸整体的光捕获叶面积和光合速率[9,29],环境中光和碳的利用率增加[24];同时高原鼠兔的选择性采食会抑制被食植物生长,释放不喜食植物生长潜势[18,33],从而改变了原有植物群落内不同植物物种利用资源的格局[8,36],客观上减小了植物群落内不同植物物种间功能特征的差异[21,23];高原鼠兔与雪雀(Montifringilla ruficollis)相伴而生[13,18],高原鼠兔粪便和雪雀会更加均匀地散布更多的植物种子[14,29],促进植物功能特征在生态空间上更加均匀分布[13];此外,高原鼠兔裸斑斑块的存在,会增加土壤构型的异质性,反哺植物均匀度增加[36],上述3个过程导致了植物功能均匀度指数和植物物种均匀度指数的关系变为显著正相关。
草地群落植物物种丰富度的状态决定着种间功能性状的差异,当物种丰富度低时,功能性状相对差异较大[8],因此高原鼠兔干扰丰富了植物群落内的物种丰富度,从而减弱了高原鼠兔干扰区内植物功能离散度和植物物种丰富度间的负相关关系。没有高原鼠兔的高寒草甸内,植物物种丰富度较低(图1),植物群落内具有简单的种间和种内互作模式[35],每个植物物种具有较大的生态幅,物种功能性状多样化程度较高,而高原鼠兔入侵后,其选择性采食会促进部分物种趋同选择,从而限制植物群落功能多样性的变化[4,7,17],而增加的物种数会导致类似植物功能群增加[5],迫使植物群落内个体功能性状趋向性状空间重心[10,25],此时植物功能离散度与植物物种丰富度间的关系会减弱。
4 结论
高原鼠兔干扰对植物物种多样性和功能多样性的影响,随着测度指标的不同而出现分异,说明评价植物物种多样性和功能多样性时,选择合适的指标至关重要。虽然本研究结果证实,高原鼠兔干扰能够明显改变植物物种多样性和功能多样性间的关系,但随测度植物物种多样性和功能多样性指标的不同而呈现不同响应结果,譬如植物功能丰富度指数和物种丰富度指数间关系从对数函数变为二次函数,但植物功能均匀度指数和物种均匀度指数间的拟合函数类型不会发生改变,但增加了它们间的关联性,显著减弱了植物功能丰富度指数和物种多样性指数间的正相关性,以及植物功能离散度指数与物种丰富度指数间的负相关性,增强了植物功能离散度指数与物种多样性指数间的显著负相关性。而植物功能丰富度指数和植物物种均匀度指数、植物功能均匀度指数和物种丰富度指数、植物功能均匀度指数和物种多样性指数,以及植物功能离散度指数和物种均匀度指数间的关系没有发生明显的改变。本研究结果虽然初步证实了高原鼠兔干扰能够影响高寒草甸内植物物种多样性和功能多样性间的关系,但今后需要测定更多的植物功能多样性指标,遴选哪些测度植物功能多样性的指标能够较高的反映两者间的关系。本研究为啮齿动物干扰的高寒草甸植物群落物种共存机制提供了信息。
猜你喜欢
林业资源管理(2022年6期)2023-01-18
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
现代园艺(2021年23期)2021-12-01
海外文摘·文学版(2021年3期)2021-05-19
新农业(2020年18期)2021-01-07
花卉(2020年24期)2021-01-04
河南畜牧兽医(2020年21期)2020-01-10
活力(2019年21期)2019-04-01
北京航空航天大学学报(2017年3期)2017-11-23
