
科学史视角下“基因在染色体上”的探索过程∗
2022-11-22 08:12王玉龙广东实验中学广东广州510055
生物学通报 2022年1期
王玉龙 (广东实验中学 广东广州 510055)
在学习了摩尔根(Morgan)红眼、白眼果蝇杂交实验后,学生对基因在染色体上的结论将信将疑。教师进一步解释布里吉斯(Bridges)的果蝇杂交实验中出现的初级例外情况后,学生逐渐接受了基因在染色体上的观点,但很难想到染色体变异的方向,对布里吉斯本人为何能提出相应的假说解释依然充满疑惑。对上述疑惑的解答需要从基因在染色体上(染色体遗传)的探索过程讲起。
1 基因与染色体的关系假说
1883—1892年,魏斯曼(Weismann)所提出的“种质论”认为,生物可分为“种质”和“体质”2 个部分。体质由种质发育而来,在世代之间不连续,种质在世代之间是连续的。虽然该理论之后被证明是错误的,但有力驳斥了“用进废退”和“获得性状遗传”的观点。魏斯曼结合受精现象的观察认为,遗传的物质基础存在于细胞核内部,并预测了遗传物质在细胞核内某种载体之上,为染色体-DNA(核酸)-基因关系的确立奠定了基础。
1900年,3位科学家几乎同时重新发现了孟德尔(Mendel)的划时代论文《植物杂交的试验》,总结了孟德尔遗传理论的精华,可概括为“颗粒遗传学说”。该学说后面逐渐取代了达尔文(Darwin)坚持的“泛生论”和魏斯曼提出的“种质论”的遗传学说,1909年约翰逊(Johannsen)将“颗粒遗传学说”中的“颗粒”命名为基因[1]。
基因究竟在哪里?鲍维里(Boveri)以海胆为材料,取出卵细胞的细胞核,植入精子,进一步处理得到2 个精子受精的“受精卵”,通过研究其发育情况,证明了细胞核在遗传中的决定作用,同时发现不同的染色体有不同的性质。鲍维里还根据其对蛔虫的研究,发现染色体具有遗传的连续性和独立性,认为受精卵分裂形成的细胞,其中的染色体一半来自父方、一半来自母方。1903年,萨顿(Sutton)通过研究蝗虫生殖细胞的形成过程,发现了基因和染色体存在明显的平行关系,进而推测基因在染色体上。1905年,威尔逊(Wilson)认为生物体的性状多于染色体数目,若基因和染色体有关,则一条染色体上含有多个基因,即会存在基因连锁。
在摩尔根研究果蝇之前,关于基因和染色体的关系假说已存在,只是缺少相应的实验证据予以证明。但摩尔根本人的学术研究经历使其并不认同孟德尔遗传学说,他曾用家鼠与野鼠做杂交实验,结果发现毛色性状的遗传毫无规律,使其对孟德尔学说产生了怀疑;当陆续有更多的动物遗传实验难以用孟德尔学说解释时,也就对此学说有了根深蒂固的成见[2]。
2 性染色体的发现
1891年,亨金(Henking)发现昆虫精巢细胞中含有一个不同的染色体,并将其命名为X 染色体。1901年,麦克隆(McClung)提出X 染色体与性别有关,但错误地认为X 染色体决定雄性[3]。1905年史蒂文斯(Stevens)首次提出甲壳虫的Y 染色体与雄性有关,雄性为XY,雌性为XX。但当其研究果蝇装片时,由于装片制作效果不理想,并未发现Y染色体,认为果蝇的雌性为XX,雄性只有一条X染色体[3]。性染色体及性别决定的发现,为后续伴性遗传的研究奠定了基础。
3 摩尔根果蝇杂交实验
1908年,摩尔根建议其学生佩恩(Payne)验证拉马克(Lamarck)“用进废退”学说的正确性,佩恩以果蝇为材料,在暗室中饲养,观察其是否会因为不需要眼睛而产生无眼后代。结果发现一直饲养至69 代时,均未出现无眼果蝇,进而证明了“用进废退”学说的不合理性。摩尔根也由此发现了果蝇是理想的遗传学实验材料,便创建了“蝇室”,饲养果蝇进行遗传学实验。1910年5月,摩尔根偶然在蝇室内野生型果蝇中,发现了一只白眼雄果蝇,他马上意识到该果蝇的重要性,在其死亡之前完成了与红眼雌果蝇的杂交。所得F1代中有1 237 只红眼果蝇和3 只白眼果蝇,摩尔根认为这3 个例外情况是进一步发生突变导致的,未对其进行讨论。继续将F1的红眼果蝇相互交配,所得F2代中有2 459 只红眼雌果蝇,1 011 只红眼雄果蝇,782 只白眼雄果蝇。摩尔根在上述实验的基础上,继续进行回交实验,让F1的红眼雌果蝇和最初出现的白眼雄果蝇杂交,结果出现红眼雌性、红眼雄性、白眼雌性和白眼雄性4 种表型,比例接近1∶1∶1∶1[5]。
通过分析数据可知,红眼果蝇与白眼果蝇的比例并非3∶1,但摩尔根认为考虑到白眼果蝇存活率较低的原因,可认为该比值为3∶1,但白眼果蝇仅限于雄性,这一点与孟德尔实验并不相同。摩尔根虽然对孟德尔学说抱有很大成见,但在事实面前,摩尔根还是以颗粒遗传学说作为解释。由于当时并未发现果蝇中有Y 染色体,加之摩尔根对基因在染色体上的学说也有很大疑虑,所以其解释过程并非如高中《生物》教材中所介绍的那般,更不可能提出如同很多高中生物学教师所讲的“白眼基因位于X 染色体的非同源区段,位于X、Y 同源区段,位于Y 的非同源区段”等假说。而是使用了如下解释过程,如图1所示。
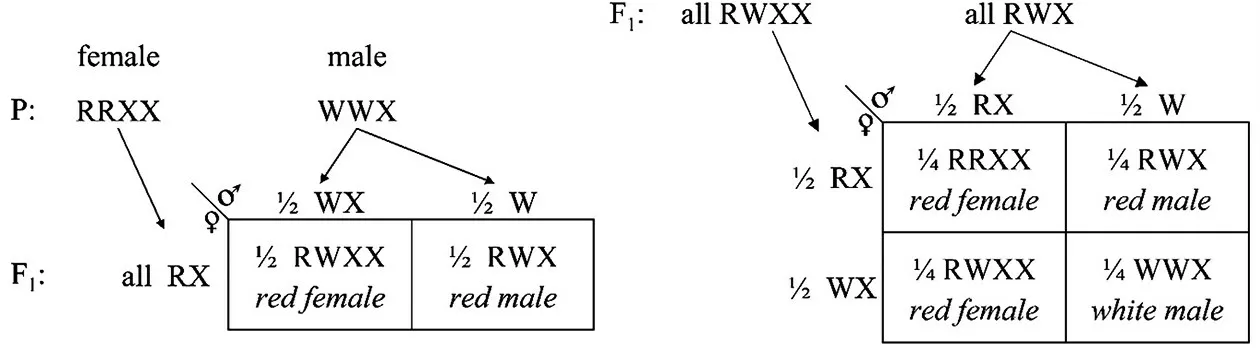
图1 1910年摩尔根论文中的遗传图解[5]
在上述解释中,R代表红眼“因子”(基因),W代表白眼“因子”,雌果蝇有2 条X 染色体,雄果蝇只有1 条X 染色体。摩尔根将亲本白眼雄果蝇表示为WWX,红眼雌果蝇表示为RRXX,白眼雄果蝇产生的精子中均含有W,但一半的精子含有X,一半无X 染色体。红眼雌果蝇产生的配子全部含有R因子和X 染色体,摩尔根认为R与X 染色体紧密偶联,F1的红眼雄果蝇产生的配子为RX和W2 种(不能产生WX和R的配子)。可见摩尔根十分谨慎,并未提出R在X 染色体上、Y 染色体上没有R的假说。
为验证自己的解释,摩尔根还设计了4 组杂交实验(其中有一组与回交实验相同,等于是3 组杂交实验)。第1 组用白眼雌果蝇和白眼雄果蝇杂交,其产生的雄配子应为WX、W,雌配子为WX,杂交子代应全为白眼果蝇,如图2所示。
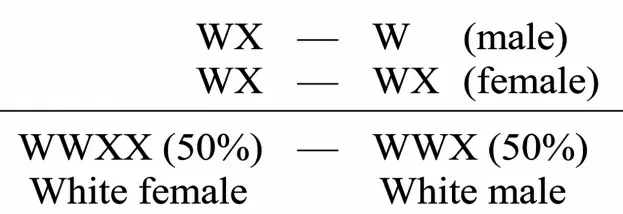
图2 摩尔根对白眼雌果蝇与白眼雄果蝇的杂交解释[5]
第2 组实验用F2的红眼雌果蝇与白眼雄果蝇杂交,由于F2的红眼雌果蝇应该有RRXX和RWXX2 种类型,故杂交结果应该为:有些杂交组合子代应全为红眼,有些杂交组合应出现比例相等的红眼雌性、红眼雄性、白眼雌性和白眼雄性4 种类型。如图3所示。
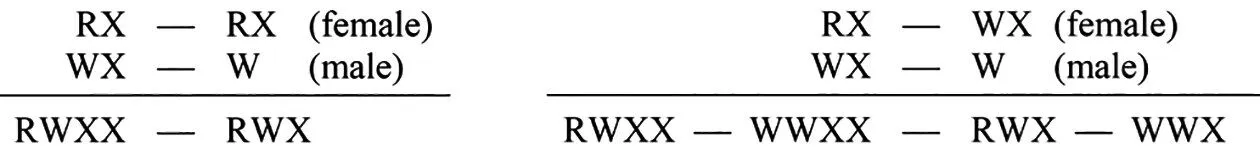
图3 摩尔根对F2 红眼雌果蝇与白眼雄果蝇的杂交解释[5]
第3 组实验用白眼雌果蝇与F1红眼雄果蝇杂交,子代应出现等比例的红眼雌果蝇和白眼雄果蝇,如图4所示。
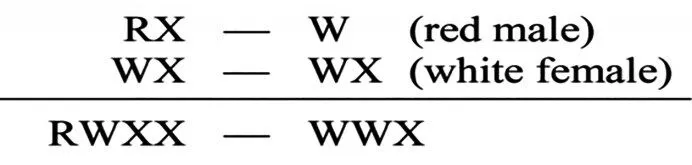
图4 摩尔根对白眼雌果蝇与F1 红眼雄果蝇的杂交解释[5]
上述几组杂交实验结果与预期完全吻合,进而说明了摩尔根假说的合理性。高中《生物》教材中直接用红眼基因位于X 染色体上的假说解释此实验现象,当然事实上确实如此,作此解释也更为科学,但却超出了摩尔根彼时的认识,当时的摩尔根并未接受基因位于染色体上的学说。后续的若干研究成果提供了越来越清晰的实验证据,摩尔根才接受了基因在染色体上的事实。
4 基因在染色体上线性排列的遗传图谱
自摩尔根的白眼雄果蝇之后,又陆续发现了黄体、朱砂眼等突变型果蝇,且发现这些性状与性染色体紧密偶联。摩尔根用黄体红眼雌果蝇与灰体白眼雄果蝇杂交,F1中雄蝇全是黄体红眼、雌蝇全是灰体红眼,F1相互交配得F2,F2中的雄蝇表现为黄体红眼∶灰体白眼∶灰体红眼∶黄体白眼=99∶99∶1∶1,F2中的雌蝇表现为黄体红眼∶灰体红眼=1∶1。经分析可知F2中雄蝇的重组率为1%,用F2中的黄体白眼雄果蝇与Fl中的灰体红眼雌果蝇(双重杂合子)交配,后代不论雌、雄均表现出了重组性状,重组率均是1%。1911年,摩尔根对此现象所作的解释是:上述2 对“因子”均位于性染色体上,即表现为“相引”。“相引”的因子在染色体上呈线性排列,减数分裂时同源染色体联会并发生交叉互换。1911年摩尔根的学生斯特提文特(Sturtevant)想到在染色体上线性排列的基因,可通过子代重组的概率推算出2 个基因之间的距离,并根据此想法,绘制了第1 张染色体的遗传图谱[2],如图5所示。
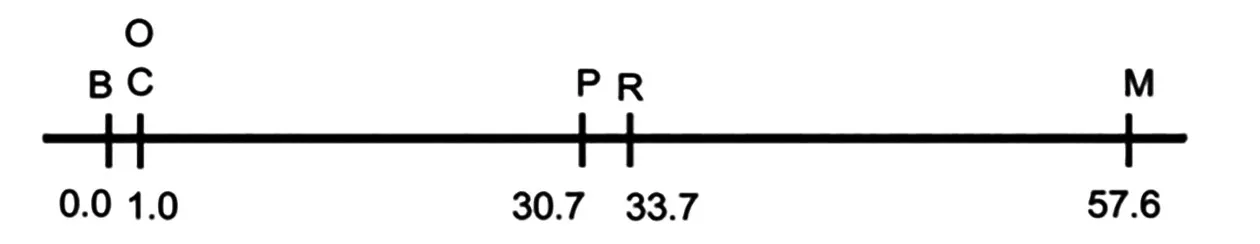
图5 斯特提文特确定的第1 个遗传图谱[2]
5 连锁群
摩尔根依据基因在染色体上线性排列的关系,提出了“连锁群”的概念(位于同一染色体上,相互之间存在连锁遗传关系的基因群)。1911年,发现了果蝇有3 个连锁群,摩尔根由果蝇的染色体组成预测果蝇应有4 个连锁群。1914年穆勒(Muller)发现了果蝇的卷曲翅基因,与无眼基因及此后发现的无刚毛胸基因构成了果蝇的第Ⅳ连锁群[6]。
基于上述实验现象,很多生物学家接受了基因在染色体上的学术观点,但仍有一些人持怀疑态度,包括摩尔根本人也有所担心,所以,需要更为直接的实验证据证明。
6 布里吉斯的果蝇杂交实验
布里吉斯、斯特提文特和穆勒是摩尔根“蝇室”的3 个重要成员,1913年布里吉斯重复了摩尔根的白眼雌果蝇与红眼雄果蝇杂交实验,经科学和耐心的统计,获得了20 496 只子代,其中有10 735 只红眼雌果蝇,9 849 只白眼雄果蝇,但同时有极少数例外情况,即有5 只白眼雌果蝇,7 只红眼雄果蝇(之后称此2 种例外为“初级例外”)。
按照摩尔根的学说,子代雄果蝇应全部为白眼,雌果蝇应全部为红眼,不应出现该例外的情况。布里吉斯通过对果蝇的染色体观察,已知X染色体与常染色体的比值为1 是雌性,比值为1/2是雄性,即雌性为XX,雄性为X,当时并未发现Y 染色体的存在。通过分析可知红眼对白眼为显性,结合之前红眼基因在性染色体上的观点,布里吉斯在解释该例外白眼雌果蝇时,认为其2 条Xw染色体应该均来自于母本白眼雌果蝇,造成该现象的原因是母本雌果蝇在减数分裂时2 条X 染色体未分离,形成了XwXw的卵细胞,该卵细胞与正常减数分裂产生的无X 染色体的精子结合,形成了XwXw白眼雌果蝇。例外的红眼雄果蝇其XW的染色体应来自于父方的XW,该精子与1 个不含X 染色体的卵细胞结合,形成了XW的红眼雄果蝇,而不含X 染色体的卵细胞产生的原因,同样是由于母本雌果蝇在减数分裂时2 条X 染色体未分离导致的,故以上2 种初级例外情况的成因是相同的,出现的概率也是一致的。
1914年,布里吉斯通过更仔细的细胞学观察发现,雄果蝇中还含有1 个Y 染色体,但Y 染色体不决定性别,只决定雄果蝇的可育性,性别取决于X 染色体的数量,即XX、XXY 为雌性,X、XY、XYY 为雄性,XXX 的个体致死,XO 雄果蝇不育。进而指出解释关于初级例外的表述中亲本所产生的不含X 的精子应改为含Y 的精子,推测初级例外的白眼雌果蝇应为XwXwY,并指出该果蝇减数分裂可形成的配子有:XwY、Xw、XwXw、Y,通过细胞学检查发现确实如此[7]。
布里吉斯为进一步验证上述学说,将“初级例外”中的白眼雌果蝇(XwXwY)和红眼雄果蝇(XWY)进行杂交,并预测该杂交的子代中除了有白眼雄果蝇和红眼雌果蝇之外,还将出现少量的“次级例外”,即白眼雌果蝇(XwXwY)和红眼雄果蝇(XWY),同时红眼雌果蝇中有XWXwY、XWXw2 种类型,且比例为1∶1,白眼雄果蝇中有XwYY、XwY2 种类型,且比例为1∶1,如图6所示[8]。实验结果表明该杂交的子代中,出现了约4.3%的次级例外,该例外是野生型染色体组成的白眼雌果蝇和红眼雄果蝇杂交所不会产生的,通过对子代果蝇的细胞学观察,也证实了确实有XYY 和XXY 的染色体组成。
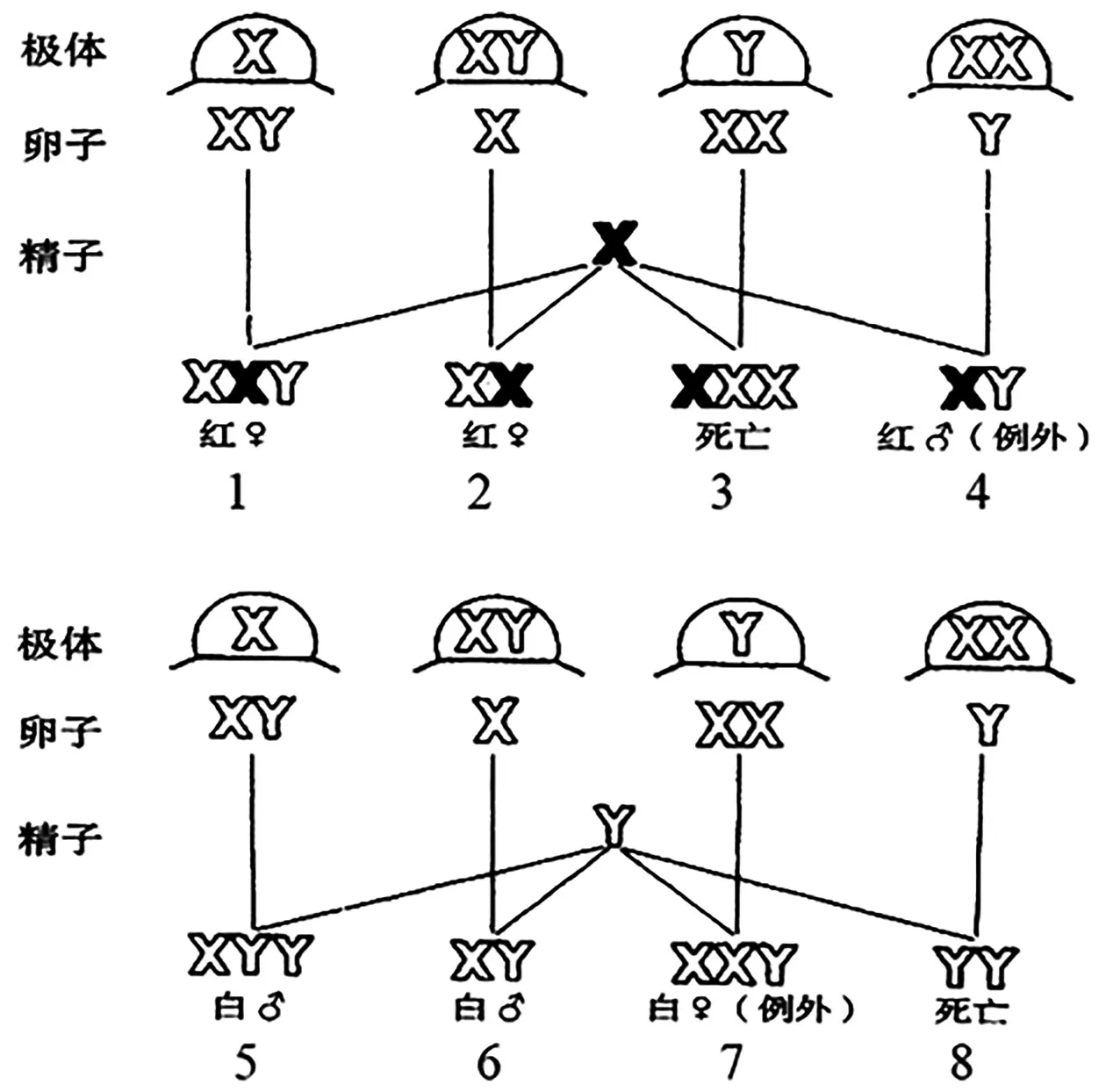
图6 白眼XwXwY 雌果蝇与红眼XWY 雄果蝇杂交[8]
布里吉斯发现的初级例外和次级例外情况,在性染色体上也出现例外,即染色体的行为、性染色体连锁基因的行为、性别决定三者之间完全吻合,这种吻合是很难用偶然巧合解释的,故由此推断性连锁基因位于X 染色体上。
7 黄身雌果蝇与灰身雄果蝇的杂交实验
1922年,摩尔根用纯合黄身雌果蝇与灰身雄果蝇杂交(当时已知灰身对黄身为显性,且均与X染色体相偶联),所产生的F1为黄身雌果蝇与灰身雄果蝇。该结果是之前的学说所不能解释的,通过观察亲本果蝇的染色体组成发现,黄身雌果蝇的X 染色体是由2 条X 染色体相连形成的,同时还有1 条Y 染色体,故其染色体组成可表示为XXY,如图7所示[9]。该果蝇减数分裂所产生的卵细胞为Y和XaXa,灰身雄果蝇产生的精子为XA、Y,故杂交产生的子代有XaXaY、XAY、XAXaXa(致死)、YY(致死),进而解释上述杂交实验现象。该实验表明黄身基因随X 染色体而遗传,基因和染色体的行为又是高度吻合的,进而表明基因在染色体上。

图7 摩尔根所观察的变异黄身雌果蝇的染色体组成[9]
8 染色体的细胞学图像
1928年,苏联科学家杜布赞斯基(Dobzhnsky)在摩尔根实验室用X 射线处理果蝇,让其发生染色体变异,通过对果蝇的装片观察,发现了3 号染色体的一段易位至较小的4号染色体上。杜布赞斯基通过对染色体断裂点的研究分析,绘制出果蝇的3 号染色体的细胞学图像,如图8所示[10]。将该图与基于遗传学实验数据而绘制的基因连锁图比较,发现二者是完全吻合的,进而为基因在染色体上提供了较为直接的证据。
9 果蝇的唾液腺多线染色体
1928年,德国科学家海茨(Heitz)改进了细胞学染色方法,发现染色体的某些部分染色更密集,即染色体会呈现出深浅不同的条带或横纹。1930年,海茨与鲍尔(Bauer)一起合作研究了毛蚊的唾液腺染色体,观察到绳索状结构,并认为是“巨型染色体”。该结果使遗传学家佩因特(Paimter)很受启发,意识到果蝇唾液腺染色体上的条带可用以研究染色体变异中的断裂点,也能作为定位基因在染色体上的坐标,并绘制了果蝇的染色体图(图9)[11]。佩因特发现染色体条带密集处其所含的基因数量也较多(图10)[12],并通过遗传学实验发现,基因的连锁图发生逆转时,染色体条带也发生逆转,遗传上表现易位或缺失时,染色体条带也发生易位或缺失。布里吉斯也以果蝇的唾液腺染色体为材料进行了研究,绘制了果蝇减数分裂中期的染色体图像与唾液腺染色体的对比图像,发现在相同放大倍数下,减数分裂中期的4 号染色体是黑点状,而果蝇唾液腺中的4 号染色体却能看到很多细小条纹(图11)[13]。由此意识到果蝇的唾液腺染色体是研究基因的物质基础的理想材料,并得出了果蝇唾液腺染色体上的基因连锁图,如图12所示[14],该图谱成为了科学家的参考标准。果蝇唾液腺多线染色体的发现,为基因在染色体上提供了最为直观和不可辩驳的证据,在物理层面上直接验证了该结论。
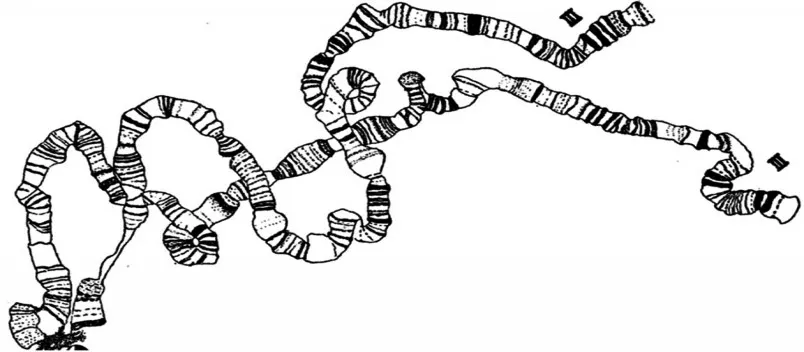
图9 佩因特绘制的果蝇唾液腺多线染色体[11](局部)

图10 佩因特绘制的X 染色体上基因的连锁图[12](局部)
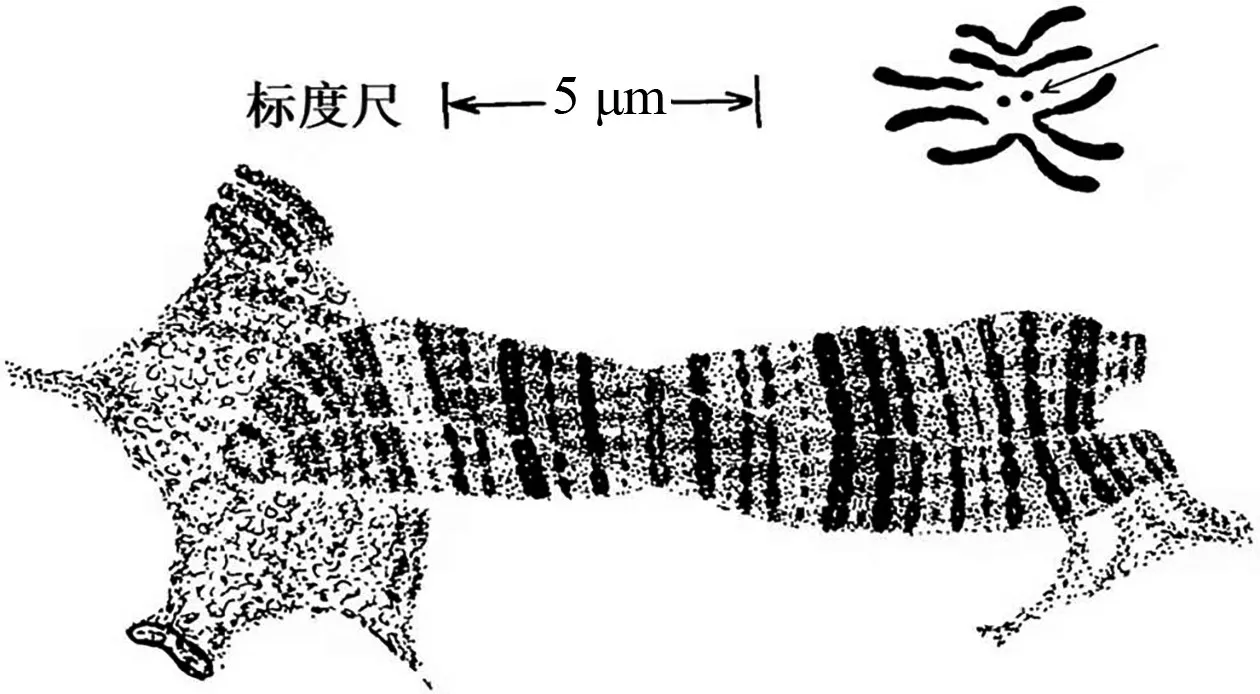
图11 果蝇唾液腺染色体(下部条纹状结构)

图12 布里吉斯绘制的果蝇唾液腺染色体的基因连锁图[14]
果蝇是一个经典的遗传学材料,摩尔根及其“蝇室”的众多成员充分挖掘和利用了果蝇的研究价值。随着科学研究的进行,呈现了越来越多的实验证据,均共同指向了基因在染色体上。但由于基因的微观性及“不可视”性,一开始包括贝特森(Bateson)在内的众多知名科学家,对基因在染色体上的染色体遗传学说表示质疑,甚至包括摩尔根本人当时也不敢肯定基因在染色体上。1933年摩尔根被授予诺贝尔奖时,当年他并未去领奖,不禁让人猜测是因为他所采用的证明方法正是其当年所嘲笑过的孟德尔方法[2]。随着越来越多、越来越直接的证据表明基因在染色体上,一些科学家才开始转变态度,逐渐接受了基因在染色体上,最终使遗传学的研究转移至基因和染色体的方向上。
古人云“智者见于未萌,愚者暗于成事”,在染色体遗传的发展过程中,正是摩尔根、布里吉斯、斯特提文特、杜布赞斯基、佩因特等科学家,将肉眼难以看见的基因通过各种实验,以现象、数据、图像等形式逐渐呈现,进而一步步勾勒和构建出了染色体遗传的理论体系。
猜你喜欢
科学之谜(2021年7期)2021-09-29
作文与考试·小学高年级版(2021年17期)2021-09-23
红领巾·探索(2021年9期)2021-09-14
作文大王·笑话大王(2019年11期)2019-12-02
学生天地·小学中高年级(2018年3期)2018-05-17
杂文月刊(2018年5期)2018-05-14
幼儿教育·父母孩子版(2017年1期)2017-04-05
故事作文·高年级(2016年8期)2016-08-25
红领巾·萌芽(2014年2期)2014-11-26
海外英语(2013年6期)2013-08-27
