SOX2-OT/SOX2 axis regulates lung cancer H520 cell migration via Gli1-mediated epithelial-mesenchymal transition
2022-11-03 12:34DONGHongliangZENGLiliWUYanMIAOShuangNINaLIUNaiguoCHENWeiweiDUJing
南方医科大学学报 2022年10期
DONG Hongliang ,ZENG Lili ,WU Yan ,MIAO Shuang ,NI Na ,LIU Naiguo ,CHEN Weiwei,DU Jing
1Medical Research Center,2Department of Oral and Maxillofacial Surgery,Binzhou Medical University Hospital,Binzhou 256600,China;3School of Stomatology,Binzhou Medical University,Yantai 264003,China
Abstract: Objective To explore the regulatory role of SOX2-OT in migration of lung squamous cell carcinoma H520 cells and the underlying mechanisms.Methods Wound-healing and Transwell migration assays were performed to examine the changes in migration and invasion capacity of lung squamous cell line H520,which expressed higher levels of SOX2-OT than other lung cancer cell lines,following RNA interference-mediated SOX2-OT knockdown.The transcription levels of epithelial-mesenchymal transition (EMT)-related components was detected by qRT-PCR and immunoblotting.Gli1 gain-of-function analysis was performed in H520 cells with SOX2-OT knockdown and the changes in EMT phenotype of the cells were examined.miR-200c mimic and inhibitor were used to analyze the mechanism by which SOX2-OT positively regulates Gli1 and the mediating role of SOX2.Results SOX2-OT knockdown significantly lowered the invasiveness and migration capacity of H520 cells and caused changes in EMT phenotype of the cells.Overexpression of Gli1,which was positively regulated by SOX2-OT,reversed the inhibitory effect of SOX2-OT knockdown on migration of H520 cells.Transfection of the cells with miR-200c inhibitor effectively reversed SOX2-OT knockdown-induced down-regulation of SOX2.Conclusion The SOX2-OT/SOX2 axis positively regulates migration of lung squamous H520 cells via Gli1-mediated EMT.
Keywords:SOX2-OT;lung squamous cell carcinoma;Gli1;SOX2;epithelial-mesenchymal transition
INTRODUCTION
According to the GLOBOCAN estimates,lung cancer is the second most commonly diagnosed cancer worldwide and the leading cause of cancer death[1].Non-small cell lung cancer (NSCLC),including lung squamous cell carcinoma (LUSC) and lung adenocarcinoma (LAD),constitutes nearly 85% of the total lung cancer cases.Metastasis of LUSC especially to the brain is associated with a high mortality rate,and about half of lung cancer patients have brain metastases at the time of diagnosis[2].The exploration of molecular diagnostic markers and potential therapeutic targets for LUSC metastasis is therefore of vital importance.
Years passed. I was twenty-six, and my friends were getting nervous about my prospects4. They kept lining5 up blind dates for me. Many of these dates were fiascoes, and they were interfering6 with my social life. So I made up a few rules about blind dates:
Thedevelopmentofhighthroughputgene-sequencing technology and bioinformatics allowed the discovery of numerous long non-coding RNAs (lncRNAs) that are closely linked to tumor metastasis.These lncRNAs are capable of regulating gene expressions at the transcription,post-transcription and translation levels.We previously analyzed the role of SRY-related HMG box-2(SOX2),a stem cell-specific transcription factor,in tumorigenesis and stemness of lung cancer cells[3,4].Its host gene SOX2-OT is located on chromosome 3q26.33 in human,and its intron contains SOX2 that regulates the propagation and differentiation of cancer stem cells[5,6].The abnormal expression of SOX2-OT and SOX2 caused by frequent amplification of human chromosome 3q26 is a genetic feature of LUSC and occurs in more than 70%of the patients[7,8].Big data analysis showed that SOX2-OT expression is upregulated in lung cancer,breast cancer,and various other malignancies,and its transcription level is closely related with tumor size,early invasion,and metastasis,and is negatively correlated with the patients'overall and disease-free survival[9-11].
Cancer cell invasion and distant metastasis are complex biological processes closely related with epithelial-mesenchymal transition (EMT)[12].As the initial step for tumor metastasis,EMT is regulated by multiple cellular remodeling transcription factors such as vimentin,Snail,and Twist[13].The glioma-associated oncogene Gli1 encodes the most critical transcription factor downstream of the Hedgehog signaling pathway to regulate the expressions of multiple stem cell markers and the genes related with drug resistance and metastasis of tumors[14].We previously demonstrated that the expression of Gli1 was negatively correlated with the survival outcomes of lung cancer patients,and inhibition of Gli1 enhanced the sensitivity of lung cancer cells to chemotherapeutic agents[3,4,15].In addition,Gli1 was reported to be specifically and highly expressed in LUSC tissues,and blocking the Hedgehog signaling pathway effectively inhibited the proliferation of LUSC cells[16].However,whether SOX2-OT modulates LUSC cell migration through Gli1-mediated EMT remains unclear.In this study,we aimed to further explore the pivotal role of SOX2-OT in regulating migration of LUSC H520 cells and the underlying mechanism.
METHODS
Cell culture
We detected the expression levels of EMT-related genes(E-cadherin,vimentin,Snail,and Twist) by qRT-PCR and immunoblotting in H520 cells with SOX2-OT knockdown,and the results showed that the expression levels of vimentin and Snail were decreased and that of E-cadherin was increased significantly (Fig.3A-C),but the expression of Twist underwent no significant changes.As previous studies suggest that the key transcription factor Gli1 of the Hedgehog cascade plays a pivotal role in EMT and cellular invasion[17,18],we further explored the effect of SOX2-OT knockdown on Gli1 expression.Our data showed that compared with the control shRNA,shRNA #4 and #5 targeting SOX2-OT reduced Gli1 expression by 67% and 58% at the mRNA level and by 59% and 23% at the protein level,respectively(Fig.4A-C).
Plasmid construction and transfection
The plasmids pcDNA3.1-Gli1 and pcDNA3.1-Sox2 were constructed using the Clone Express MultiS One Step Cloning kit of Vazyme (Nanjing,China) and verified by sequencing.Short hairpin RNA (shRNA) plasmids against SOX2-OT were purchased from Genechem(Shanghai,China).The mimic and inhibitor for has-miR-200c-3p were purchased from Ribobio(Guangzhou,China).The plasmid vectors were extracted by DNA Midiprep kit (Qiagen,Germany) and delivered into cells using Lipofectamine 3000 Mixture (Thermo Fisher Scientific,USA) in a 6-or 12-well cell culture plate following the manufacturer's instructions.At 48 h after transfection,the stably transfected cells were selected by treatment with 400 μg/mL G418 or 1 μg/mL puromycin.
RNA purification and qRT-PCR assays
Wound-healing assay was conducted as previously described[3].Briefly,the cells in exponential growth were seeded into a 12-well plate,and when the cells grew to 80%confluence,the cell monolayer was scratched with a sterile pipette tip.The plate was repeatedly washed with PBS to remove cell debris and incubated with serum-free medium for 24 h.Wound healing was photographed using an inverted fluorescence microscope and the rate of closure was assessed.

Tab.1 Sequences of the primers for qRT-PCR
Immunoblotting
The cells were collected and lysed in NP-40 lysis buffer on ice for 30 min.Immunoblotting was performed following the protocols described previously[15].The membranes were incubated with the primary antibodies(mouse monoclonal antibodies against vimentin,Snail,Twist,or E-cadherin,purchased from Santa Cruz Biotechnology and used at the dilution of 1:800) at 4 ℃overnight and then with matching horseradish peroxidase-conjugated secondary antibody (mouse monoclonal antibody against α-tubulin,obtained from Cell Signaling Technology and diluted at 1∶1000) at 25 ℃for 1 h.The bands of the target proteins were visualized using the ECL detection system (Tanon GIS,Shanghai,China),with α-tubulin as the internal control.All Western blot analyses were performed in triplicate and ImageJ software package (National Institutes of Health,USA) was used for densitometric analysis of the protein blots.
Transwell assay
All statistical analyses were performed using Graphpad Prism 8.0 Software.Data are presented as theMean±SD.Student'sttest was used for comparison between two groups,and comparison among more groups was carried out using one-way ANOVA.APvalue less than 0.05 was considered to indicate a statistically significant difference.
Data from GEPIA comparing the transcription level of SOX2-OT between 486 lung squamous cell carcinoma(LUSC) and 338 adjacent normal tissues indicated that SOX2-OT was significantly overexpressed in LUSC as compared with normal tissues (Fig.1A).We also compared the expression levels of SOX2-OT in LAD cell lines(A549 and H1299)and SCC cell lines(SW900 and H520),and the results of qRT-PCR showed that the expression of SOX2-OT was significantly higher in H520 cells than in A549,H1299,and SW900 cells (Fig.1B).We thus used H520 cells for subsequent loss-of-function experiments.
Wound-healing assay
Total RNA was extracted from the cells,purified and then reverse transcribed into first-strand cDNA using TransScript All-in-One First-Strand cDNA Synthesis kit(TransGen Biotech,Beijing,China).Real-time quantitative PCR (qRT-PCR) was carried out using SYBR Premix Ex Taq (Takara,Kyoto,Japan) with β-actin as the inner control.The RNA samples were denatured for 5 min at 94 ℃followed by 35 thermal cycles of 30 s at 94 ℃,30 s at 53 ℃,and elongation for 30 s at 72 ℃.The reaction was terminated at 25 ℃for 5 min.Data from qRT-PCR were collected using a Bio-Rad CFX96Touch Real-time PCR Detection System(Bio-Rad,Hercules,CA,USA),and analyzed using the 2-ΔΔCtmethod.The primers used for amplification of the target mRNAs are listed in Tab.1.
Bioinformatic analysis
The young man was terribly afraid of being recognised by someone in this strange position, and would have turned on his heel and gone away had it not been for the thought of Minon-Minette
The Gene Expression Profiling Interactive Analysis(GEPIA http://gepia.cancer-pku.cn/) database was used for analysis of the expression levels of SOX2-OT in LUSC.The binding sites between SOX2 and SOX2-OT were predicted by StarBase (https://starbase.sysu.edu.cn/).The cBioPortal for Cancer Genomics (cBioPortal http://www.cbioportal.org/) database was used for correlation analysis between SOX2 and SOX2-OT.
Statistical analysis
Transwell assay was performed as described previously[3].Briefly,200 μL serum-free Dulbecco's Modified Eagle Medium (DMEM,Thermo Fisher Scientific) containing 3000 cells and 500 μL DMEM containing 20%FBS were added in the upper and lower chambers of the 24-well plate (Transwell,Corning,Corning,NY,USA),respectively.After incubation at 37 ℃for 72 h,the Transwell chamber was removed and fixed in paraformaldehyde for 20 min followed by crystal violet staining.Cells that migrated through the membrane were counted under a microscope(Nikon,Tokyo,Japan).
The wolf, seeing her come in, said to her, hiding himself under the bedclothes, Put the cake and the little pot of butter upon the stool, and come get into bed with me. 7
RESULTS
SOX2-OT is highly expressed in LUSC specimens and H520 cells
And it is well known that if the yard-cock belonging to this family happened to crow at midnight, they would declare it was morning, although the watchman and all the clocks in the town were proclaiming the hour of twelve at night
SOX2-OT knockdown suppresses migration and invasion of H520 cells
In H520 cells,transfection with shRNAs #4 and #5 against SOX2-OT significantly reduced SOX2-OT expression by 79% and 62%,respectively,as shown by qRT-PCR (Fig.1C).Wound healing assay and Transwell migration assay demonstrated significant reductions in the migration rate and the number of invasive cells following SOX2-OT knockdown in H520 cells(Fig.2).Of the two shRNA constructs,shRNA #4 showed a stronger efficacy for SOX-OT knockdown and more effectively attenuated migration capacity of H520 cells than shRNA#5,and shRNA#4 was thus used for SOX-OT knockdown in H520 cells in subsequent Gli1 overexpression experiment(Fig.4D-F).
EVEN a prairie fire it survives; at a zephyr s breath it revives -- such is the resilience of life in grass that I came to appreciate when reading poetry in my youth
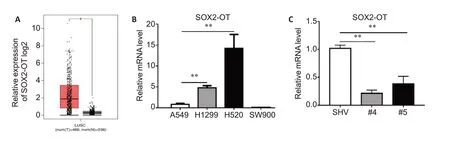
Fig.1 Expression level of SOX2-OT mRNA in lung squamous cell carcinoma(LUSC)specimens and in different lung cancer cell lines.A: Scatter plot generated by GEPIA for comparing transcription levels of SOX2-OT between 486 LUSC and 338 adjacent normal tissues.B: qRT-PCR analysis showing aberrantly higher SOX2-OT expression in H520 cells than in the other cell lines.C:SOX2-OT transcription level in H520 cells after transfection with two shRNAs (#4 and #5) for SOX2-OT knockdown and the empty vector (SHV).β-actin was used as the internal control.All data are presented as Mean±SD if not specified otherwise.*P<0.05,**P<0.01.

Fig.2 Effect of SOX2-OT knockdown on invasive capacity of H520 cells.SOX2-OT knockdown significantly inhibits metastatic capacity of H520 cells as shown by wound healing assay (A,B;original magnification:×100)and attenuates invasiveness of the cells as shown by Transwell assay(C,D;×200).**P<0.01.
SOX2-OT promotes H520 cell migration by activating Gli1-mediated EMT phenotype
The lung cancer cell lines H1299,A549,H520,and SW900 were purchased from the American Type Culture Collection (Manassas,USA) and maintained in our laboratory.All the cell lines were cultured in RPMI-1640 medium supplemented with 10% fetal bovine serum(Hyclone,Logan,UT,USA) and penicillin/streptomycin(100 mg/mL)at 37 ℃with a relative humidity of 95%and 5%CO2.

Fig.3 Changes of expressions of EMT-related genes inH520 cells with SOX2-OT knockdown detected by qRT-PCR (A) and Western blotting (B,C) with β-actin and α-tubulin as the inner control,respectively.*P<0.05,**P<0.01.
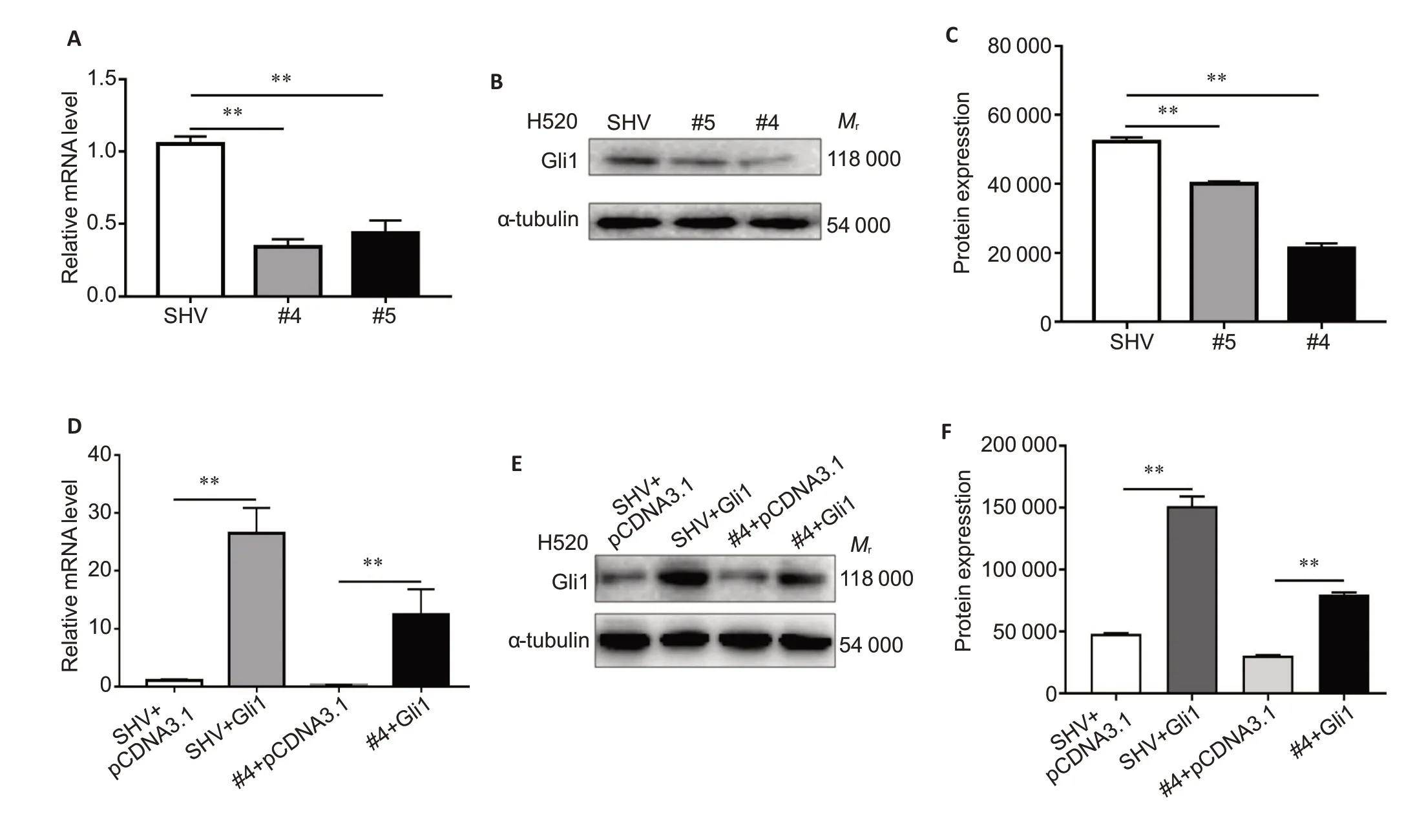
Fig.4 Effect of modification of SOX2-OT expression on Gli1 expression in H520 cells.SOX2-OT knockdown in H520 cells down-regulates expressions of Gli1 mRNA and protein detected with qRT-PCR (A) and Western blotting (B,C).Transfection with Gli1-overexpressing plasmid (pcDNA3.1-Gli1) counteracts the inhibitory effect of SOX2-OT knockdown shown by qRT-PCR (D) and Western blotting (E,F).Cells co-transfected with SHV and pcDNA3.1 backbone served as the control.**P<0.01.
Gli1 overexpression promotes H520 cell migration and reverses SOX2-OT knockdown-induced EMT phenotype
Using a Gli1 gain-of-function system,we first investigated whether Gli1 overexpression stimulated the migration of H520 cells.Both wound healing assay and Transwell migration assay demonstrated that Gli1 overexpression enhanced the invasiveness of H520 cells as compared with empty vector-transfected cells (Fig.5A and C).Gli1 overexpression also strongly promoted the metastatic capacity of H520 cells (Fig.5B and D).In H520 cells with SOX2-OT knockdown,Gli1 overexpression obviously reversed the inhibitory effect of SOX2-OT knockdown on cell metastasis (Fig.5A-D),markedly increased the expression levels of vimentin and Snail,and lowered the expression of E-cadherin.In the co-transfected cells,Gli1 overexpression rescued the EMT phenotype in H520 cells induced by transfection with shRNA#4(Fig.5E).

Fig.5 Gli1 overexpression attenuates the inhibitory effect of SOX2-OT knockdown on cell invasion and EMT-related phenotype in H520 cells.A,B:Wound healing assay of H520 cells transfected with both shRNA against SOX2-OT(#4)and pcDNA 3.1-Gli1 (×100).C,D: Transwell assay of H520 cells of H520 cells transfected with both shRNA against SOX2-OT(#4)and pcDNA 3.1-Gli1(×200).E:mRNA expression levels of vimentin,Snail and E-cadherin in H520 cells with different treatments detected with qRT-PCR.**P<0.01.
SOX2-OT positively modulates Gli1 by protecting SOX2 from miR-200c-mediated interference
The results of bioinformatic analysis did not reveal any microRNA that bridge SOX2-OT and Gli1,suggesting an indirect regulatory mechanism of SOX2-OT.As an upstream transcription factor of Gli1,SOX2 was predicted by StarBase to be modulated by SOX2-OTviamiR-200c (Fig.6A).The results of qRT-PCR demonstrated that the transcription of SOX2 and SOX2-OT was inhibited by miR-200c mimic and enhanced by miR-200c inhibitor (Fig.6B).To further test weather SOX2-OT regulates SOX2viamiR-200c,we co-transfected the cells with miR-200c inhibitor and SOX2-OT knockdown plasmid,and the results showed that miR-200c inhibitor counteracted SOX2-OT knockdown-induced downregulation of SOX2 and SOX2-OT (Fig.6C).Consistent with these data,analysis of 178 lung cancer specimens from cBioPortal database demonstrated a positive correlation between the expression of SOX2 and SOX2-OT (Fig.6D).Furthermore,SOX2 overexpression attenuated SOX2-OT knockdown-induced downregulation of GLI1,suggesting that SOX2-OT positively regulates Gli1 through SOX2(Fig.6E).

Fig.6 SOX2-OT positively modulates SOX2 by sponging miR-200c in H520 cells.A:Bioinformatic analysis of the binding sites of miR-200c on SOX2 and SOX2-OT predicted by StarBase.B:Transcription of SOX2 and SOX2-OT is inhibited by miR-200c mimic and elevated by miR-200c inhibitor.C: miR-200c inhibitor counteracts SOX2-OT knockdown-induced downregulation of SOX2 and SOX2-OT.D: Expressions of SOX2 and SOX2-OT are positively correlated based on data from cBioPortal database.E: SOX2 overexpression attenuated SOX2-OT knockdown-induced inhibition of GLI1 expression.**P<0.01.
DISCUSSION
Previous studies reported that SOX2-OT was aberrantly overexpressed in the tumor mass and plasma of lung cancer patients and was associated with a poor prognosis[19-21].In this study,we further confirm the high expression of SOX2-OT in both LUSC tissues and the LUSC cell line H520.
“Even if I had the money, there was simply no way of replacing those glasses. Along with not being able to see well, I experienced headaches every day, so my coworkers and I were much in prayer about this. Then your crates arrived. When my staffed removed the covers, they found a pair of glasses lying on top.”
In vivoandin vitrostudies have shown that SOX2-OT regulates the proliferation of various tumor cells and tumor stem cells and participates in drug resistance and metastasis of the tumor cells[22,23].Inhibition of SOX2-OT sensitizes lung cancer cells to erlotinib or cisplatin-based therapy by modulating the activation of AKT/ERK in the EGFR pathway[24].In this study,we found that SOX2-OT transcription level was significantly higher in H520 cells than in other cell lines and revealed the role of SOX2-OT for positive regulation of cell invasion using a loss-of-function phenotypic assay.We previously found that the expression levels of the key components in the Hedgehog signaling pathway were negatively correlated with the prognosis of lung cancer patients,and inhibition of this pathway attenuated the proliferation and metastasis of lung cancer cells bothin vitroandin vivo[15].Interference of Gli1 expression was found to inhibit the proliferation of lung adenocarcinoma stem cells and induce their apoptosis,suggesting the importance of Gli1 in maintaining the stemness of lung cancer stem cells[25].Gli1was reported to promote the invasion and infiltration ability of lung cancer cells by abnormally upregulating EMT markers (such as vimentin,Twist,and N-cadherin),and treatments targeting Gli1 may potentially reduce EMT-mediated metastasis of lung cancer and other tumor cells[17,18,26].In addition to the autocrine-related modulatory mechanism,breast cancer cells that undergo EMT affect the tumor microenvironment also through paracrine Gli1,thereby promoting the metastasis of the non-EMT cells[27].Our data demonstrate that SOX2-OT regulates the expression of Gli1,and an enhanced Gli1 expression attenuates SOX2-OT knockdown-induced suppression of cell migration and EMT,suggesting that SOX2-OT regulates cell invasion via Gli1.
lncRNAs usually act as competing endogenous RNAs (ceRNA) to sponging microRNAs and protect candidate RNAs from degradation,and these ceRNAs are capable of activating downstream target genes through microRNA (miRNA) sponge vectors[28].SOX2-OT also acts as a ceRNA to promote cancer cell invasion and metastasis in various cancer types[29-33]and has been shown to promote stemness and cell invasion of glioblastoma and hepatocellular carcinomaviamiR-122 sponge vectors[23,34].In prostate cancer cells,SOX2-OT knockdown inhibits cell metastasis by releasing miR-452-5p and disabling the Wnt/β-catenin pathway[35].SOX2-OT was also shown to stimulate lung cancer cell migration and invasion via miR-132 sponge vectors[36].Previous studies suggest that SOX2-OT positively regulates SOX2viamiR-200c in pancreatic ductal adenocarcinoma cells[37],and targeted inhibition of SOX2 can significantly inhibit the proliferation of lung cancer stem cells and suppress malignant behaviors of the tumor cells[3,4,38,39].In melanoma cells,SOX2 activates Gli1 through directly binding to the region-423 bp upstream the transcriptional starting site to enhance GLI1 transcription[40].Our data reveal that in H520 cells where SOX2-OT expression is upregulated,SOX2-OT positively modulates Gli1 by sponging miR-200c and protecting SOX2 from interference;but this mechanism does not exclude the possibility that other mechanisms exist by which SOX2-OT either directly or indirectly modulates Gli1 expression.Nevertheless,our findings still highlight the pivotal role of the SOX2-OT/Gli1 axis in regulating LUSC cell invasion,indicating the potential of the SOX2-OT/Gli1 axis as a candidate target for clinical diagnosis and therapy of lung cancers.