
木通科花瓣的发育、微形态及超微结构研究
2022-11-09 10:42张小卉
陕西师范大学学报(自然科学版) 2022年5期
刘 男,苏 珊,张小卉
(陕西师范大学 生命科学学院,药用资源与天然药物化学教育部重点实验室,陕西 西安 710119)
花是被子植物重要的有性繁殖器官,在被子植物进化过程中扮演着重要角色。花瓣作为花的第二轮器官,可能来源于苞片(木兰目(Magnoliales)和八角目(Illiciales))或雄蕊退化(毛茛目(Ranunculales))[1],其发育过程中包含着多种系统演化信息[2]。毛茛目的成熟花瓣形态多样,在颜色、大小、形状、结构、蜜腺、蜜汁成分、表皮细胞类型等方面表现出丰富的多样性[1,3-9];其中,花瓣上的蜜腺结构可以产生蜜汁来吸引传粉者,具有重要的生物学意义[10-14]。
木通科(Lardizabalaceae)作为毛茛目的核心成员之一,其花瓣短小、绿色,被称为蜜腺叶[15-16]。该科共有9个属,约50个种,主要分布在亚洲东南部以及南美洲的智利[16-19]。我国木通科的有花瓣类群隶属于3个分支,分别为大血藤属(SargentodoxaRehder & E.H.Wilson)的第一分支、串果藤属(SinofranchetiaHemsl.)的第二分支以及由野木瓜属(StauntoniaDC.)有瓣亚属和八月瓜属(HolboelliaHook.)组成的第三分支。目前,有关木通科的研究主要集中在系统发育、花器官发生发育、胚胎学、花瓣发育基因及种质资源等方面[20-26],有关木通科花瓣的发育特征、蜜腺结构、花瓣在科内是否存在分化等信息匮乏,阻碍了对该类群繁殖生物学及毛茛目花瓣多样性的进一步认知[22]。
针对上述问题,本研究拟对我国特有或东南亚特有的木通科大血藤属、串果藤属及牛姆瓜属开展以下研究:(1)观察花瓣的形态发育特征;(2)阐明花瓣蜜腺组织的位置及泌蜜方式;(3)比较不同属花瓣的微形态及超微结构特征,分析其祖先花瓣性状,推断花瓣在繁育过程中的功能。基于以上研究,探讨木通科花瓣的多样性特征,为深入了解毛茛目花瓣多样性及演化问题提供一定的科学依据。
1 材料与方法
1.1 植物材料
大血藤(Sargentodoxacuneata(Oliv.) Rehd. Et Wils.)、串果藤(SinofranchetiachinensisHemsl.)和牛姆瓜(HolboelliagrandifloraReaub.)3个属的植物均于2014年至2016年采自秦岭区域(大血藤和牛姆瓜的雌花、雄花、花蕾采自陕西佛坪,串果藤的雌花、雄花、花蕾采自陕西眉县)。采集的所有花芽和成熟花均固定在2%戊二醛(glutaraldehyde)的固定液中。
1.2 扫描电子显微镜法
使用扫描电子显微镜观察花瓣的形态发育和微形态材料,将不同发育阶段的花芽及成熟花在解剖镜下剥离,分别经梯度乙醇逐级脱水、乙酸异戊酯置换后,置于二氧化碳临界点干燥仪(K850)中进行干燥,再经黏台、喷金镀膜后,在扫描电子显微镜下(日立HITACHI S-3400N)观察、拍照。
1.3 透明法
将花瓣维管组织分化材料置于5%的NaOH溶液中,60 ℃温箱过夜,使材料透明;然后使用0.1%的甲苯胺蓝染色,在倒置荧光显微镜(Leica DM5000B,DFC490成像系统)下观察、拍照。
1.4 荧光染色法
花瓣的蜜腺组织放置在0.05%的苯胺蓝溶液中,室温染色12 h,激发光波长为500~580 nm,显微镜(Leica DFC490)下观察、拍照。
1.5 半薄切片法
对花瓣结构的固定材料依次进行磷酸缓冲液清洗、1%锇酸溶液(OsO4)固定、乙醇系列逐级脱水、环氧丙烷置换、Epon812树脂包埋、60 ℃聚合48 h、切片机(Leica EM U6型)切片(厚度2 μm)和0.05%甲苯胺蓝溶液染色,封固后在倒置荧光成像系统(Leica CTR5000,DFC490)下观察、拍照。
1.6 透射电子显微镜法
成熟花瓣蜜腺细胞内部结构的观察采用超薄切片法,材料的包埋方法同半薄切片法(见1.5),切片厚度为60 nm。将材料置于铜网碳膜上干燥后,依次用2%醋酸铀溶液(uranyl acetate)染色30 min、2%柠檬酸铅溶液(lead citrate)染色10 min,清洗干燥后,使用透射电子显微镜(日立HT-77000)在电压60 kV条件下观察、照相。
1.7 祖先性状演化分析
从NCBI数据库获得木通科10个种的matK、rbcL和trnL-F序列,参考已有研究,选取星叶草科(Circaeasteraceae)独叶草(KingdoniaunifloraBalf.f. & W.W.Sm.)、毛茛科(Ranunculaceae)黄连(CoptischinensisFranch.)、小檗科(Berberidaceae)南天竹(NandinadomesticaThunb.)、防己科(Menispermaceae)中华青牛胆(Tinosporasinensis(Lour.) Merr.)为外类群[27],在Genious软件中进行序列比对,使用Beast v1.4.0构建系统发育树,将结果导入Mesquite中,使用MK模型进行祖先性状重建。
2 结果与分析
2.1 花的形态
由花形态观察结果可以看出(图1),木通科植物的花小、单性、稀两性、雌雄同序或异序,常成总状花序,花器官通常为三基数。其中,大血藤的花具有萼片6枚,黄绿色,呈花瓣状(图1a、1b、1c);花瓣(蜜腺叶)6枚,黄白色,较小,呈卵形(图1d)。大血藤雄花具有雄蕊6枚,退化心皮无或少数(图1a);两性花具有雄蕊6枚,雌蕊10~50枚(图1b),雌、雄蕊成熟期不同;雌花具有雌蕊约90枚,螺旋状生于突起的花托上,花瓣状退化雄蕊6~11枚(图1c)。
串果藤的花具有萼片6枚,绿色,外缘紫色,呈花瓣状;花瓣6枚,较小,呈倒匙形,顶端二裂,黄绿色,发育后期边缘为紫色(图1e、1f)。串果藤雄花具有雄蕊6枚,退化心皮3枚(图1e);雌花具有退化雄蕊6枚,心皮3枚(图1f)。牛姆瓜花具有6枚肉质、花瓣状的萼片(图1g、1h),绿白色或紫色;花瓣6枚,微小,近圆形(图1i)。牛姆瓜雄花中雄蕊6枚,退化心皮3枚;雌花中退化雄蕊6枚,心皮3枚。
2.2 花瓣的个体发育及组织分化
在3个属的植物花瓣原基发生后,依据发育过程中关键性状出现的特征,可将花瓣形态发育过程划分为4个阶段:阶段1,原基发生,花瓣原基为新月形;阶段2,暂停发育(延迟发育);阶段3,花瓣延展,基部形成柄,发育为具有短尖的宽卵形或顶端二裂的倒匙形;阶段4,花瓣成熟。雌花和雄花的花瓣发生发育特征基本相同(图2、3)。
在大血藤雄花的幼小花蕾中,当雄蕊分化出花粉囊后,在发育阶段3花瓣增大为宽卵形,中央纵向伸长,顶端出现脊状突起,基部形成明显的柄部(图2a、2b、2c),此时维管束单一无分支(图3a)。大血藤成熟花瓣为近菱形或倒卵形,高约1.2 mm,有的顶端出现二裂或多裂现象,维管束在中部形成2~3个分支(图3b)。大血藤雌花和两性花的花瓣与雄花花瓣发育特征相似(图2d、2e、2f),分泌组织位于顶部,由分泌表皮细胞和位于表皮下的约5层分泌薄壁细胞构成(图3c、3d、3e)。
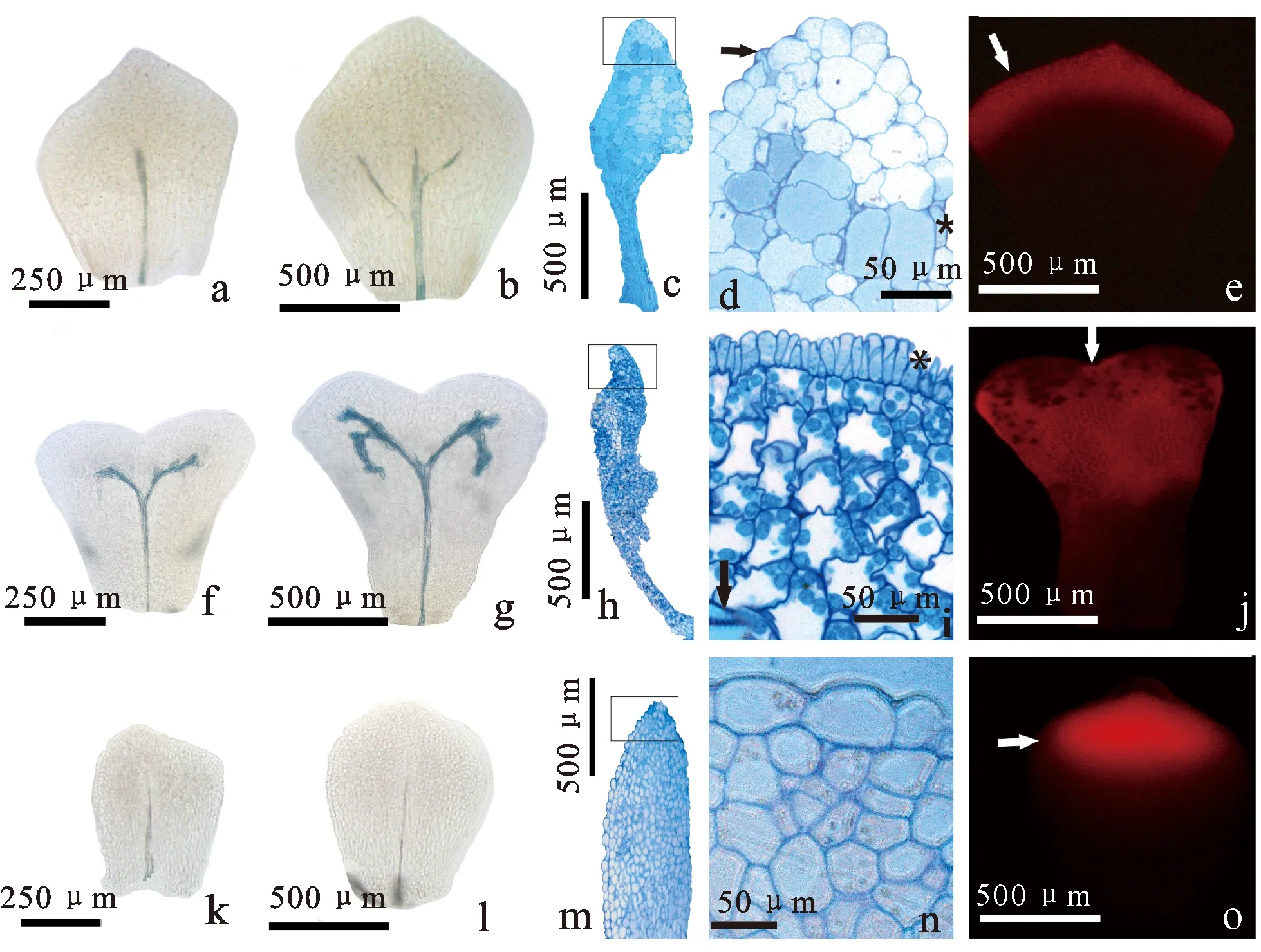
a、b. 大血藤不同发育阶段的花瓣,标示维管组织的形成过程;c、d. 大血藤花瓣的结构,图d为图c的上部放大,指示蜜腺组织结构(箭头示分泌表皮,星号示变态气孔);e. 大血藤成熟花瓣,指示分化的蜜腺组织(箭头所示红色区域);f、g. 串果藤花瓣的维管组织分化,指示维管束末端多次分支;h、i. 串果藤花瓣结构,图i为图h的分泌区放大(箭头示维管束,星号示分泌表皮);j. 串果藤蜜腺组织分布区域(箭头所示);k、l. 牛姆瓜花瓣的维管组织分化;m、n. 牛姆瓜花瓣结构,图n为图m的分泌区放大;o. 牛姆瓜蜜腺组织分布区域。
串果藤雄花花瓣在经历延迟发育后(图2g),在阶段3发育为倒梯形,随后在顶端中央部分出现浅凹(图2h)和二裂(图2i),并在基部形成明显的柄部(图2j),维管束分为2个简单的分支(图3f)。开花时,花瓣为顶端二裂的倒匙形,高约1.5 mm(图2k),维管束深入瓣部且再次分支(图3g)。串果藤雌花花瓣的发育特征与雄花相似(图2l),成熟花瓣的分泌组织位于花瓣的瓣部,约占花瓣的2/3(图3h、3j),由指状分泌表皮细胞、5~6层富含淀粉粒的分泌薄壁细胞以及维管束组成(图3i)。
牛姆瓜的花瓣发育特征与大血藤花瓣基本相似,开花时雄花花瓣为微小卵形,高约500 μm(图2m、2n),具有1条不分支的维管束(图3k、3l),分泌组织位于花瓣顶端,由表皮细胞及约6层分泌薄壁细胞组成(图3m、3n、3o)。雌花的花瓣为约250 μm高的三角形(图2o),未见维管束和分泌组织。
2.3 花瓣的表皮微形态
在木通科3个属的植物花瓣中共发现有5种类型的表皮细胞,其中3种表皮细胞具有分泌功能(图4)。表皮细胞类型1:细胞形状为不规则多边形,长约40 μm,宽约12 μm,具有分泌功能,细胞表面具有丝状分泌物(图4b)。表皮细胞类型2:细胞形状为不规则多边形,长约38 μm,宽约13 μm,不具有分泌功能,细胞表面光滑(图4c)。表皮细胞类型3:
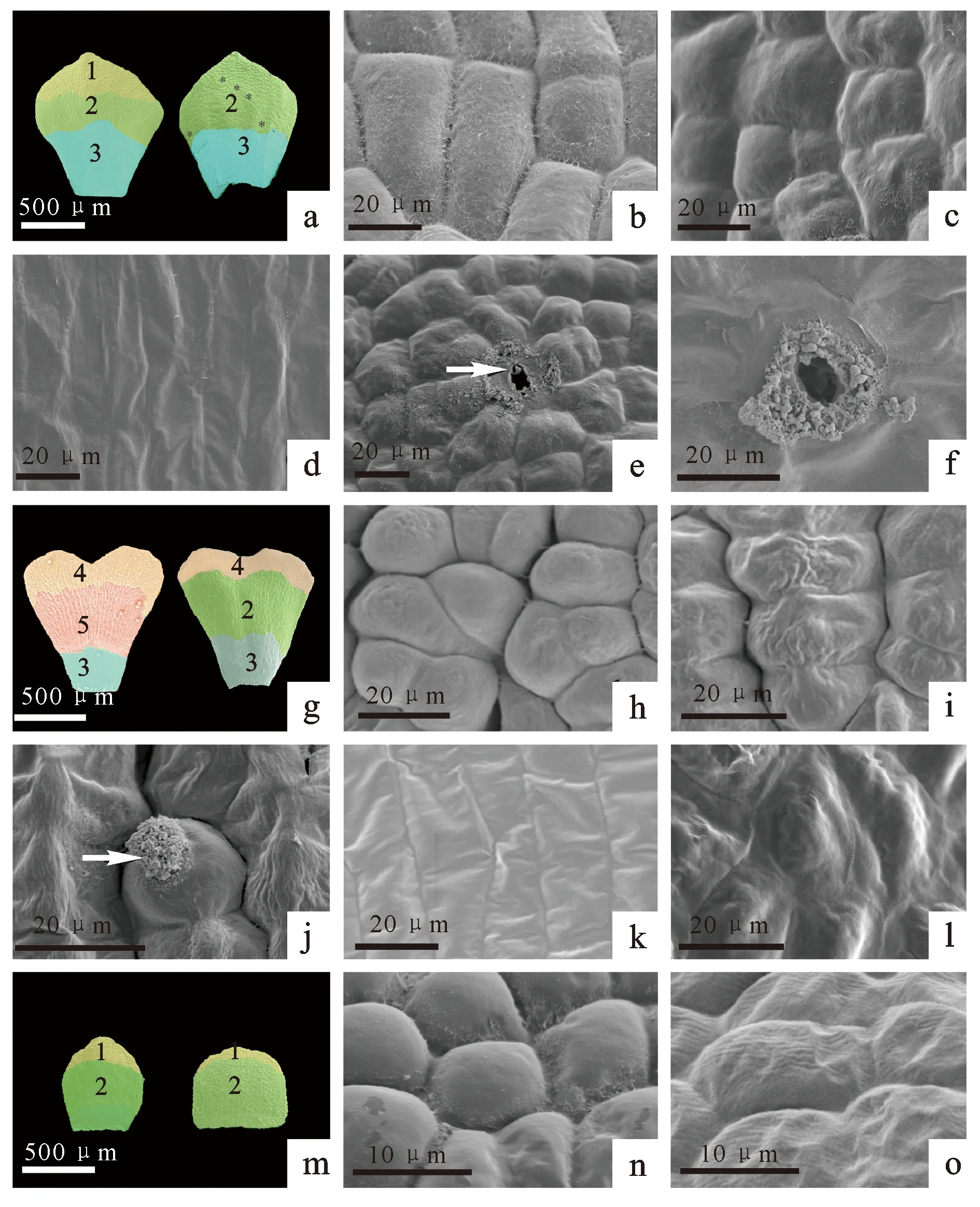
a. 大血藤雄花花瓣(左图为近轴面,右图为远轴面);b、c、d. 大血藤雄花花瓣表皮细胞类型,b、c、d分别是图a中细胞类型1、2、3的放大;e. 图a中星号所示区域放大,指示花瓣远轴端的变态气孔器;f.图e的放大,箭头示分泌物;g. 串果藤雄花花瓣(左图为近轴面,右图为远轴面);h. 串果藤表皮细胞类型4,为近圆形分泌细胞;i. 串果藤表皮细胞类型5,为乳突状分泌细胞;j. 串果藤表皮细胞类型5发育后期,箭头示分泌物从破裂的细胞中释放;k. 串果藤表皮细胞类型3;l. 串果藤表皮细胞类型2,不规则多边形非分泌细胞;m. 牛姆瓜雄花花瓣;n. 牛姆瓜表皮细胞类型1;o. 牛姆瓜表皮细胞类型2。图中数字代表细胞类型,不同颜色区域所示细胞类型不同。
细胞呈长矩形,长约60 μm,宽约18 μm,不具有分泌功能,细胞表面光滑或存在横向条纹(图4d)。表皮细胞类型4:细胞形状为近圆形,横截面直径约为10 μm,具有分泌功能,表面粗糙,细胞与细胞间存在丝状分泌物(图4h)。表皮细胞类型5:细胞形状为乳突状,长约8 μm,宽约17 μm,具有分泌功能,细胞表面有条纹,中央有乳状突起(图4i)。
大血藤成熟花瓣的表皮细胞共有3种类型。表皮细胞类型1位于近轴面顶端(图4a、4b),类型2位于近轴面中部和远轴端上部(图4a、4c),类型3位于花瓣基部(图4a、4d)。在花瓣远轴面,分布着少量的变态气孔器(图4a),分泌的小颗粒堆积在气孔周围(图4e、4f)。
串果藤成熟花瓣的表皮细胞共有4种类型。其中,类型4近圆形分泌细胞位于花瓣近轴面和远轴面的顶端(图4g、4h);类型5乳突状分泌细胞位于花瓣近轴面的中部,盛花期时,细胞壁破裂,分泌物释放(图4i、4j);类型2和类型3表皮细胞分别位于花瓣的柄部(图4g、4k)和远轴端的中部(图4l)。
牛姆瓜成熟花瓣的表皮细胞仅有2种类型。类型1不规则多边形分泌细胞位于雄花花瓣近轴面和远轴面的顶端(图4m、4n),类型2多边形细胞位于花瓣的中部和基部(图4m、4o)。雌花花瓣仅具有类型2表皮细胞。
2.4 花瓣的超微结构
木通科3个属花瓣分泌表皮细胞的外切向壁均具有发达的微通道,细胞质浓厚,含有丰富的淀粉粒、线粒体和粗面内质网,薄壁细胞间存在大量的胞间连丝(图5、6)。
在大血藤盛花期,花瓣分泌表皮细胞的细胞核较大,内切向壁具有明显的内突生长(图5a、5b、5c、5d),表皮下的薄壁细胞呈不规则多边形,细胞壁迂回折叠,具有丰富的细胞器,粗面内质网较多(图5e、5f、5g)。细胞间隙发达,蜜汁贮存其内(图5h)。串果藤花瓣的分泌表皮细胞形态较为规则,细胞中淀粉粒丰富(图6a、6b、6c、6d),表皮细胞之间、表皮细胞与表皮下的分泌薄壁细胞之间的细胞壁上具有明显的内突生长(图6e),筛管与筛管间可见发达的筛板(图6f、6g)。牛姆瓜雄花花瓣的分泌表皮细胞壁外侧可见丝状分泌物(图6h),分泌表皮细胞与分泌薄壁细胞之间具有发达的胞间隙,储存有蜜汁(图6i)。
2.5 木通科花瓣祖先性状重建
木通科祖先性状重建分析表明(图7),木通科祖先具有花瓣(52.5%);基部的大血藤属和串果藤属祖先节点性状显示花瓣存在(分别是51.2%和56.6%);次基部的猫儿屎属花瓣在演化过程中发生丢失;在由八月瓜属等6个属组成的分支中,花瓣状态较为复杂,其祖先节点的状态为花瓣存在(58.0%)。
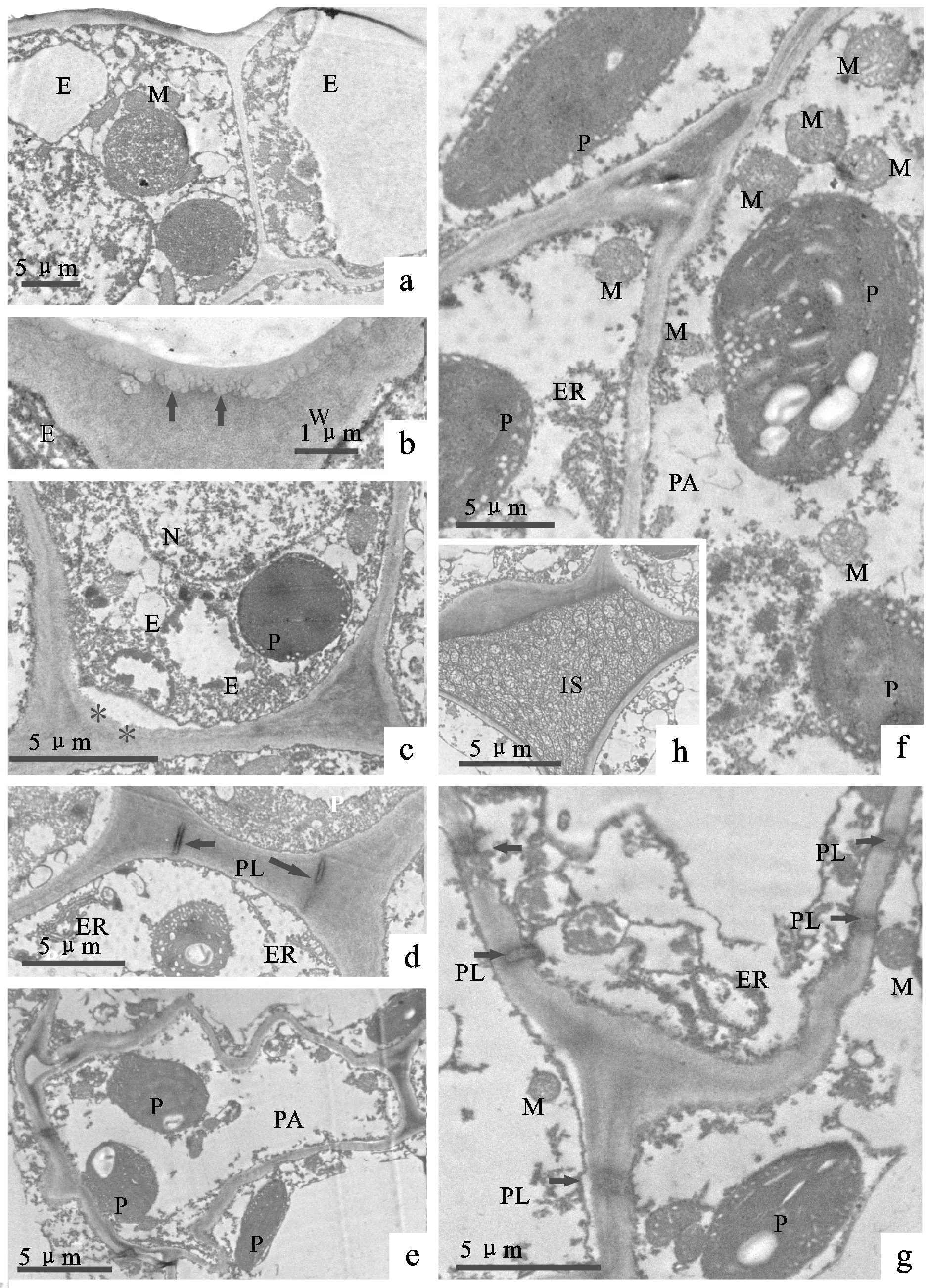
a. 分泌表皮细胞,展示发达的线粒体;b. 表皮细胞外切向壁放大,箭头指示丰富的微通道;c. 表皮细胞内切向壁,星号指示内突生长;d. 表皮细胞与表皮下的分泌薄壁细胞,箭头指示胞间连丝;e、f、g. 分泌薄壁细胞;h. 分泌薄壁细胞间发达的胞间隙。E. 表皮细胞; ER. 内质网; IS. 细胞间隙; M. 线粒体; N. 细胞核; P. 质体; PA. 薄壁组织; PL. 胞间连丝; W. 细胞壁。
3 讨论
3.1 木通科花瓣的形态发育特征及性状分析
木通科为毛茛目的核心类群[20],而毛茛目植物花瓣(蜜腺叶)被认为是从典型的不育雄蕊进化而来[1, 26, 28]。毛茛目(除领春木科无花瓣外)各科花瓣由于早期发育特征不同,最终形成的花瓣形式多样,其花瓣在发育过程中会出现凸起或凹陷,成熟花瓣表现出管状、囊状、二唇状、高脚杯状等立体结构[9]。在小檗科中,花瓣早期边缘加厚,中央会形成浅凹或在基部产生一对突起,最终形成卵形、漏斗状或扇形的成熟花瓣;防己科的花瓣在发育后期,顶端会产生对折、二裂或卷折[28-29]。木通科为单系支[27], 在木通科有花瓣的类群中, 分布在南美洲的拉氏藤属(LardizabalaRuiz & Pav.)和勃奎拉属(BoquilaDecne.)仍鲜有报道,但本研究结果表明,大血藤属、串果藤属、牛姆瓜属的花瓣早期发育特征较为相似,其花瓣发育均经历了延迟,并在发育阶段3后产生分歧。串果藤的花瓣横向生长较快,顶端产生二裂;大血藤和牛姆瓜的花瓣由宽卵形逐渐发育为柄部分化不明显、顶端尖的倒卵形(大血藤)或卵圆形(牛姆瓜)。木通科大多数类群在演化过程中保留了祖先具有花瓣的性状特征,但在少数属中出现了花瓣丢失现象。

a、b、c、d.串果藤分泌表皮细胞,a展示淀粉粒,b展示外切向壁(箭头示丰富的微通道),c、d分别示内切向壁的内突生长(星号所示)和胞间连丝(箭头指示胞间连丝);e. 串果藤分泌薄壁细胞,展示丰富的线粒体(M);f. 串果藤导管分子与筛管分子;g. 串果藤筛管分子,展示筛板; h. 牛姆瓜分泌表皮细胞,箭头指示外切向壁上的微通道;i. 牛姆瓜分泌薄壁细胞,展示淀粉粒和内质网。E. 表皮细胞; ER. 内质网; M. 线粒体; N. 细胞核; P. 质体; PA. 薄壁组织; PL. 胞间连丝; Se. 筛质元素; V. 容器元素; W. 细胞壁。
3.2 木通科花瓣蜜腺组织的结构及泌蜜方式
毛茛目的花瓣既有简单的片状结构,也有复杂的立体结构,其蜜腺在花瓣上的位置和结构也存在不同[1-2, 12, 30-32],有的类群蜜腺隐藏在花瓣基部的鳞片、短囊或距内(如毛茛属(RanunculusL.)、耧斗菜属(AquilegiaL.)、淫羊藿属(EpimediumL.))[7,33-34],而有的类群蜜腺暴露在简单微小的花瓣瓣部(如红毛七属(CaulophyllumMichx.)、蝙蝠葛属(MenispermumL.))[35-36]。维管组织的参与程度与蜜汁分泌量密切相关[12],毛茛科中不同属植物的蜜腺组织结构不同,如飞燕草属(ConsolidaGray)的蜜距中既有木质部又有韧皮部,而耧斗菜属和乌头属(AconitumL.)的蜜距中仅有韧皮部伸入到分泌薄壁细胞之间[32]。蜜汁通常通过微通道或表皮细胞外切向壁破裂释放到细胞外[33-34]。
在本研究中,木通科3个属的花瓣为简单的片状,蜜腺组织位于花瓣瓣片顶端,由分泌表皮细胞和5~6层分泌薄壁细胞组成,偶有维管束参与分泌活动(串果藤属)。蜜汁通过微通道、分泌表皮细胞壁破裂等方式泌出,与毛茛科的蜜汁排出方式相符[33-34];此外,大血藤花瓣还有变态的气孔作为泌蜜通道,这种泌出方式在被子植物中较为普遍[37]。
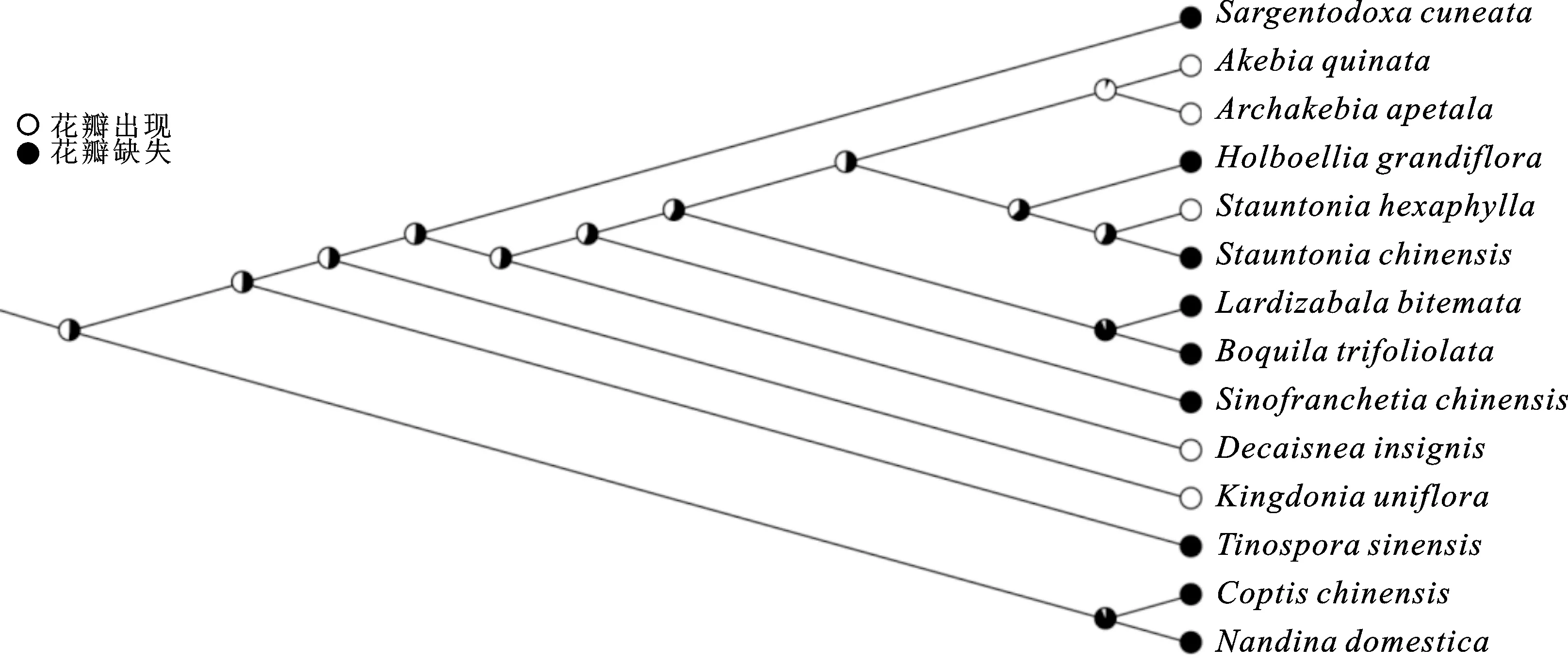
图7 MK模型重建木通科祖先花瓣状态Fig.7 Reconstruction of ancestral petal state using MK model
当前,有关木通科的传粉生物学研究较少,有学者曾根据大血藤属的绿色花推断其可能为风媒传粉[22],但本研究表明该科花瓣具有分泌功能,且在大血藤的两性花中观察到了访花昆虫,因此该科植物的传粉方式和繁殖策略有待进一步深入研究。
4 结论
通过对木通科3个属花瓣的个体发育特征进行比较,发现在木通科有花瓣类群中,不同分支上的花瓣在发育时期顶端出现了二裂、变尖、基部伸长等不同变化,并最终造成花瓣形成了顶端二裂的倒匙形、倒卵形、宽卵形等形态。木通科3个属的成熟花瓣均具有分泌功能(除牛姆瓜的雌花花瓣),蜜腺组织位于花瓣顶端,分泌组织由分泌表皮细胞和分泌薄壁细胞构成,蜜汁多通过表皮细胞壁微通道、变态气孔或表皮细胞壁破裂的方式向外界释放。木通科祖先性状重建分析结果表明,其祖先具有花瓣特征(52.5%),在演化过程中大多数类群保留了祖先状态,即存在花瓣,少数类群丢失了花瓣特征。
猜你喜欢
今日农业(2022年3期)2022-06-05
建材发展导向(2021年14期)2021-08-23
中华养生保健(2020年10期)2021-01-18
中国生殖健康(2020年7期)2021-01-18
中国生殖健康(2020年6期)2020-02-01
祝您健康·文摘版(2019年4期)2019-06-11
新作文·小学高年级版(2017年10期)2017-11-13
学苑创造·A版(2016年10期)2016-11-19
饮食科学(2016年3期)2016-07-04
奥秘(2015年4期)2015-09-10
