
哺乳动物FSHR基因选择压力分析∗
2022-11-22 08:12:12王伟萍
生物学通报 2022年1期
王伟萍 贺 刚 吴 斌,3∗∗
(1 江西省水生生物保护救助中心 江西南昌 330000 2 江西省水产科学研究所 江西南昌 330000 3 南京师范大学生命科学学院 江苏南京 210023)
卵泡刺激素(follicle stimulating hormone,FSH)在促进和维持性腺发育和生殖功能中发挥重要作用,但其生理作用需通过分布于细胞膜表面的卵泡刺激素受体(follicle-stimulating hormone re‐ceptor,FSHR)介导,卵泡刺激素受体是GPCR(Gprotein-coupled receptors)家族的成员[1]。研究发现,FSHR基因与排卵和繁殖力等存在密切关系[2-3]。而检测选择压力的最大似然法检验已被广泛接受,其中,选择系数ω(ω表示非同义/同义置换速率比值)直观地反映了生物体在密码子水平上的适应性进化趋势。ω>1、ω=1 和ω<1 分别代表在进化过程中基因承受正选择、中性选择及负选择(净化选择)[4]。本研究通过对哺乳动物FSHR基因序列进行选择压力分析,分析其适应性进化趋势,为水生哺乳动物,特别是长江江豚(Neophocaena asiaeorientalis)的保护提供参考。
1 材料与方法
1.1FSHR基因序列 基于NCBI(https://www.ncbi.nlm.nih.gov/)上公布的基因序列,选取33 种哺乳动物(11 种鲸豚类、11 种灵长类、11 种其他类),分别代表一般1 胎1 仔的水生类群、一般1 胎1 仔的陆生类群、一般1 胎多仔的典型类群(表1)。

表1 FSHR 基因转录本信息
1.2 方法 首先,通过FasParser2.10.0 软件[5],获得FSHR基因转录本最长开放阅读框序列(open reading frame,ORF),随后通过MEGA X[6]将上述序列对齐。采用DAMBE6.4 软件[7]对FSHR基因转录本最长ORF 序列进行饱和度检测,使用MEGA X中的ML 方法构建系统发育树,并运用PhyloSuite v1.2.2[8]为Paml 程序运行提供所需文件的格式转换[4]。
其次,应用Paml 软件codeml 模块中的4 种不同的模型对FSHR基因进行选择压力分析。本研究采用位点模型中的M0:假设所有位点具有相同的ω值;M1:假设存在2 类位点,保守位点0<ω<1,中性进化位点ω=1,并且估算这2 类位点的比率(p0、p1);M2:假设存在3 类位点,保守位点、中性进化位点、正选择位点ω>1,并且估算这3 类位点的比率(p0、p1、p2);M3:离散模型,假设所有位点的ω值呈离散分布;M7:假设所有位点0<ω<1 且呈现beta 分布;M8:在M7 模型的基础上,增加一类正选择位点(ω>1)。通过M1/M2、M7/M8 重建似然比检验LRT,使用M2 和M8 鉴别受到正选择的位点,分析各位点承受正选择的压力位点。
最后,用枝模型分析水生哺乳动物的FSHR基因是否存在加速进化。枝模型假设系统发育树上的不同枝之间有不同的选择压(即不同的ω值),以检测不同种分枝的正选择压。这里采用枝标签分3 种情况进行标记:1)鲸豚类动物为前景枝;2)灵长类动物为前景枝;3)其他陆生动物为前景枝,均分别用M0 和M2 模型进行相关分析。
2 结果与分析
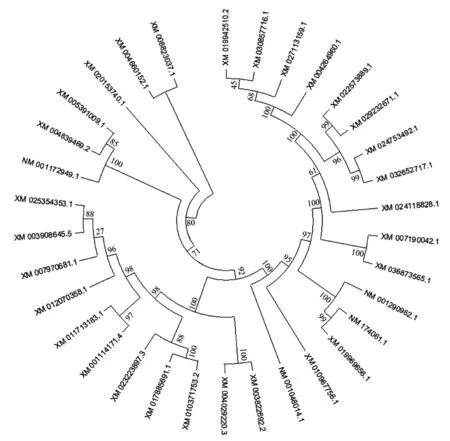
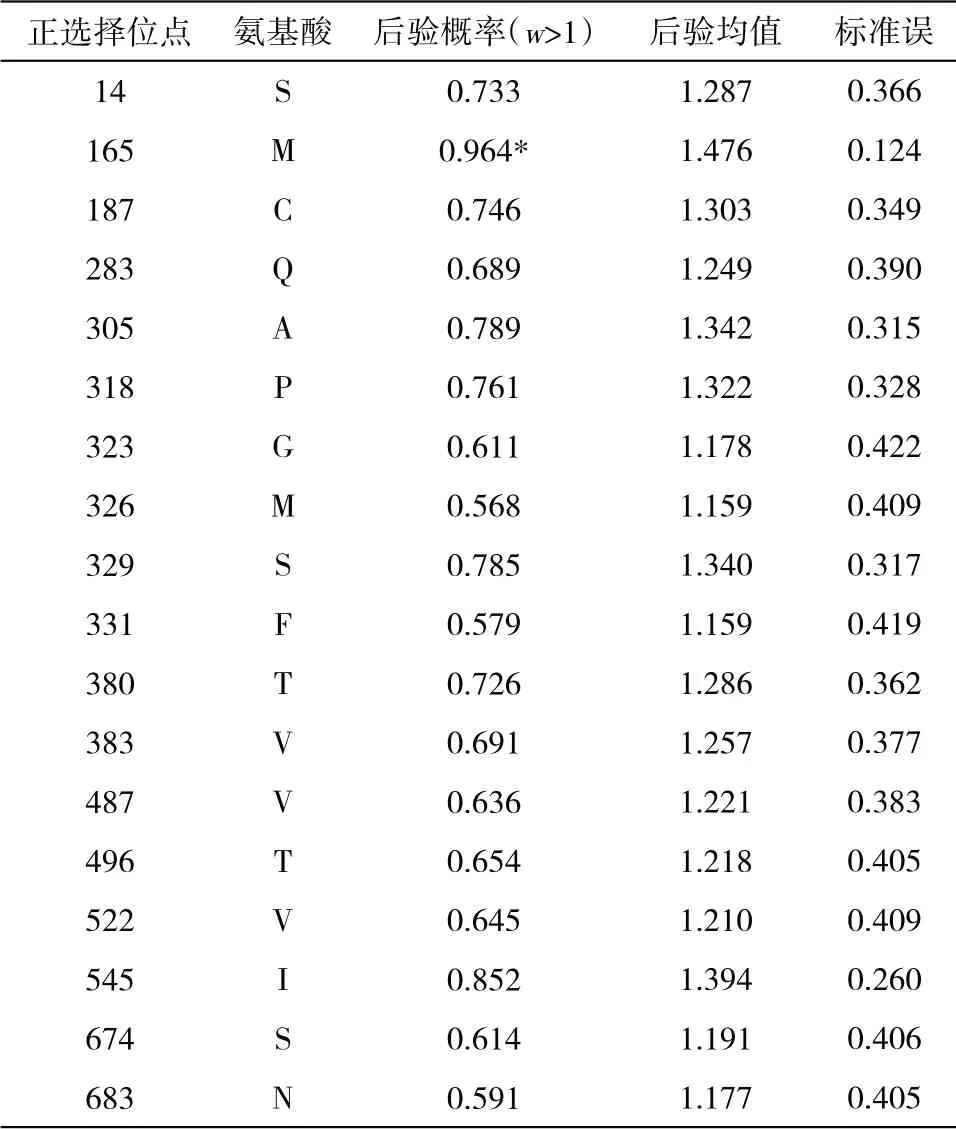
2.1 核苷酸的序列饱和度检测结果 采用DAMBE6.4软件对FSHR基因转录本最长ORF 序列进行饱和度检测,均满足条件P=0.0000,ISS 图1 FSHR 基因转录本最长ORF 序列饱和度分析 2.2 系统发育树的构建 本研究运用MEGAX 中的ML 方法构建FSHR基因转录本最长ORF 序列系统发育树(图2)。构建的系统发育树拓扑结构与传统分类基本一致,且bootstrap 值也基本一致,大多数在90 以上,极少数低于70,推测FSHR基因进化速率适中,基本形成了比较稳定的功能。 图2 FSHR 基因转录本最长ORF 序列构建的系统发育树 2.3 位点模型 比较M0 和M3、M7 和M8,2ΔlnL分别为306.07、20.30,通过χ2检测,差异极显著。但比较M1 和M2,2ΔlnL为0,通过χ2检测,差异不显著。M3 模型结果表明,p0、p1 和p2 分别为0.74904、0.25096 和0.00000,对应的dN/dS(w)为0.04768、0.66855 和25.36134。M1 模型结果表明,p0 和p1 分别为0.83578 和0.16422,对应的dN/dS(w)为0.07879 和1.00000。M8 模型结果表明,以XM_008823037.1 为参考序列,FSHR基因转录本最长ORF 序列正选择位点有19 个,但只有aa 165 M 一个位点的P>95%,表明FSHR基因所承受的正选择压力相对较少,选择压力分析结果见表2。 表2 FSHR 基因正选择位点结果 2.4 枝模型 枝模型检验结果显示,灵长类和其他陆生动物为前景枝,比较M0 和M2,2ΔlnL分别为1.22 和1.45,通过χ2检测,差异均不显著。而鲸豚类动物为前景枝,比较M0 和M2,2ΔlnL为5.60,通过χ2检测,差异显著。即M2 模型优于M0 模型,不同的枝有不同的选择压力,且二者均为0<ω<1,表明鲸豚类动物FSHR基因存在加速进化,结果见表3。 表3 枝模型的似然比检验和种系选择压结果 动物对环境的适应及适应性进化一直是研究热点问题,高度保守的繁殖相关基因的进化系统研究能简便有效地揭示出其潜在的生物学功能。例如,鼬科、浣熊科、小熊猫科和臭鼬科均出现胰液核糖核酸酶基因复制,且检测到正选择作用和适应性位点,同时结合RNA 组织表达等实验结果,表明该基因不仅有食性相关的功能,可能还有其他功能[9]。动物采取不同的繁殖策略以适应所处环境,通常由行为及内分泌共同调节。有研究表明,同一单元内雌性川金丝猴(Rhinopithecusroxellana)可能通过季节层面地错开生殖状态、怀孕机会及邀配行为的方式以减缓生殖竞争并适应环境的季节性变化[10]。有研究表明,FSHR基因含量的高低可能与哺乳动物的繁殖力之间存在一定相关性[3],FSHR基因正常表达是卵巢获得促性腺激素反应能力的关键,受体的缺失会引起卵泡不能正常发育,严重的则会引起繁殖力降低甚至不孕[11]。通过对33 种哺乳动物的FSHR基因的选择压力检测,发现M3 模型中p2 为0,表明FSHR基因主要承受的选择压力是净化选择。Kamath和Getz[12]研究指出,在长期的进化过程中选择压力不可能一直效应于整个基因中,而是作用于某些特殊效用位点上,例如,某个基因编码区域的特定位点。因此,FSHR基因的净化选择可能主要是FSHR基因对哺乳动物至关重要,较保守。但在FSHR基因也检测出多个正选择压力位点,选择信号是选择作用遗留在基因上的印迹。任何一种生物想要生存下去,都必须不断地进行繁殖,保证自己所属物种的数量,繁殖是动物实现自身适合度的方法[13]。这可能是为了应对不同的环境及不同的婚配制度带来的多重挑战,保证物种的可持续性发展,动物通常采用不同的繁殖策略与行为策略以提高自身的交配机会、成功率及后代的存活率[14]而发生正向选择突变。对鼠类研究指出,无论是依赖光周期的天算式繁殖,还是综合实际水热和食物条件的鼠算式繁殖,都显示出鼠繁殖策略的灵活性。鼠对环境调节的有效把握和相对较快的传代速度使其更有可能演化出新的适应策略[15]。水生哺乳动物,例如,长江江豚无论在野外还是在人工环境下均无产双胞胎的记录,长江江豚的交配可能发生在一年中的大多数月份,并可能有一个以上的交配高峰,抚幼行为比较常见[16],因此,推测鲸豚类FSHR基因的加速进化可能有助于其实现繁殖适应。鲸豚类作为物种演化历史上一类非常重要的次生性水生哺乳动物,成功地适应了水生环境并且快速地辐射进入到全球的各水域。为了适应水生生境,鲸类外部形态和生理、生态产生了一系列的适应性改变,在表型性状上发生了明显变化,特别是在繁殖、生长等性状上差异较大,但由进化所导致繁殖适应的遗传机制尚不清楚,缺乏系统研究。一个种群繁殖力的强弱直接影响该种群以后的发展前途及最终命运[17],因此,对以长江江豚为代表的鲸豚类动物繁殖适应策略的深入研究有利于科学开展相关保护工作。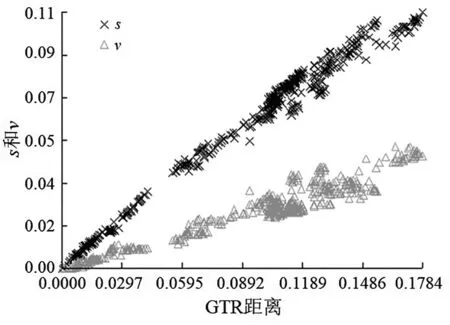

3 讨论
猜你喜欢
上海金属(2021年6期)2021-12-02 10:47:20
昆明医科大学学报(2021年3期)2021-07-22 07:40:04
生物学通报(2019年3期)2019-02-17 18:03:58
小学科学(学生版)(2018年5期)2018-06-15 08:48:00
奇妙博物馆(2018年12期)2018-04-25 02:16:04
奇妙博物馆(2018年12期)2018-04-25 02:16:02
高中生·天天向上(2016年11期)2017-02-28 07:40:18
环境科技(2016年4期)2016-11-08 12:18:58
长江蔬菜(2014年1期)2014-03-11 15:09:48
椰城(2013年4期)2013-11-16 06:31:11
