
解除生殖滞育的巴氏新小绥螨捕食能力
2022-11-18 08:26:16李晓萌唐庆峰
中国生物防治学报 2022年5期
李晓萌,李 玲,高 超,唐庆峰*
(1.安徽农业大学植物保护学院/作物有害生物综合治理安徽省重点实验室,合肥 230036;2.安徽省科技厅农村与社会科技发展中心,合肥 230091)
巴氏新小绥螨是多种叶螨、蓟马等小型吸汁性害虫的天敌,具有食性广、发育历期短、死亡率低、繁殖能力和扩散能力强等优势,被广泛应用于生物防治之中[1]。已经实现了规模化饲养与商业化应用,成为国内生产量最大的本土捕食螨种[2]。
螨类发育过程受到多种环境因素和食物的影响,光周期是影响巴氏新小绥螨各螨态发育的主要环境因素,短光照可以诱导巴氏新小绥螨滞育[3-6]。董婷婷[7]报道巴氏新小绥螨存在经常性的生殖滞育现象。近年关于螨类的滞育在生理学、分子生物学、形态学、行为学等方面有诸多研究。螨类在进入滞育之后,如果不出现诱导其滞育解除的特殊处理或生物环境因素,将长时间处于代谢低下、发育迟缓或停止的状态,但滞育之后的螨类抗逆性增强,可以长时间存活[8,9]。可以利用其这一特性,在储存运输过程中,先诱导产生生殖滞育,降低其生命活性,增强其抗逆性,提高存活率,在需要大量种群时再诱导其解除滞育,大量繁殖,这一方法能够极大延长货架期,保证种群数量的稳定。
1 材料与方法
1.1 供试虫源
巴氏新小绥螨和腐食酪螨由中国农业科学院植物保护研究所提供,实验室连续饲养3代以上,建立稳定种群。
1.2 饲养条件
巴氏新小绥螨普通品系:25 ℃±1 ℃、RH:80%±5%、光周期14L∶10D;滞育品系:18 ℃±1 ℃、RH:80%±5%、光周期8L∶16D;滞育解除品系:28 ℃,光周期14L∶10D。光照强度均为2000 lx。
1.3 饲养方法
1.3.1 群体饲养 将提供的巴氏新小绥螨放入直径8 cm,高12 cm的500 mL圆筒保鲜盒内,以纱布封口。保鲜盒内铺入5 cm左右经过高温灭菌后的麦麸,以麦麸饲喂替代猎物腐食酪螨,再以腐食酪螨饲喂巴氏新小绥螨。每3 d喷一次无菌水,搅拌均匀后置于光照培养箱中。
1.3.2 小室饲养 将用亚克力板定制的小室两端用燕尾夹固定。每小室饲喂一头巴氏新小绥螨。将小室置于(450×360×45)mm中号托盘中,托盘中间放置大块海绵片,海绵片上放置浸水滤纸保湿。
1.3.3 试螨收集 取巴氏新小绥螨、腐食酪螨与麦麸混合物放入漏斗中,固定于烤虫架,置于200 W白炽灯下20 cm,烤大约2 h左右,如螨落下不多,可用勺子对混合物进行搅拌,直至收集完成。
1.4 不同品系巴氏新小绥螨对替代猎物腐食酪螨各螨态的取食选择作用
根据预试验中巴氏新小绥螨对腐食酪螨捕食量的试验结果,试验共设3个处理组,分别为滞育品系:18 ℃,光周期8L∶16D;普通品系:25 ℃,光周期14L∶10D;解除滞育品系:28 ℃,光周期14 L∶10D;光照强度均为2000 lx,RH 80%±5%。分别选取各品系的巴氏新小绥螨成螨,饥饿处理24 h后备用。在每个凹形小室中放入经过24 h饥饿处理后的滞育品系、普通品系、滞育解除品系巴氏新小绥螨雌成螨1头和替代猎物腐食酪螨卵、幼螨、若螨和成螨各10头,分别置于各处理的光照培养箱中,重复3次。24 h后观察记录不同品系巴氏新小绥螨雌成螨对腐食酪螨各螨态的捕食量。
1.5 不同品系巴氏新小绥螨对替代猎物腐食酪螨捕食功能反应
根据预试验中巴氏新小绥螨对腐食酪螨捕食量的试验结果,试验共设3个处理组,同1.4。每个处理组分别放入5头、10头、15头、20头、25头腐食酪螨幼螨于5个凹形小室中,每个小室放入1头经24 h饥饿处理的滞育品系、普通品系、滞育解除品系的巴氏新小绥螨雌成螨,盖上亚克力板,用长尾夹固定两端,放置在盛有海绵覆盖滤纸的瓷盘(加水保湿)上,置于光照培养箱中,3次重复。24 h后观察记载捕食量。
桩号计算主要包括计算位于ZY、QZ、YZ间曲线距离整数倍的里程桩号,其可以通过计算离这3个数值中最小整数倍进行获取。获取ZY与QZ之间整数倍的代码如下:
1.6 数据统计与分析
1.6.1 选择取食作用 天敌对猎物嗜食性的强弱可用选择系数Q表示:Q=某螨态被取食数占总取食数百分比/某螨态数占猎物总数百分比,Q>1时,捕食螨对该猎物的螨态表现为嗜食性;Q<1时,为非嗜食性;当Q=1时,为随机捕食[10]。
1.6.2 捕食功能反应 采用Holling圆盘方程Na=aTN/(1+aThN)对数据进行拟合。N:猎物初始密度;Na:猎物被捕食量;a:瞬时攻击率;T:捕食者总利用时间(T=1 d);Th:处理1头猎物所需要的时间;a/Th:捕食者的捕食能力;1/Th:日最大捕食量[11,12];Nt:替代猎物密度;1/Na:每日捕食量。
应用SPSS 22.0、GraphPad Prism、DPS等数据处理系统综合处理。用最小二乘法估算出模型的Th、1/Na、Nt等参数,通过已知和求得的数据,拟合出曲线以及曲线方程,由计算出的理论值与试验获得的数据之间采用χ2检验,采用Duncan氏新复极差法进行差异显著性检验。
2 结果与分析
2.1 巴氏新小绥螨对替代猎物腐食酪螨各螨态的选择性
在猎物各螨态数量和比例相同的情况下,巴氏新小绥螨普通品系对腐食酪螨不同螨态的选择性和被食比例存在显著差异,捕食腐食酪螨卵、幼螨、若螨和成螨的被食比例分别为26%、43%、21%和10%,选择系数分别为1.05±0.15、1.70±0.23、0.82±0.23和0.41±0.17,表现出对猎物幼螨具有明显的嗜食性,对猎物的卵为随机选择性,对猎物的若螨和成螨非嗜食性(表1)。

表1 巴氏新小绥螨普通品系对腐食酪螨各螨态的捕食选择性(25 ℃,光周期14L:10D)Table 1 Prey-stage preference of N.barkeri direct development stain for T.putrescentiae (25 ℃, Photoperiod 14L:10D)
在猎物各螨态数量和比例相同的情况下,巴氏新小绥螨解除滞育品系对腐食酪螨不同螨态的选择性和被食比例存在显著差异,捕食腐食酪螨卵、幼螨、若螨和成螨的被食比例分别为25%、41%、22%和12%,选择系数分别为1.01±0.11、1.66±0.37、0.86±0.13和0.48±0.17,表现出对猎物幼螨具有明显的嗜食性,对猎物的卵为随机选择性,对猎物的若螨和成螨非嗜食性(表2)。

表2 巴氏新小绥螨滞育解除后对腐食酪螨各螨态的捕食选择性(28 ℃,光周期14L∶10D)Table 2 Prey-stage preference of N.barkeri post-diapause stain for T.putrescentiae (28 ℃, Photoperiod 14L:10D)
在猎物各螨态数量和比例相同的情况下,巴氏新小绥螨滞育品系对腐食酪螨不同螨态的选择性和被食比例存在显著差异,捕食腐食酪螨卵、幼螨、若螨和成螨的被食比例分别为21%、48%、24%和7%,选择系数分别为0.82±0.20、1.94±0.20、0.96±0.29和0.29±0.16,表现出对猎物幼螨具有明显的嗜食性,对猎物的卵、若螨和成螨非嗜食性(表3)。
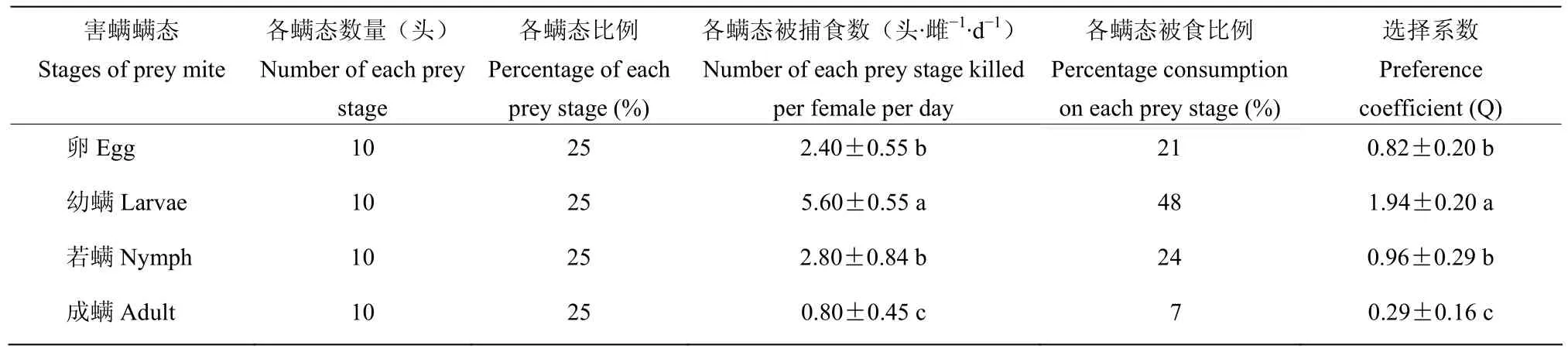
表3 巴氏新小绥螨滞育品系对腐食酪螨各螨态的捕食选择性(18 ℃,光周期8L∶16D)Table 3 Prey-stage preference of N.barkeri diapause stain for T.putrescentiae (18 ℃, Photoperiod 8L∶16D)
普通品系、滞育解除品系与滞育品系的巴氏新小绥螨对腐食酪螨不同螨态的捕食选择性和被食比例呈现出基本一致的趋势,均表现出强烈的选择性。三个品系的巴氏新小绥螨均对猎物幼螨表现出嗜食性,其选择系数从高到低为幼螨,卵,若螨,成螨;但也表现出一定的差异性,普通品系与滞育解除品系对猎物的卵和幼螨均表现为嗜食性,滞育品系的巴氏新小绥螨仅对猎物的幼螨表现为嗜食性。三个品系的巴氏新小绥螨对腐食酪螨不同螨态的被食比例均以幼螨最高,均达40%以上,成螨最低,均低于12%(表1~3)。
2.2 巴氏新小绥螨对替代猎物腐食酪螨各螨态的日捕食量
普通品系、滞育品系、滞育解除品系的巴氏新小绥螨对替代猎物腐食酪螨的日均捕食量存在显著差异,并表现出基本一致的趋势。捕食猎物幼螨的日均捕食量最大,滞育解除品系、普通品系和滞育品系幼螨的日均平均捕食量分别为6.8头、5.8头和5.6头;普通品系对于替代猎物腐食酪螨卵与成螨的日均捕食量略高于滞育品系,滞育解除品系的巴氏新小绥螨对替代猎物腐食酪螨的卵,幼螨,若螨,成螨的日均捕食量均高于滞育品系和普通品系,表明诱导滞育品系的巴氏新小绥螨解除滞育后,对猎物的捕食量无不利影响(图1)。

图1 普通品系、滞育品系、滞育解除品系巴氏新小绥螨对腐食酪螨不同螨态的捕食量Fig.1 Prey consumption variation among three strains of N.barkeri (direct development strain, diapause strain and post-diapause strain)toward individual life stages of T.putrescentiae
2.3 巴氏新小绥螨对替代猎物腐食酪螨的捕食功能
在普通品系巴氏新小绥螨对不同密度替代猎物腐食酪螨的捕食试验中,当Nt为5头、10头、15头、20头、25头时,1/Na(捕食量)对应为2.60±0.89、5.00±1.00、5.80±0.84、6.20 ±0.45、6.60±0.89,用最小二乘法拟合的方程为1/Na=1.479×(1/Nt)+0.078,通过方程以及拟合的曲线,可以发现:随着替代猎物腐食酪螨的密度不断增加,巴氏新小绥螨的日捕食量先上升再逐渐趋于平缓,捕食功能符合Holling Ⅱ型模型(图2)。
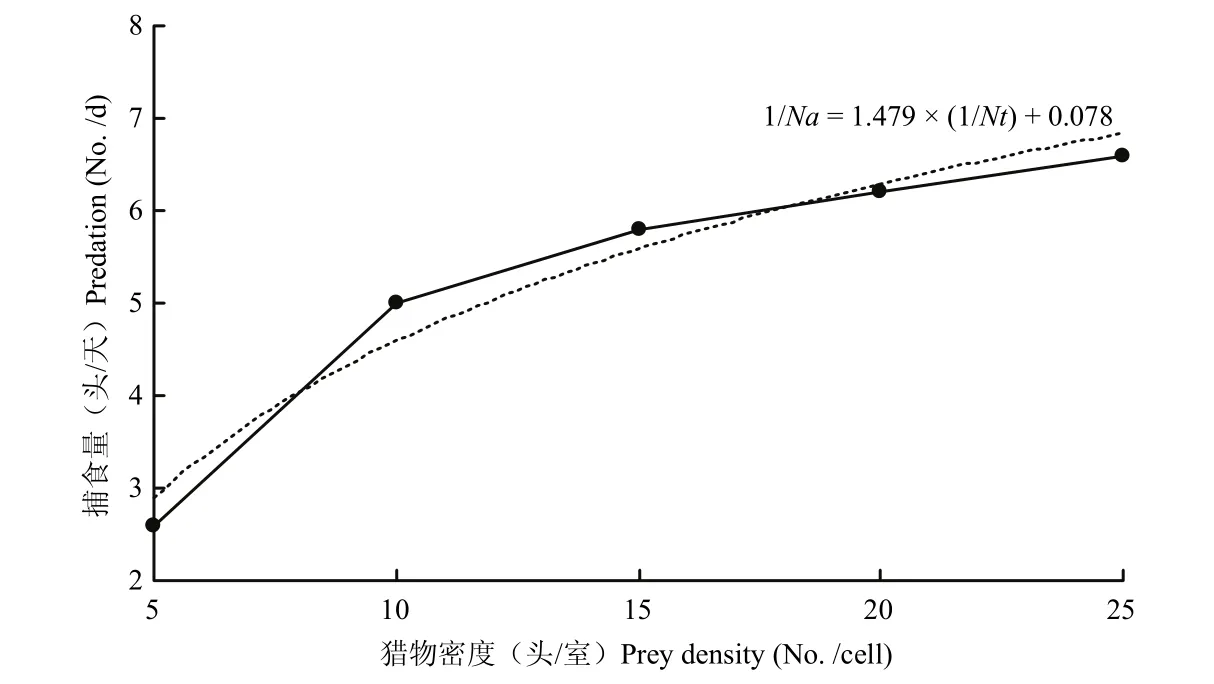
图2 普通品系巴氏新小绥螨对腐食酪螨捕食方程拟合曲线Fig.2 The fitting curve of function response of N.barkeri direct development strain to T.putrescentiae
在滞育品系巴氏新小绥螨对不同密度替代猎物腐食酪螨的捕食试验中,当Nt为5头、10头、15头、20头、25头时,1/Na(捕食量)对应为1.20±0.45、2.40±0.55、3.20±0.45、3.60±1.14、3.80±0.84,用最小二乘法拟合的方程为1/Na=3.662×(1/Nt)+0.083,通过方程以及拟合的曲线,可以发现:随着替代猎物腐食酪螨的密度不断增加,滞育品系巴氏新小绥螨的日捕食量先上升再逐渐趋于平缓,捕食功能符合Holling Ⅱ型模型(图3)。
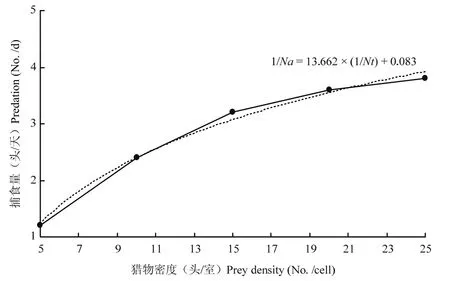
图3 滞育品系巴氏新小绥螨对腐食酪螨捕食方程拟合曲线Fig.3 The fitting curve of function response of N.barke diapause strain to T.putrescentiae
在滞育解除品系巴氏新小绥螨对不同密度替代猎物腐食酪螨的捕食试验中,当Nt为5头、10头、15头、20头、25头时,1/Na(捕食量)对应为1.20±0.45、2.40±0.55、3.20±0.45、3.60±1.14、3.80±0.84,用最小二乘法拟合的方程为1/Na=1.546×(1/Nt)+0.069,通过方程以及拟合的曲线,可以发现:随着替代猎物腐食酪螨的密度不断增加,滞育解除品系的巴氏新小绥螨的日捕食量先上升再逐渐趋于平缓,所以我们可以得出,捕食功能符合HollingⅡ型模型(图4)。
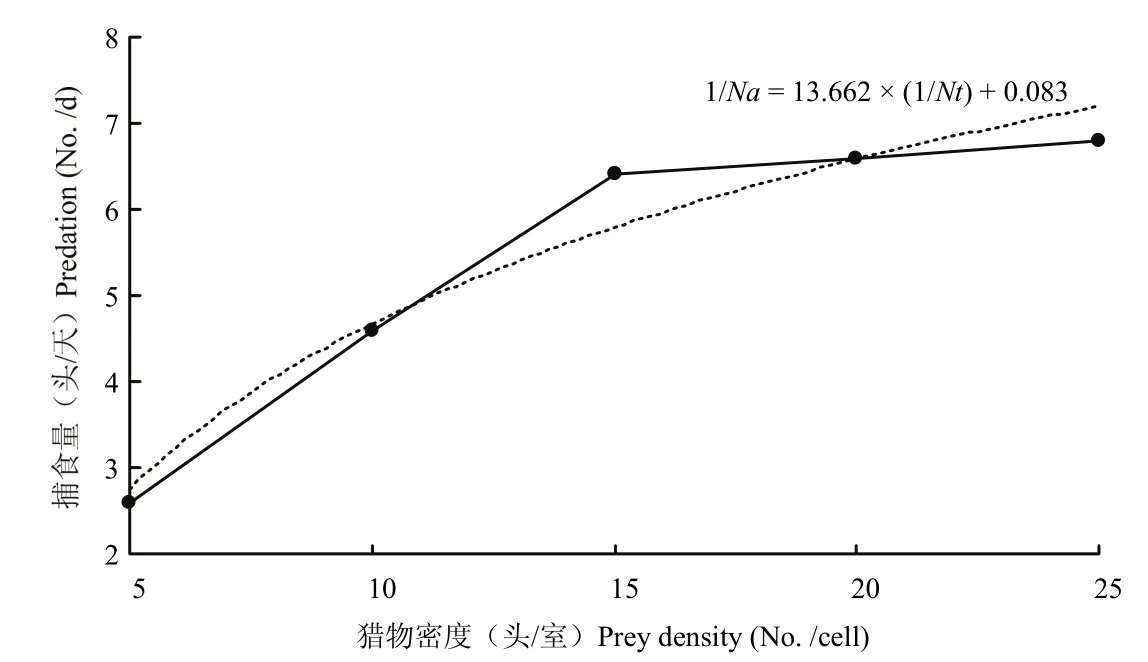
图4 滞育解除品系巴氏新小绥螨对腐食酪螨捕食方程拟合曲线Fig.4 The fitting curve of function response of N.barkeri post-diapause strain to T.putrescentiae
巴氏新小绥螨普通品系、滞育品系、滞育解除品系捕食腐食酪螨的功能反应随着猎物密度的增加,捕食者捕食量增加,但当猎物密度达到一定范围时,捕食者捕食量不增加,趋于稳定,均可以很好地拟合Holling Ⅱ型圆盘方程(图2~4)。
将三个品系巴氏新小绥螨的捕食功能方程及参数列于表4。表4显示:普通品系、滞育品系、滞育解除品系的巴氏新小绥螨对猎物腐食酪螨的攻击系数a分别为0.6761、0.2731、0.6468;理论日最大捕食量分别为12.8205头、12.0482头、14.4928头,理论日最大捕食量滞育解除品系大于普通品系与滞育品系,滞育品系与普通品系理论日最大捕食量无显著差异;三个品系的处理时间Th分别为0.0780 d、0.0830 d和0.0690 d;Holling 圆盘方程分别为Na=0.6761N/(1+0.0527N)、Na=0.2731N/(1+0.0227N)、Na=0.6468N/(1+0.0446N)。
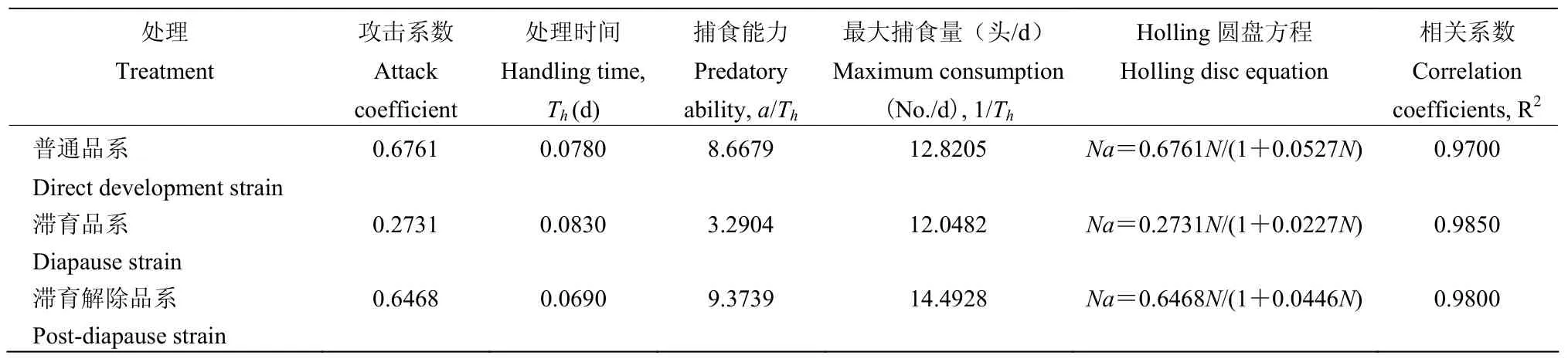
表4 巴氏新小绥螨普通品系、滞育品系、滞育解除品系捕食腐食酪螨的功能反应参数Table 4 The functional response parameters of N.barkeri direct development strain, diapause strain and post-diapause strain preyed T.putrescentiae
瞬时攻击系数a与处理时间Th之比可以作为天敌对猎物捕食能力的标准,a/Th的值越大,说明对猎物的捕食能力越强,滞育解除品系、普通品系、滞育品系的巴氏新小绥螨对猎物的捕食能力a/Th分别为9.3739、8.6679、3.2904,三个品系的巴氏新小绥螨捕食能力由大到小为滞育解除品系,普通品系,滞育品系,表明诱导生殖滞育,对滞育解除后的巴氏新小绥螨捕食能力无不利影响,且有一定程度的增强。
3 讨论
巴氏新小绥螨作为广食性捕食螨类,可用来防治粉螨、叶螨、粉虱和蓟马等[13]。本文对巴氏新小绥螨普通、滞育以及滞育解除品系的选择捕食作用与捕食功能进行了探究。巴氏新小绥螨普通品系与滞育解除品系对替代猎物腐食酪螨不同螨态的捕食选择性无显著差异,均对腐食酪螨的幼螨和卵具有嗜食性,对若螨和成螨为非嗜食性;滞育品系的巴氏新小绥螨只对腐食酪螨的幼螨具有嗜食性。滞育品系的巴氏新小绥螨理论日最大捕食量与普通品系及滞育解除品系无明显差异,但由于滞育品系缺乏对卵的嗜食性,因此在应用中可能会存在防效降低的隐患。
生殖滞育的发生与其他滞育方式有着显著的区别,巴氏新小绥螨滞育方式为典型的生殖滞育,即雌成螨滞育后不产卵,并且在外界环境恢复到适宜发育的条件下,滞育解除仍有一定的间隔期[7]。
大草蛉Chrysopa pallens(Rambur)的滞育存在生殖代价,滞育持续时间影响滞育解除后的部分生物学特性,滞育后预蛹的存活率显著降低,蛹期和产卵前期显著延长,雌成虫寿命显著缩短,成虫鲜重和单雌产卵量显著下降[14],本研究显示巴氏新小绥螨在解除滞育后,对害螨的控制能力不仅没有同大草蛉一样付出一定的生殖代价,反而更强于未滞育前。这可能与螨类和昆虫生理代谢的差异性有关,需要进一步研究。
本试验是在实验室条件下的测定结果,有待进行田间试验,以取得生产实际的应用价值。
猜你喜欢
遗传(2023年9期)2023-09-25 09:31:44
农业科技通讯(2023年1期)2023-02-12 07:08:50
当代水产(2021年10期)2022-01-12 06:21:02
中国果业信息(2021年7期)2021-12-01 20:20:32
上海包装(2019年2期)2019-05-20 09:10:56
少儿科学周刊·少年版(2017年7期)2017-09-29 08:02:41
少儿科学周刊·儿童版(2017年7期)2017-09-29 02:22:58
中国乳业(2016年4期)2016-11-07 09:50:29
广东石油化工学院学报(2016年3期)2016-05-17 05:16:24
天津科技大学学报(2016年1期)2016-02-28 16:59:44
