
茶树远缘杂交生物碱和儿茶素遗传变异研究
2022-11-15 08:53:58周梦珍晏嫦妤黄亚辉
茶叶通讯 2022年2期
罗 莉,李 敏,曾 贞,周梦珍,成 晨,晏嫦妤,黄亚辉*
1. 华南农业大学 园艺学院/茶学省部共建重点实验室,广东 广州 510642;2. 广东财经职业技术学校,广东 佛山 528231
茶的生化特性,特别是化学成分,是规定其它各项特性的基础成分[1]。其中,生物碱和儿茶素是茶中极为重要的化学成分。从群体意义上,茶树中生物碱和儿茶素的分布反映茶树的起源特点与演化路径[1],也是茶组植物种群分类的重要依据[2];从个体意义上,茶叶中生物碱、儿茶素的组成及其在加工过程中的转化,决定茶叶色、香、味、形的品质特征,影响茶叶的保健功效[3]。
茶组植物的生物碱以嘌呤生物碱为主,目前发现的嘌呤生物碱主要有咖啡碱(CAF)、可可碱(TBR)、茶叶碱(TP)和苦茶碱(TA)。茶组植物不同种的嘌呤生物碱分布模式不同:茶种(Camellia sinensis)以咖啡碱为优势生物碱,含少量可可碱和茶叶碱;毛叶茶(Camellia ptilophylla)和滇缅茶(Camellia irrawadiensis)以可可碱为优势生物碱[4],含少量或不含咖啡碱;苦茶(Camellia assamica var. kucha)以苦茶碱为优势生物碱[5],含一定量的咖啡碱和可可碱。
儿茶素类是茶叶多酚类化合物的主体成分,含量一般占绿茶干重的12% ~ 24%[6]。根据C环3位上是否连有没食子基团,儿茶素分为酯型儿茶素(也称复杂儿茶素)和非酯型儿茶素(简单儿茶素)[7]。没食子酸(GA)是茶多酚的重要组成单元,常以酯的形式连接在儿茶素的3位羟基上,形成一系列的酯型儿茶素衍生物[8]。儿茶素或表儿茶素在植物界分布极为广泛,而酯型儿茶素除了在葡萄中有微量存在以外,只有在茶树中大量存在,并且茶树中酯型儿茶素的含量远远高于非酯型儿茶素[3]。至今在茶鲜叶和绿茶中,发现的单体儿茶素有20多种,其中大量存在的主要是四种:表没食子儿茶素没食子酸酯(EGCG)、表没食子儿茶素(EGC)、表儿茶素没食子酸酯(ECG)和表儿茶素(EC)[6]。
目前,由于远缘杂交不亲和的特性,茶组植物杂交的研究主要集中于茶种内不同栽培品种之间,如罗小梅等[9]将福鼎大白茶、桂绿一号等8个茶树品种设计不同杂交组合,发现各品种间花粉生活力差异极显著,杂交结实率在5.0% ~ 28.5%。茶组内不同种间或茶组与山茶属其它组内物种之间杂交种群的研究甚少,且这些为数不多的远缘杂交试验育得的子代样本量也较少[10-12]。充分了解茶树生物碱和儿茶素在远缘杂交亲子代间的遗传变异规律,可为育成较大量远缘杂交后代样本数提供理论依据。
本试验研究以金萱(Camellia sinensis cv.Jinxuan)和南昆山毛叶茶(也称“可可茶”,Camellia ptilophyllaChang)的远缘杂交亲子代种群为材料[13],其双亲的生物碱和儿茶素分布模式及遗传物质基础都有较大差异,F1代样本量较大(64个)。同时,由于金萱(♀)适应环境能力强,种植成活率高,生长势壮旺[14-15];而南昆山毛叶茶(♂)离开原始分布区后,成活能力较差[4]。对本试验种群生物碱和儿茶素性状的研究,不仅有利于揭示茶树远缘杂交生物碱和儿茶素的遗传变异规律,还有利于发展F1代茶树早期鉴定,促使茶种与毛叶茶种互相取长补短,以期将来从杂交子代中选育出既有低咖啡因特色、又有较高儿茶素含量及适制好茶且能广泛栽培种植的茶树新品种。
1 材料与方法
1.1 材料与试剂
南昆山毛叶茶单株(♂)、华南农业大学校内种质资源圃的金萱(♀)及其种间远缘杂交育成的 64 份F1代全同胞系的一芽二叶新梢。
13种化学物质的单体标准品,均购于美国Sigma-Aldrich公司;异丙醇(AR级)、三氟乙酸,购于天津市科密欧化学试剂有限公司;甲醇(HPLC级),购于美国Thermo Fisher公司;甲醇、乙醇,均为国产分析纯。
1.2 仪器设备
Agilent 1260高效液相色谱仪,美国Agilent公司;分析天平、微波超声机,昆山市超声仪器有限公司;AB204-N型电子天平,上海梅特勒-托利多仪器有限公司;DK-8D电热恒温水浴锅,北京市永光明医疗仪器厂;循环水式真空泵(SHZ-D Ⅲ),巩义市予华仪器有限责任公司。
1.3 实验方法
1.3.1 样品处理
将采摘的各供试材料一芽二叶新梢杀青固样,于103℃烘箱中烘至足干,磨碎并过筛,分别称取0.6 g磨碎干茶样于锥形瓶,100℃水浴浸提45 min,定期摇匀,趁热抽滤,冷却后定容至100 mL,过分子筛后注入液相进样瓶。
1.3.2 HPLC分析条件
生物碱:色谱柱为 Agilent Eclipse XDB-C18(4.6×150 nm,5 μm),柱温 38℃,进样量 10 μL,流速 1.0 mL/min,检测波长 230 nm,流动相A:0.1%的三氟乙酸水溶液,流动相B:纯甲醇。洗脱梯度(以A相的变化为准):0 ~ 20 min,保持 90%;20 ~ 30 min,90% ~ 80%;30 ~35 min,80% ~ 20%。
儿茶素:色谱柱为 Agilent Eclipse XDB-C18(4.6×150 nm,5 μm),柱温 38℃,进样量 10 μL,流速0.8 mL/min,检测波长278 nm,流动相A:0.1%的三氟乙酸水溶液,流动相B:纯甲醇。洗脱梯度(以A相的变化为准):0 ~ 30 min,90% ~ 70%;30 ~ 40 min,70% ~ 20%。
1.3.3 数据处理
采用1.3.1方法提取供试样品生物碱和儿茶素后,按照1.3.2分析条件进行HPLC测定,每个试验重复3 次,以3次重复的平均值±标准差表示各试验结果,利用Excel 2016和 SPSS 22.0进行统计分析。进行皮尔逊相关性分析,并对高度相关的成对变量建立回归模型;利用聚类或主成分分析,简化及综合评价各指标及样本;利用相对遗传力和杂种优势理论分析各指标的遗传模型[13,16]。
2 结果与分析
1.1 茶树远缘杂交生物碱的遗传变异
1.1.1 亲子代生物碱的组成与含量
由表1可见,南昆山毛叶茶(♂)、金萱(♀)和F1代全同胞系的生物碱分布模式为:富含可可碱(4.25%),不含其它生物碱;较高含量的咖啡碱(3.16%)和较低含量的可可碱(0.36%),不含茶叶碱和苦茶碱;较高含量的咖啡碱(2.48%~ 5.84%)和一定量的可可碱(0.16% ~ 1.70%),不含茶叶碱和苦茶碱。

表1 父母本及F1代全同胞系生物碱的组成和含量Table 1 The composition and content of alkaloids in parental and F1 progeny of full-sib
64个F1代个体之间生物碱含量指标表现出较大的变异分化,尤其是在可可碱的含量和咖啡碱与可可碱的比值这2个指标上,变异系数分别高达41.17%和51.51%。由偏度和峰度统计值看出,F1代全同胞系在可可碱含量、咖啡碱含量及生物碱总量这3个指标上,表现出较好的连续性正态或偏正态分布规律;而在咖啡碱与可可碱含量的比值,虽然也呈连续性分布,但偏度和峰度值较大,正态性较差。
1.1.2 生物碱指标间的相互关系
如表2所示,F1代可可碱和咖啡碱/可可碱这2个指标呈显著负相关(P< 0.01),而咖啡碱(CAF)和生物碱总量(SCT)呈极显著正相关(P< 0.01)。进一步对咖啡碱和生物碱总量这两个高度相关的成对变量建立回归模型,得到显著的一元线性回归模型:YSCT=0.565+1.062XCAF(F=405.34,P< 0.05),决定系数为 0.867,拟合精度高,适合作为茶种金萱(♀)和南昆山毛叶茶(♂)杂交F1代个体早期鉴定咖啡碱和生物碱总量之间的预测模型。

表2 F1代生物碱含量指标的相关性分析表Table 2 Correlation analysis of alkaloid content in F1 generation
F1代可可碱和咖啡碱、咖啡碱和咖啡碱/可可碱指标之间未出现明显的相关关系。这种“不相关”,直接原因由表2可知,可可碱的变异系数大,而咖啡碱的变异系数较小;从基因型推测,本试验杂交亲本中富含可可碱、不含咖啡碱的特殊父本,使得可可碱和咖啡碱从父母本到F1代的遗传,与其它父母本都既含可可碱、又含咖啡碱的茶种杂交组合相比[17],遗传方式发生了本质区别。
1.1.3 基于生物碱特征的聚类分析
基于可可碱和咖啡碱含量的差异,采用离差平方和法对杂交亲子代种群进行聚类,聚类结果如图1所示。父母本及64个F1代全同胞系个体构成的远缘杂交亲子代种群在遗传距离6.0处分为四大类群:其中,第Ⅰ类群包括N09、N44、N11等28个F1代单株;第Ⅱ类群包括母本和N20、N31、N13等15个F1代单株;第Ⅲ类群包括N52、N63、N50等21个F1代单株;父本自聚为第Ⅳ类群。
进一步对Ⅰ~ Ⅳ类群的生物碱特征进行统计分析,结果如表3所示。第Ⅰ类群是F1代最大的类群,其显著特征区别于父母本,含有较高的可可碱和较高的咖啡碱。第Ⅱ类群中的F1代个体生物碱含量特征各方面都与母本相似,母本金萱也可以划分至这个类群;第Ⅱ类群的显著特征与母本相似,含有较低的可可碱和较高的咖啡碱。第Ⅲ类群是F1代第二大类群,其咖啡碱含量和生物碱总量都高于其它类群;第Ⅲ类群的显著特征区别于父母本,含有高咖啡碱。第Ⅳ类群仅含有父本,其特征为:含高可可碱,不含咖啡碱,F1代中未出现类似父本的个体。

图1 基于可可碱和咖啡碱含量的亲子代样本聚类图Figure 1 Cluster map of parent and offspring samples based on theobromine and caffeine content

表3 亲子代生物碱特征聚类统计分析表Table 3 Correlation analysis of alkaloid content index in F1 generation
1.1.4 生物碱相对遗传力和杂种优势
根据相对遗传力理论假设,相对遗传力的取值范围应在0 ~ 1[16]。基因型的杂合性是杂种优势的遗传学基础,杂交种F1表现出来的杂种优势是父母本杂交构成的一个全新的调控系统下基因表达调控的外在表现[18]。
统计父母本相对遗传力(表4),可可碱表现出倾向于母本的负向显性遗传,咖啡碱/可可碱表现出倾向于母本的正向显性遗传,生物碱总量表现出倾向于父本的正向显性遗传。咖啡碱的相对遗传力a1大于1,说明咖啡碱的遗传特性较为复杂,并非严格由基因的加性效应控制,有其它因素(显性效应、上位效应或者环境)的复杂互作对咖啡碱的遗传产生影响。

表4 茶树远缘杂交父母本生物碱性状相对遗传力Table 4 The relative heritability of alkaline form the parents of tea plant distant hybridization
分析F1代杂种优势表现(表5),可可碱表现出介于低亲值和中亲值之间的负向中亲优势,咖啡碱/可可碱和生物碱总量表现出介于中亲值和高亲值之间的正向中亲优势,咖啡碱表现出超过高亲值的正向超亲优势。

表5 茶树远缘杂交F1代生物碱杂种优势表现Table 5 Heterosis performance of alkaloids in F1 generation of tea plants distant crosses
2.2 茶树远缘杂交儿茶素的遗传变异
2.2.1 亲子代儿茶素组成与含量
由表7可见,南昆山毛叶茶(♂)和金萱(♀)及F1代全同胞系之间的儿茶素组成和含量有显著差异。如南昆山毛叶茶(♂)不含GA,而金萱和F1代全同胞系含有GA;南昆山毛叶茶(♂)主要儿茶素的含量依次为:GCG>GC>EGCG>EC>ECG>C>CG;金萱(♀)主要儿茶素的含量依次为:EGCG>C>CG>EC>GC>GCG>ECG;F1代全同胞系主要儿茶素的平均含量依次为:EGCG>GC>C>GCG>ECG>EC>CG;主要酯型儿茶素总量(TEC)、非酯型儿茶素总量(TSC)和儿茶素总量(TC)依次为:金萱(♀)>F1代均值>南昆山毛叶茶(♂);酯型儿茶素与非酯型儿茶素含量的比值(RESC)依次为:金萱(♀)>南昆山毛叶茶(♂)>F1代均值。

表6 亲子代儿茶素组成和含量统计分析表Table 6 Statistical analysis of composition and content of catechins in parent-offspring generations

表7 F1代儿茶素含量指标的相关性分析表Table 7 Correlation analysis of catechin content in F1 generation
F1代儿茶素类指标变异范围为19.28% ~56.04%,平均值为29.54%。变异系数最大的为CG(56.04%),变异系数最小的为EGCG(19.28%)。F1代儿茶素指标均表现出较好的正态或偏正态分布趋势,符合遗传学数量性状的典型特征。
2.2.2 儿茶素指标间的相互关系
如表7所示,GA、7个儿茶素单体指标(C、EC、GC、EGCG、GCG、ECG、CG)和4个儿茶素总量指标(TEC、TSC、RESC、TC)中,大部分成对变量间都有不同程度的相关关系。其中,极显著相关的成对变量有9对,分别是:C与TSC、C与TC、GC与TCS、EGCG与TEC、EGCG与TC、GCG与TEC、ECG与TEC、TEC与TC和TSC与TC,其相关方向均为正向。
综合亲子代优势儿茶素统计结果(2.2.1)和儿茶素指标间的相关性,以EGCG、GCG、GC和C作为自变量进一步建立线性回归模型,精确儿茶素指标间的定量关系,结果如表8所示。EGCG和TEC间存在极显著的一元线性回归方程,决定系数为0.935;EGCG和GCG预测TEC的二元线性回归方程决定系数为0.981;GC和C预测TSC的二元线性回归方程决定系数为0.991;GC、C、EGCG、GCG预测TC的二元、三元、四元线性回归方程的决定系数分别为0.929、0.968和0.980。

表8 F1代儿茶素含量指标的回归模型Table 8 Regression model of catechin content in F1 generation
综合实用性,考察回归模型Y1至Y6:Y1、Y3和Y4拟合精度好、预测效果准确,便捷性高,适合作为早期鉴定茶种金萱(♀)和南昆山毛叶茶(♂)杂交F1代个体儿茶素的指标模型。利用回归模型Y1、Y3和Y4,只需测定EGCG、GC和C这3种儿茶素含量,就能准确有效的预测出F1代个体的酯型儿茶素总量、非酯型儿茶素总量和儿茶素总量。
2.2.3 基于儿茶素指标的综合评价
主成分提取。对GA和7个儿茶素单体(C、EC、GC、EGCG、GCG、ECG、CG) 共 8个指标进行主成分分析,提取得到3个主成分,第一主成分Z1贡献率为56.81%,第二主成分Z2贡献率为14.20%,第三主成分Z3贡献率为10.30%,三个主成分的累计贡献率为81.31%。
因子权重分析。利用最大方差法旋转空间成分矩阵,并进行因子权重分析,进一步解析主成分Z1、Z2、Z3的内涵。由表9和图2可见,第一主成分Z1代表性指标为EGCG、GCG和ECG,可定义为“主要酯型儿茶素”;第二主成分Z2代表性指标为GA、EC和CG,可定义为“微量儿茶素”;第三主成分Z3代表性指标Y为GC和C,可定义为“主要非酯型儿茶素”。

表9 儿茶素主成分因子权重分析表Table 9 Catechin main component factor weight analysis

图2 旋转后空间中的儿茶素主成分矩阵图Figure 2 Principal component matrix of catechins after spatial rotation
F1代儿茶素综合得分排名。根据提取的儿茶素主成分表达式,计算64个F1代全同胞系个体 N01 ~ N64 的儿茶素主成分 Z1、Z2、Z3的分别得分,按照主成分贡献率统计其儿茶素综合得分,得到64个F1代全同胞系儿茶素综合得分排名,列出排名前15的个体如表10所示。

表10 F1代全同胞系个体儿茶素综合得分排名Table 10 F1 progeny of full-sib with catechin comprehensive score ranking
2.2.4 儿茶素相对遗传力和杂种优势
从父母本儿茶素的相对遗传力来看(表11),EGCG、EC、TEC表现出倾向于母本的正向显性遗传,GCG表现出倾向于母本的负向显性遗传,CG表现出倾向于父本的负向显性遗传。
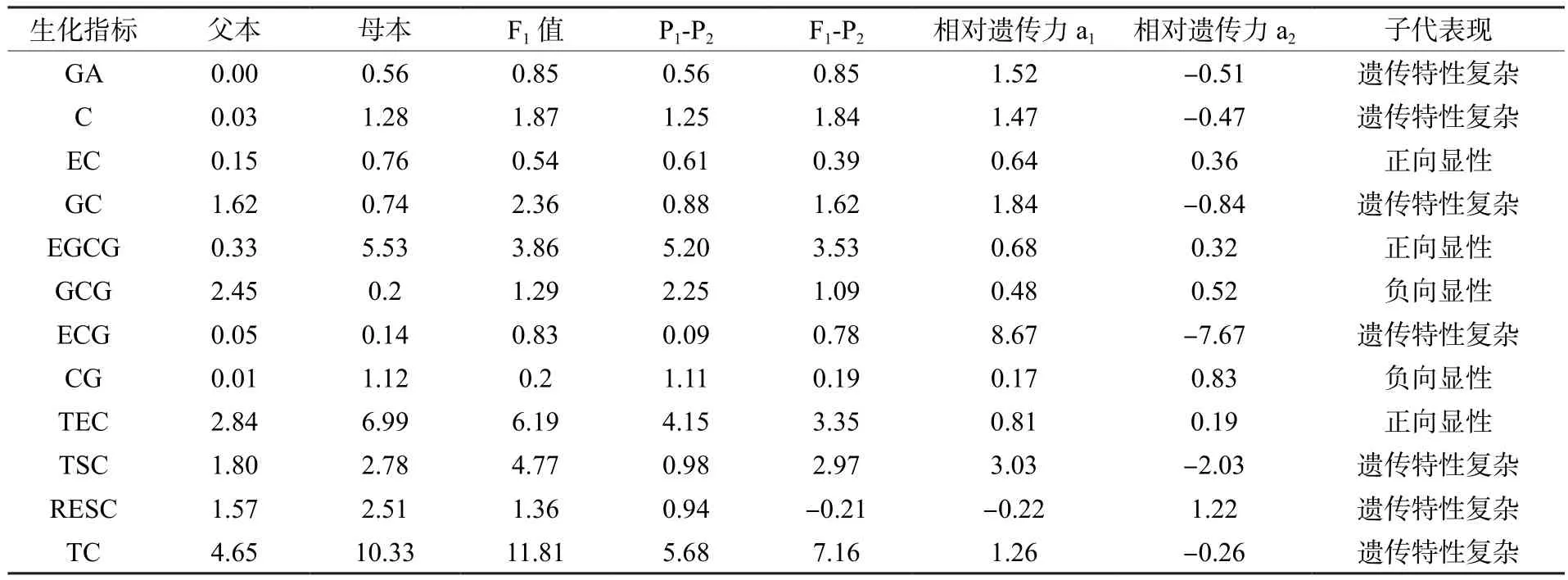
表11 茶树远缘杂交父母本儿茶素性状相对遗传力Table 11 Relative heritability of catechin traits in tea plants distant hybrids
从F1代儿茶素杂种优势表现来看(表12),EGCG、EC和TEC表现出介于中亲值和高亲值之间的正向中亲优势;GCG和CG表现出介于低亲值和中亲值之间的负向中亲优势;GA、C、GC、ECG、TSC、TC表现出超过高亲值的正向超亲优势;RESC表现出低于低亲值的负向超亲优势。
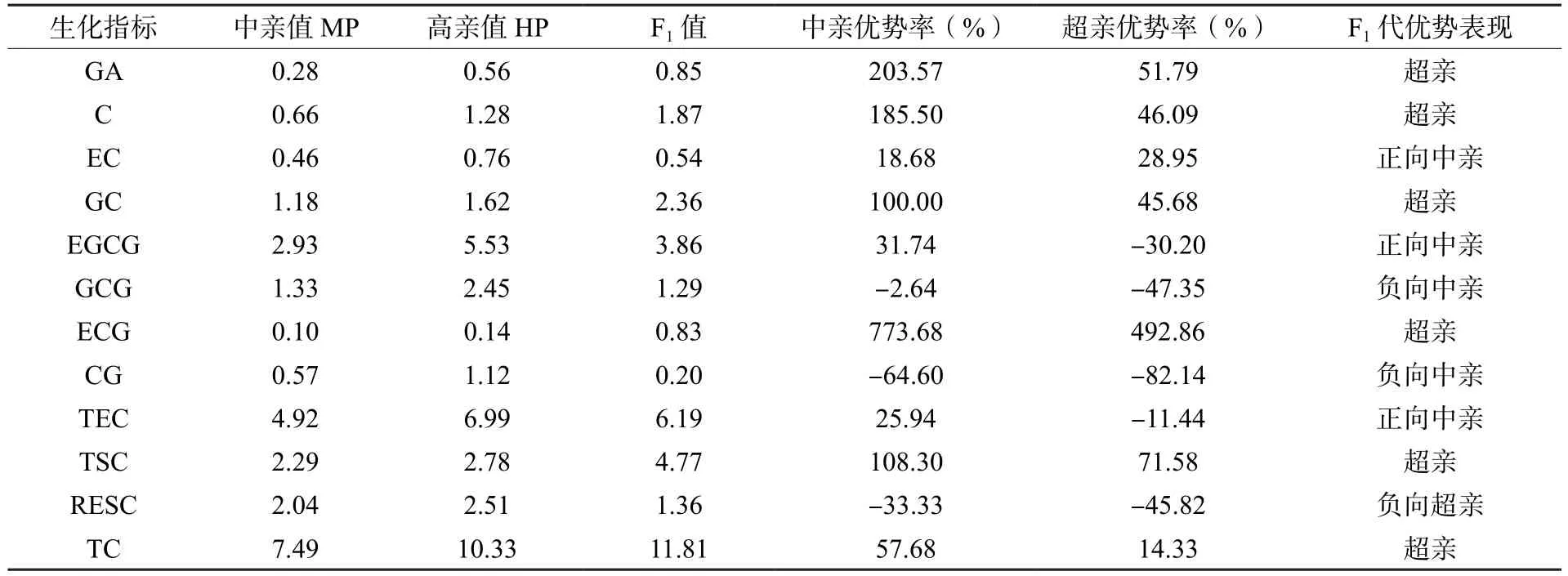
表12 茶树远缘杂交F1代儿茶素杂种优势表现Table 12 Heterosis performance of catechins in F1 generation from tea plants distant crosses
3 讨论
生物碱和儿茶素是茶叶中重要的特异性生化成分。近年来,茶叶中生物碱和儿茶素的合成机理、代谢途径、提取工艺、生理功效等方面的研究,已进行得较为广泛[19-23]。但由于茶树是多年生木本植物,杂交育种年限长,同时茶树杂交育种还存在杂交结实率低、自交不亲和等关键问题[24],导致成规模的杂交种群构建难度大,成体系的遗传机理研究进展慢。茶树生物碱和儿茶素在杂交代际间遗传规律的研究成果仍然较为有限。
涂良剑等[25]研究3个高EGCG茶树品系和1个推广品种相互杂交的F1代,发现F1代咖啡碱和儿茶素总量的遗传不是倾向于亲本的某一方,而是因不同杂交组合中亲本的遗传力而定,而GA未呈现明显的规律性;张金霞等[26]研究黄山群体种自然杂交后代,发现酯型儿茶素的变异系数高于非酯型儿茶素,变异系数最大的是C的含量;李家华等[27]研究不同品种杂交亲本和21个F1代单株,发现茶多酚在后代中的遗传难以用总含量作为依据判断,而经过提取儿茶素主成分和聚类分析后,茶多酚表型以3∶10的分离比例被分成类似于母本的类型和类似父本类型;中国农科院茶叶研究所[17,28-29]依托杂交亲子代种群,鉴定出25个茶树儿茶素含量相关的QTL,10个与可可碱和咖啡碱含量相关的QTL,以及跟儿茶素总含量相关的AA、AG和GG三种特定基因型。以不同茶树杂交群体材料对生物碱和儿茶素遗传变异的研究,结果之间有一定的差异性,主要是由以下4点原因造成:一是父母本的表型性状有所不同,即双亲生物碱和儿茶素的分布模式、含量特点不同;二是父母本的基因型有所不同(根本原因):双亲遗传距离不同,导致杂交表观遗传方式和杂种优势表现强弱不同[30];三是种群的规模大小不同;四是选取的具体指标和分析方法不同。
经济性状表现好或特殊、稀有的种质资源都属于优异种质资源,可以直接用于茶树新品种选育或者间接为茶树遗传改良提供优良基因来源[31]。本文茶树远缘杂交种群的双亲均为华南地区优异茶树种质资源,母本金萱经济性状好,父本“可可茶”具有无咖啡因特色。通过对64个远缘杂交F1代个体的分析筛选,得到低可可碱材料1份(0.16%)、高可可碱材料1份(1.70%)、低咖啡碱材料1份(2.48%)和高咖啡碱材料1份(5.84%),可用于进一步分子生物学研究,揭示咖啡碱和可可碱的遗传机理;也可用于直接或间接地选育生物碱特色显著的茶树新品种。儿茶素对成茶品质具有一定预测性[32],F1代筛选出儿茶素总量大于15%的材料 8 份(N46、N49、N52、N54、N56、N57、N63、N64),酯型儿茶素与非酯型儿茶素比值大于2.00 的材料2份(N16、N20),后续可对其成茶适制性进行鉴定,推动茶树杂交新品种选育的发展。
猜你喜欢
科学(2023年2期)2023-05-30 08:57:46
环境科学与管理(2023年3期)2023-04-29 03:02:27
安徽农业大学学报(2022年2期)2022-11-21 06:51:15
麦类作物学报(2022年5期)2022-05-20 09:43:24
食品安全导刊(2021年20期)2021-08-30 06:39:40
西南农业学报(2018年5期)2018-06-05 07:08:22
茶叶学报(2017年3期)2017-11-23 07:56:27
上海农业学报(2017年3期)2017-04-10 12:39:24
家庭科学·新健康(2016年5期)2016-05-12 00:05:19
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28 12:22:07
