
遮荫对青钱柳苗生长和黄酮类化合物积累的影响
2022-10-25 12:32:26于邦友刘可可夏培兴
安徽农业大学学报 2022年3期
邱 璐,于邦友,刘可可,夏培兴,邓 波*
(1. 安徽农业大学林学与园林学院,合肥 230036; 2. 阜阳市林业科学技术推广站,阜阳 236015)
青钱柳(Cyclocarya paliurus(Batal.) Iljinskaja)是胡桃科青钱柳属高大落叶乔木,是我国特有的木本药用植物,广泛分布于我国的亚热带地区,喜光,适宜生长在温暖湿润的地方。其叶片可制茶,以叶片作为原料的保健品也广受市场欢迎[1]。以往研究表明,青钱柳叶可以提取具有降血压、降血糖、抗氧化等药理保健功能的生理活性物质,包括黄酮类、多糖、甾体类等,其中黄酮类化合物是青钱柳中主要的活性成分,也是药用青钱柳栽培中主要的目的产物[1-2]。随着近年来青钱柳产业化的快速发展,市场对青钱柳叶的需求日益增大,但青钱柳的自然资源比较稀少,主要以发展人工林为主。因此,优化青钱柳的栽培技术与环境调控措施以获得更丰富的产物成为目前人工林发展的必然需求。青钱柳作为一种木本药用植物,为了保证产量与原料品质,其生长状况和黄酮类化合物的积累都值得关注,所以要重点探究影响青钱柳生长与次生代谢物积累的光照环境条件,以提高、稳定产物的品质与产量。
光为植物光合作用提供能量,是影响植物生长发育与次生代谢物积累的重要环境因素之一,光照强度的改变能够直接影响植物的光合作用,从而对植物体内营养物质积累、植物生长发育产生显著影响[3-4]。面对光照环境的改变,植物也能够通过调整生物量分配、光合结构、形态建成、次生代谢物积累模式等来提高自己抗逆境的能力,实现对环境变化的适应[5-7]。目前有许多关于光强对植物生长、黄酮类化合物积累影响的研究,如相比于遮荫条件,全光照可以显著增加麻栎(Quercus acutissimaCarruth) 和闽楠(Phoebe bournei( Hemsl.) Yang) 单株总叶面积和地上、地下生物量及总生物量,并显著影响蒲公英(Taraxacum mongolicumHand-Mazz)与银杏(Ginkgo bilobaLinn.)叶片中黄酮类化合物的积累,而遮荫显著影响了青钱柳黄酮类化合物的积累高峰[8-11]。综上可见,调控光照强度能够对植物的生长发育与黄酮类化合物积累产生显著影响,且每种植物都有最有利于其生长与次生代谢物积累的光照条件。
在生产中,青钱柳主要的收获部位为叶片,但自然生长的青钱柳树形高大,难以采摘,因此为保证叶片产量与采集效率,青钱柳的矮化以及密植管理十分重要。本文通过遮荫的方法,探究青钱柳的生长、黄酮类化合物的积累对光强的响应规律和有利于药用植物产量积累与品质提高的环境条件,以期为选择适合青钱柳生长与次生代谢物积累的栽培环境提供理论依据。
1 材料与方法
1.1 试验区概况
试验在安徽省合肥市林业高科技开发中心进行。试验区属于亚热带湿润季风气候,四季分明,气候温和,年平均温度为15.7 ℃,平均相对湿度为77 %。雨量适中,年平均降水量为900~1 000 mm,主要集中在5—6 月的梅雨季。年日照时间约为2 000 h,年均无霜期为228 d。
1.2 供试材料
试验材料取自安徽省宣城市绩溪县,位于北纬29°57′~30°20′,东经118°20′~118°55′之间、黄山山脉和西天目山山脉结合带,属北亚热带季风气候,年平均气温15.9 ℃,年平均降水量1 630.3 mm。于2018 年3 月将1 年生青钱柳幼苗移植至安徽省合肥市林业高科技开发中心的基地大田中,于2019 年4月选取其中长势均匀的青钱柳幼苗进行遮荫处理。
1.3 试验设计
采用单因素试验设计,使用50%遮阳率的遮阳网设置3 种光照处理,分别为不遮荫、50%遮阳率遮阳网遮荫一层(光照强度约为全光下的42%)、50%遮阳率遮阳网遮荫两层(光照强度约为全光下的19%)。因此,本研究共包括3 个处理,每个处理包括3 个重复,共9 个试验单元,每个单元包含20 株苗。自2019 年4 月中旬开始对上述青钱柳苗进行遮荫处理,遮阳网搭置于距地面约2 m 高处。连续遮荫3 个月后,于7 月测量各处理下青钱柳苗的苗高、地径、光合和荧光参数、叶绿素含量、生物量、黄酮化学组织定位、黄酮含量和黄酮产量。
1.4 研究方法
1.4.1 苗高和地径生长测定 于2019年7月对所有青钱柳苗进行苗高和地径测定,并计算各光照处理下的平均苗高和地径生长状况。
1.4.2 光合和荧光参数测定 根据苗高和地径的测量结果,选取各处理中接近平均值的5 株青钱柳苗,挑选从顶芽向下数的第5 片复叶中间的完全功能小叶,作为测量对象进行标记,光合参数采用LI-6400XT 光合系统(LI-COR,Inc. Lincoln NE,USA),于自然光条件下,每天早上9:00—11:00 测定,测定参数为净光合速率(μmol·(m-2·s-1))。
荧光参数用 FMS2 便携脉冲调制式荧光仪(Hansatech instruments Ltd. Norfolk, United Kingdom)对之前标记的小叶进行测定。初始荧光Fo于等待植株暗适应30 min 后测定,暗适应最大荧光Fm用饱和脉冲照射后测得,经过测量光照射一段时间后,植株每隔30 s 照射1 次饱和脉冲,测得光适应最大荧光F′m,此时关闭测量光,并打开远红光,测量得Fo′。测量结果按以下公式计算:PSⅡ原初光能转化效率Fv/Fm=(Fm-Fo)/Fm;PSⅡ光能捕获效率Fv′/Fm′ =(Fm′-Fo′)/Fm′。
1.4.3 叶绿素含量测定 采集用于测定光合与荧光参数的同一批所有植株同一部位的新鲜小叶,按单株分类,清洗、擦干、切碎后,用85%丙酮溶液浸透处理,分别在λ= 663 nm 和λ= 645 nm 下测定吸光度。叶绿素含量按以下公式计算:Cchl-a= 12.7A663-2.69A645;Cchl-b= 22.9A645-4.64A663;Cchl-a+b=20.2A645+ 8.02A663。
1.4.4 生物量测定 根据青钱柳苗高和地径的测定结果,选取不同光照处理中接近平均值的3 株青钱柳苗,按单株采集根、茎秆和叶片3 个部分,洗净后于70 ℃烘干,分别称重,单株总生物量为其根、茎秆和叶片的干重总和。根冠比计算公式为:根冠比 = 根生物量/茎叶生物量。
1.4.5 黄酮组织化学定位 在每个处理中选取5 株青钱柳苗,摘取植株同一部位的完全功能叶进行组织化学分析。分析方式参照廖云海等描述的方法,用1 %的三氯化铝乙醇溶液将包含中脉的叶片冷冻切片(厚度为20 μm)染色10 min,封片后立即放置于荧光显微镜下观察并分组拍照[12]。在荧光显微镜下观察黄酮类化合物时,会呈现蓝白色荧光。
1.4.6 黄酮含量测定 首先均匀采集每个试验单元中所有青钱柳苗的中上部位的完全功能叶,相同光照处理下的所有小叶组成一个样品,重复采集3 次,共采集9 组样品。然后去除叶柄,烘干、磨碎。每份样品取2 g 左右,用石油醚进行4 h 的80 ℃水浴提取,过滤,将固形物室温晾干,作为黄酮提取样品待用。黄酮的提取采用超声波辅助提取法[13],将样品用75%乙醇水溶液65 ℃超声提取45 min,重复2 次,将2 次提取液混合,挥发一段时间,用甲醇滴洗固形物并定容至10 mL,作为样品待测。
总黄酮的测定采用比色法[14],于室温下取1 mL样品置于10 mL 容量瓶,加入0.3 mL 5% NaNO2溶液,静置5 min 后加入0.3 mL 1% AlCl3甲醇溶液、1 mol·L-1NaOH 溶液混匀,显色15 min 后在410 nm处测定吸光度。以芦丁标准品(中国药品生物制品检定所,北京)为对照品,计算样品中的总黄酮含量(线性范围:芦丁2.3~56.0 μg·mL-1,R2> 0.99),结果以每克青钱柳烘干样品中含有相当于芦丁当量的毫克数表示。
黄酮单体的测定采用HPLC - DAD 法,共测定槲皮素、异槲皮苷和山奈酚3 个黄酮单体。主要采用Agilent 1200 系列液相色谱(Waldbronn,德国)进行分析,分析前将样品过0.45 μm 有机滤膜。槲皮素和山奈酚参照《中华人民共和国药典》[15]中银杏黄酮水解的方法,用酸将样品水解为苷元后待测。取1 mL 样品,挥干溶剂后加入25 mL 体积为4 : 1的甲醇-25% HCl,加热回流30 min,待冷却后定容至25 mL,检测时用VA:VB= 55 : 45 等度洗脱(A-色谱纯甲醇,B-0.3%磷酸),检测波长为365 nm,柱温为30 ℃,流动相流速为1.0 mL·min-1。异槲皮苷则将提取液过滤后直接进行分析,检测的流动相为A-色谱纯甲醇,B-0.5 %磷酸。梯度洗脱程序为:0~25 min,15% A;15~26 min,15%~90% A;26~36 min,90% A;36~37 min,90%~15% A;37~45 min,15% A。检测波长350 nm,柱温、流动相流速与槲皮素和山奈酚一致。试验中用于获得外标校准曲线的异槲皮苷购自中国药品生物制品检定所(北京),槲皮素和山奈酚购自Sigma- Aldrich 公司(St. Louis,MO,USA)。根据黄酮含量计算出黄酮产量:黄酮产量/(mg·株-1)= 黄酮含量/(mg·g-1)× 叶生物量/(g·株-1)。
1.5 数据处理与统计分析
本试验以单因素方差分析结果来比较环境对青钱柳生长、生物量、荧光参数、黄酮含量和黄酮产量等相关数量指标的影响;以Duncan 多重比较来分析处理之间的差异。所有统计数据均由SPSS 22.0软件进行方差分析,用Excel 作图,分析结果的差异显著性检验水平P< 0.05。
2 结果与分析
2.1 遮荫处理对青钱柳苗高和地径的影响
遮荫处理显著影响了青钱柳的苗高和地径生长(P< 0.05)。如图1 所示,随着遮荫强度增大,青钱柳的苗高生长量呈先增高后下降的趋势:一层遮荫 > 二层遮荫 > 全光,轻度遮荫促进了青钱柳苗的苗高生长,达到最大生长量,而二层遮荫下青钱柳的苗高生长受到了抑制。地径整体变化趋势与苗高不一致,地径生长量一直随着遮荫程度的加强而下降。
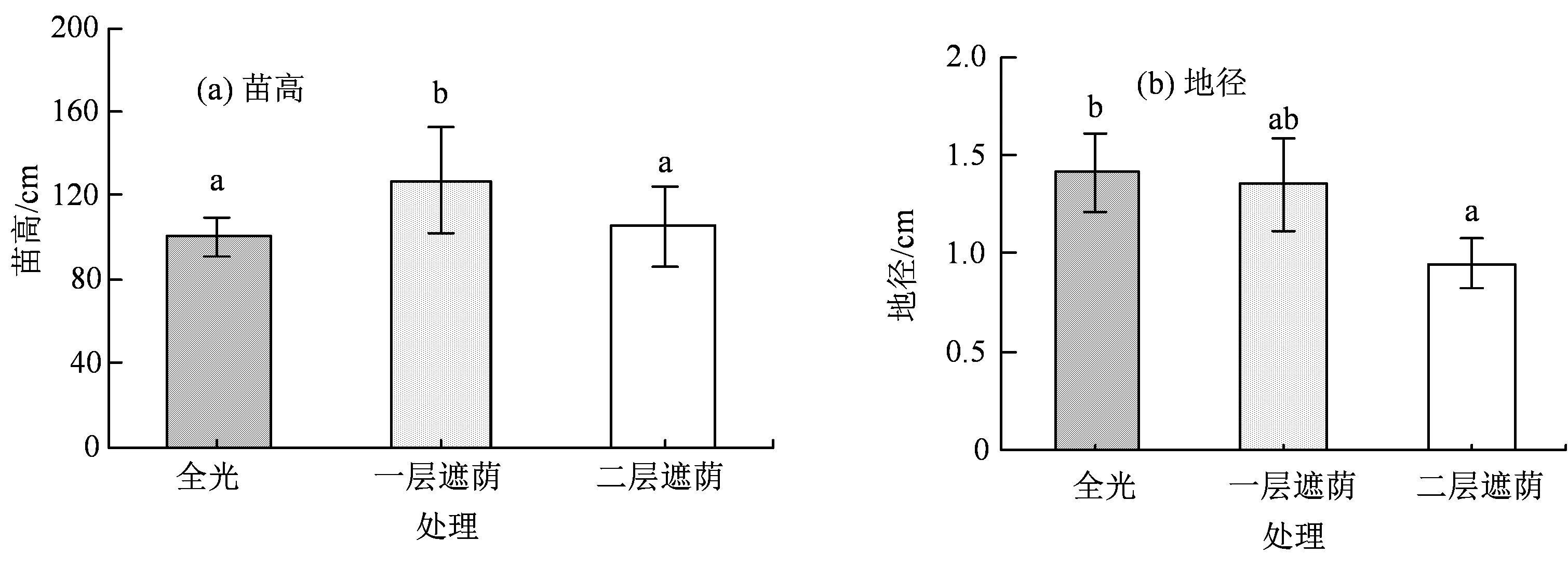
图1 不同遮荫强度下青钱柳的苗高和地径Figure 1 The seedling height and rootcollar diameter growth of C. paliurus (Batal.) Iljinskaja under three different shade intensities
2.2 遮荫处理对青钱柳生物量积累与分配的影响
不同的遮荫处理对青钱柳生物量的积累和分配产生了显著影响(P< 0.05,图2)。青钱柳苗的茎秆、叶片和总生物量均在一层遮荫条件下达到最大,但于全光下无显著差异,而两层遮荫下的生物量积累明显小于全光和一层遮荫下。说明适度遮荫可以促进青钱柳苗的生长,但遮荫过度则会强烈抑制其生长。另外,遮荫处理使青钱柳苗的地上和地下部分生物量的分配发生了明显变化。随着遮荫强度的加大,根冠比呈显著下降,这也反映了青钱柳苗在形态上通过提高地上部分生物量的分配以适应弱光环境。
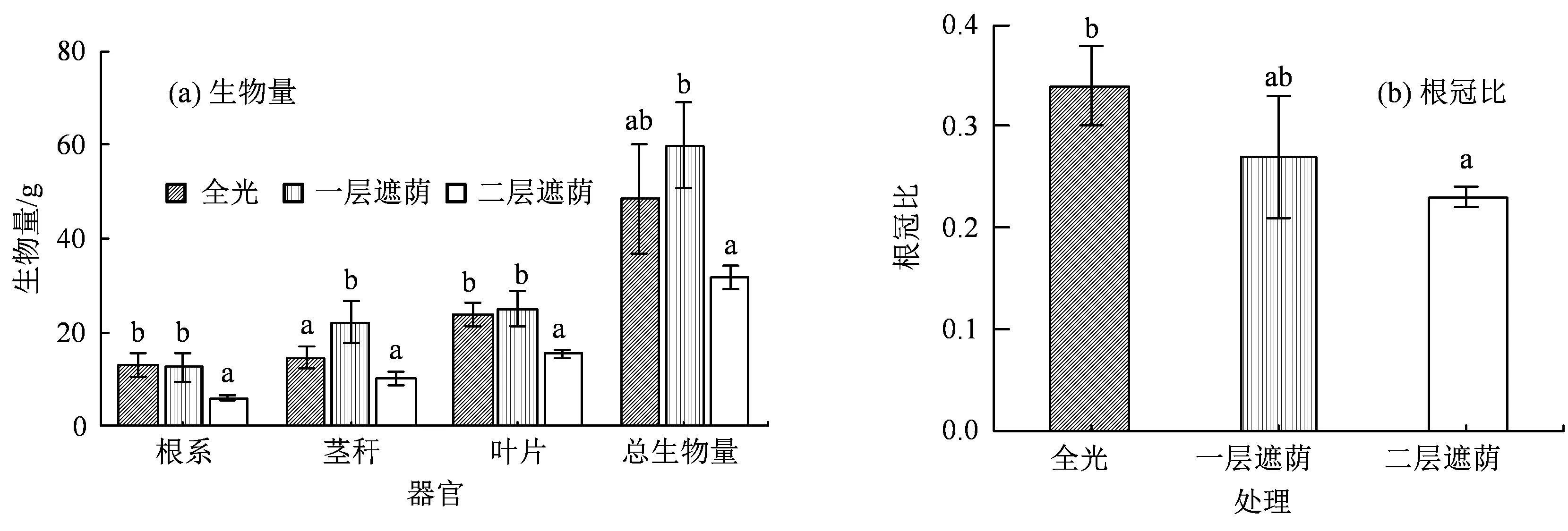
图2 不同遮荫强度下青钱柳不同器官中的生物量积累与分配Figure 2 The biomass accumulation and allocation in different organs of C. paliurus (Batal.) Iljinskaja under different shade intensities
2.3 遮荫处理对青钱柳光合和荧光参数的影响
遮荫对青钱柳光合与荧光参数产生了显著影响(P< 0.05,表1)。不同光照处理间青钱柳叶片的净光合速率随遮荫程度的加强而下降,变异范围为10.48~12.42 μmol·(m-2·s-1),全光时净光合速率分别是一层遮荫时的1.04 倍、二层遮荫时的1.19 倍,说明遮荫抑制了青钱柳的光合作用与光合作物的产出。为了反映遮荫处理对青钱柳叶片光能利用效率的影响,分别测定了 PSⅡ原初光能转化效率(Fv/Fm)、PSⅡ光能捕获效率(Fv'/Fm')两个荧光参数。结果显示,PSⅡ原初光能转换效率在3 种处理下均高于0.8,在二层遮荫时略高于全光和一层遮荫,而PSⅡ光能捕获效率总体随遮荫程度的加强呈上升趋势,在全光和一层遮荫时没有显著变化,二层遮荫时显著大于全光和一层遮荫时,为全光和一层遮荫下的1.52 倍(表1)。由此可见,随着光照的减弱,青钱柳叶片的光合机构也做出了适应性调整,提高了其对光能的利用效率。
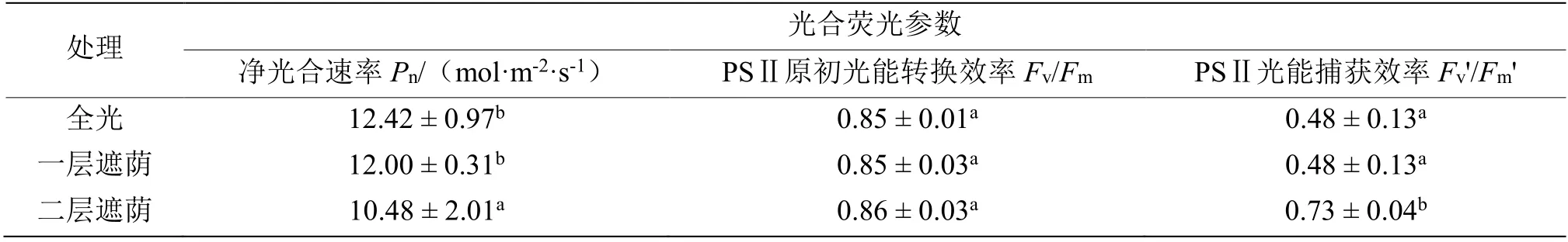
表1 不同遮荫强度下青钱柳的光合与荧光参数Table 1 The photosynthetic parameters and fluorescence parameters of C. paliurus (Batal.) Iljinskaja under different shade intensities
2.4 遮荫处理对青钱柳叶绿素含量的影响
由图3 可知,遮荫对青钱柳叶片的光合色素含量与组成产生了显著影响(P< 0.05),青钱柳叶绿素含量一直随遮荫程度的加强而增加,而叶绿素a/b总体呈下降趋势,说明遮荫下的叶绿素b 的相对比例上升。叶绿素b 是吸收光能的重要光合色素之一,这与上一部分的结果一致,可见面对弱光胁迫,青钱柳植株的光合机构会通过提高光合色素的含量来增强对光能的捕获吸收,从而实现对光环境的适应。
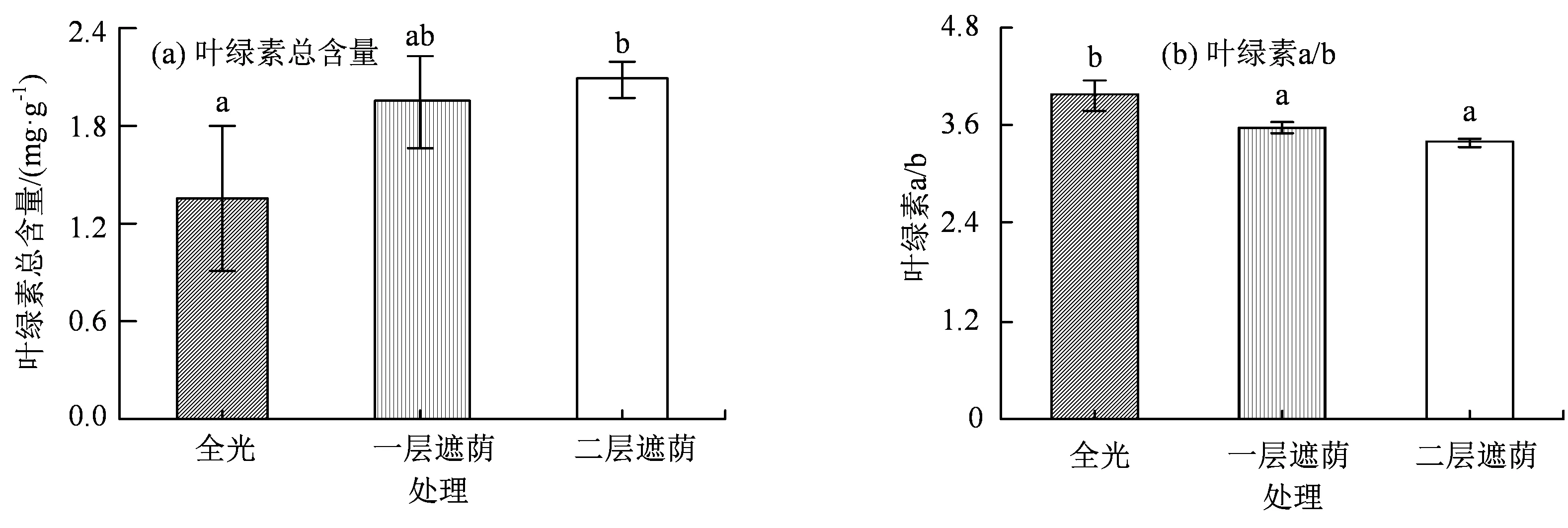
图3 不同遮荫强度下青钱柳的叶绿素含量与叶绿素a/b 的变化Figure 3 The total chlorophyll content and the chlorophyll a/b ratio of C. paliurus (Batal.) Iljinskaja under different shade intensities
2.5 遮荫处理对青钱柳黄酮含量的影响
遮荫处理对青钱柳中的总黄酮和黄酮单体(异槲皮苷、山奈酚和槲皮素)含量均产生了显著影响(P< 0.05)。如图4 所示,异槲皮苷、山奈酚和槲皮素的含量有着相同的变异趋势,即随着遮荫程度的增强而下降,变异区间分别为异槲皮苷为1~5.73 mg·g-1,山奈酚为0.57~1.79 mg·g-1,槲皮素为0.15~1.11 mg·g-1,全光下的黄酮单体含量显著大于一层遮荫与二层遮荫下。3 种处理下的黄酮单体含量总体上呈异槲皮苷 > 山奈酚 > 槲皮素,且全光下的异槲皮苷含量远高于山奈酚和槲皮素的含量,可见异槲皮苷为主要的黄酮单体。黄酮单体下降幅度整体上呈槲皮素 > 异槲皮苷 > 山奈酚,猜测槲皮素可能为三者之中最有效的光保护剂。总黄酮含量随遮荫强度增加呈下降趋势,全光下的总黄酮量显著大于一层遮荫与二层遮荫。综上可见,遮荫处理使得青钱柳叶中黄酮类化合物的积累受到了抑制。
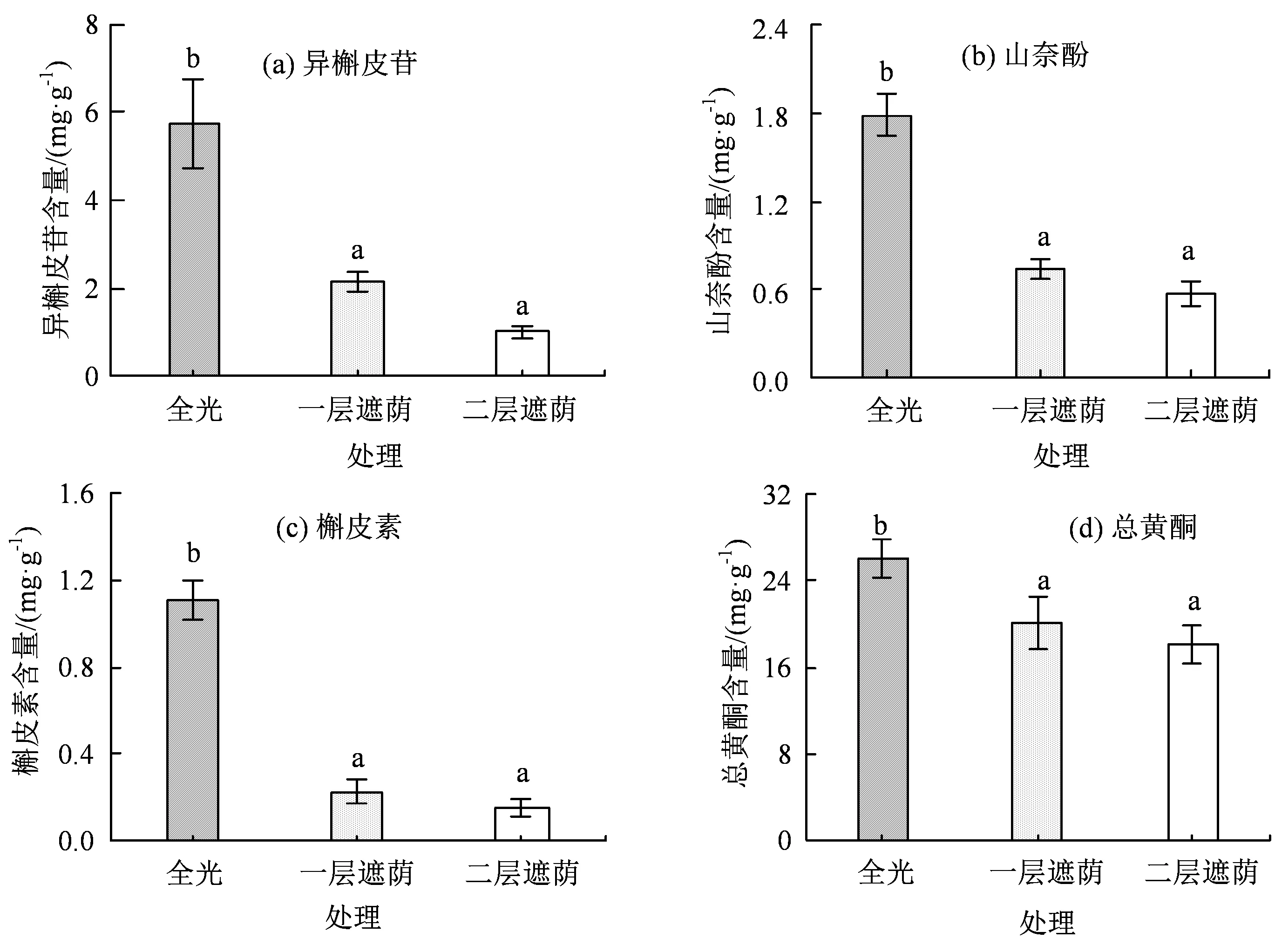
图4 不同遮荫强度下青钱柳中的3 个黄酮单体与总黄酮含量Figure 4 The contents of three individual flavonoids and total flavonoid in C. paliurus (Batal.) Iljinskaja under different shade intensities
2.6 青钱柳中黄酮的组织化学定位分析
不同的遮荫强度下的青钱柳叶片黄酮积累模式基本一致,并且随着遮荫程度的加深,黄酮的积累量逐渐减少。如图5 所示,全光下黄酮主要分布在叶肉上表皮部分和主脉部分的微管组织中,包括副维管束、次生韧皮部和次生木质部,腺毛中也有部分分布。随着光照的减弱,黄酮在表皮上的积累量逐渐减少,更多的储存在微管组织和腺毛中。可见黄酮是重要的抗光氧化物质,能够感知光照胁迫,并富集在受光照胁迫的部位。
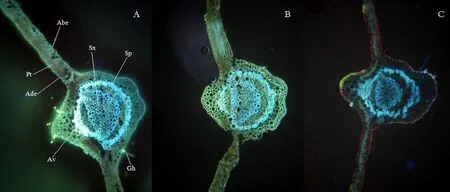
图5 不同遮荫强度下青钱柳中黄酮积累的组织化学定位Figure 5 Histochemical analysis of flavonoid accumulation in leaves of C. paliurus (Batal.) Iljinskaja
2.7 遮荫处理对青钱柳黄酮产量的影响
遮荫对总黄酮和黄酮单体(异槲皮苷、山奈酚、槲皮素)产量均产生了显著性影响(P< 0.05)。如图6 所示,总黄酮和各黄酮单体的产量均随遮荫水平的增强而呈下降趋势。总黄酮产量在全光下为一层遮荫时的1.22 倍,二层遮荫时的2.22 倍;槲皮素产量表现出较大的下降幅度,在全光时为一层遮荫的2.56倍,二层遮荫的8.77 倍;异槲皮苷产量在全光时为一层遮荫的2.29 倍,二层遮荫的4.82 倍;山奈酚产量在全光下为一层遮荫的4.76 倍,二层遮荫的11.39倍。青钱柳总黄酮和3 个黄酮单体产量均在一层遮荫时呈现较大的下降幅度,于二层遮荫时达到最小值,此时山奈酚产量的下降幅度最大,达到了91.2%;槲皮素次之,为88.6%,然后是异斛皮苷,为79.2%。由此可见,光照环境的变化对青钱柳的生物量积累与黄酮类化合物的产量产生了重要影响,且不同的黄酮单体产量对遮荫的响应也不同。
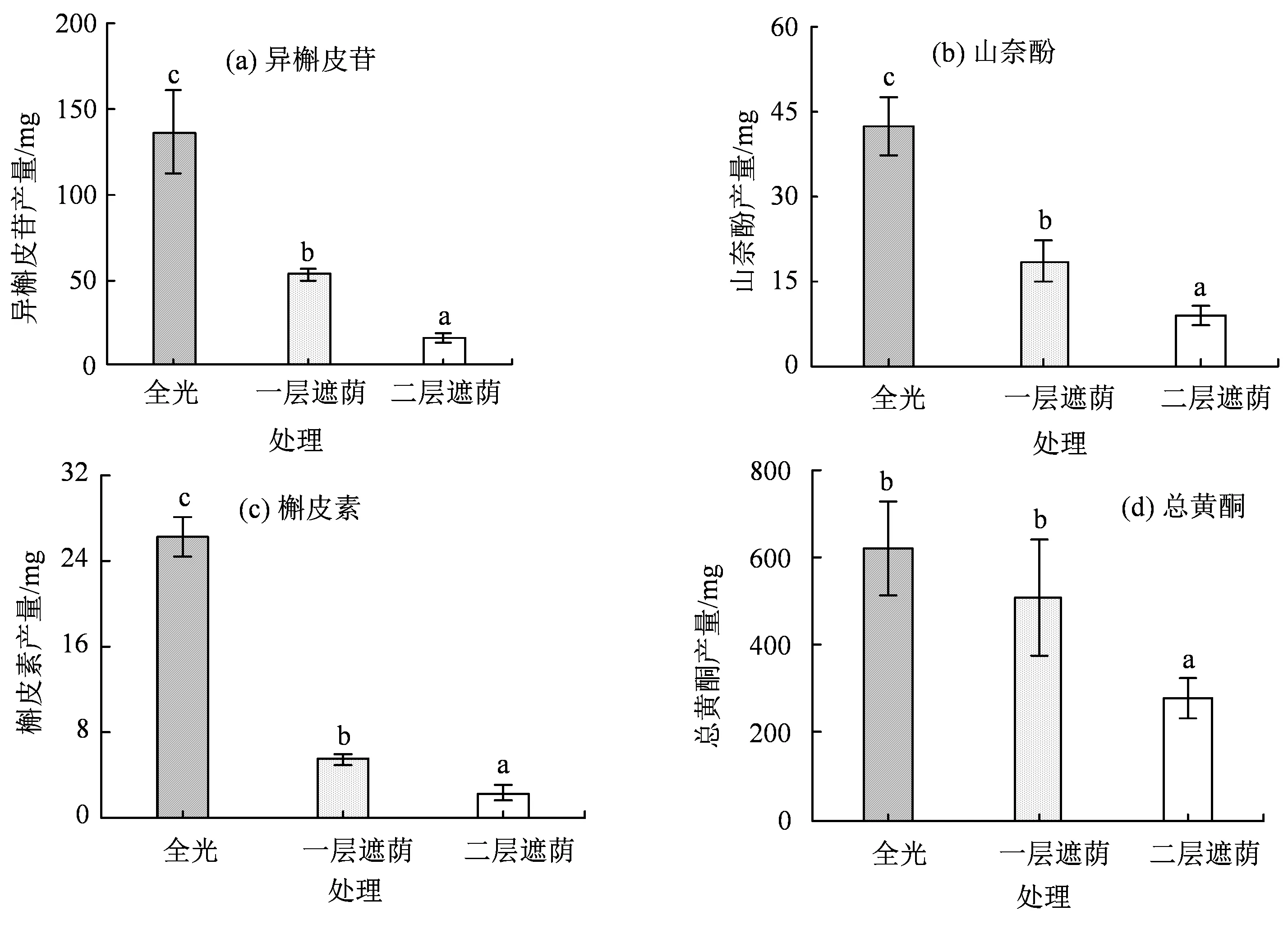
图6 不同遮荫强度下青钱柳的总黄酮和黄酮单体的单株产量Figure 6 The yield per plant of total flavonoid and three individual flavonoids of C. paliurus (Batal.) Iljinskaja under different shade intensities
3 讨论与结论
当植物受到弱光胁迫时,光合能力受到限制,影响了光合产物的合成,从而植株的苗高、地径生长也会受到不同程度的抑制,而为了适应光环境变化,植物也会在形态学水平上进行适应性调整[16-18]。本研究中,轻度遮荫可促进青钱柳苗高生长,但遮荫一直抑制了地径的生长。之前已有一些研究发现,适度遮荫的环境更有利于桤木(Alnus formosanaBurk)幼苗的生长;青桐(Firmiana platanifolia(Linn.f.) Marsili)幼苗的苗高随遮荫程度的增加而升高,而地径随遮荫强度的增加呈递减趋势;遮光率50%时,福建柏(Fokienia hodginsii(Dunn)A. HenryetThomas)苗高和地径达到最大,在遮光率为87.5 %时最小[19-21]。这与本研究结果一致,环境条件的变化能够对植株的生长发育产生影响,同时,在一层遮荫下,青钱柳呈现出“瘦高”的形态特征,可见青钱柳为适应弱光环境更趋向于纵向生长,以增加光能的接收与利用面积。在生物量积累和分类上,植物在遮光环境下会减少根系在全株总干重的比例,有利于同化有机物的积累和呼吸消耗的降低[22-24]。本研究中,分析了对青钱柳对不同遮荫处理下根、茎、叶与总的生物量积累与分配,结果表明,总体上青钱柳的生物量积累随着光照强度的下降而显著下降,同时生物量的分配也发生了一系列变化。适度遮荫下,青钱柳茎秆、叶片生物量的积累得到了促进,而深度遮荫下的植物光合作用受到抑制,有机物的产出减少,生物量的积累显著下降。另外,青钱柳根冠比也随之下降,可见青钱柳逐渐减少了根系的物质分配,将更多的生物量分配给了茎秆与叶片,用于青钱柳树高与冠幅的增长,并减缓了地下部分的生长,实现了对弱光环境的适应。
光是影响植物光合作用最重要的环境因素,能够直接影响光合系统的结构与活性,引起叶绿素含量、叶绿素比值、光化学反应等变化[25-27]。研究结果显示青钱柳叶净光合速率随着遮荫程度的加深而降低,说明遮荫抑制了青钱柳叶片的光合作用产物的积累。在植物进行光合作用时,光合色素对光能的吸收和利用起到重要的作用,其含量与比例是体现植物对光环境适应和利用能力的重要指标。其中叶绿素b 主要吸收短波长蓝紫光,升高叶绿素b 的比例可以增强叶片在弱光环境中对短波长蓝紫光的利用能力。研究显示,遮荫可使连翘(Forsythia suspensa(Thunb.)Vahl.)的净光合速率、叶绿素a/b降低,光能利用效率、叶绿素含量增加,以增强在弱光环境下的生长能力[28],也导致崖柏(Thuja sutchuenensisFranch.)光能转换效率和叶绿素含量的增加[29]。本研究中,遮荫条件下叶绿素含量和叶绿素b 相对比例均显著高于全光条件下,随遮荫程度的增加而升高,这与前人的结果一致,证明了青钱柳能够通过提高其光合色素的含量来提高对光能的捕捉能力,以更好地吸收蓝紫光,对弱光环境进行适应。与此同时,PSⅡ原初光能转换效率Fv/Fm是体现植物光合生理状态的重要指标,体现了PSⅡ反应中心处于开放态时的量子产量和植物的受光抑制程度,一般为0.80~0.85,Fv/Fm值升高能够体现植物较高的光利用效率[30-31]。本研究中,Fv/Fm值的变异区间为0.85~0.86,深层遮荫下拥有更高的光能转换效率,说明弱光下叶绿体的光合电子传递链可以更加高效地吸收光能,用于光合作用。Fv'/Fm'值反映了 PSⅡ反应中心未完全开放情况下的PSⅡ光能捕获效率的反应,本研究中,随着遮荫程度的增加,Fv'/Fm'值于二层遮荫时显著增加,说明弱光环境下青钱柳叶片的PSⅡ反应中心开放程度加强,更多的光能被用于化学反应,从而提高了光能利用率。
黄酮是一种有效的抗氧化物质,当接受到更强的紫外辐射时,植物会诱导产生黄酮等紫外吸收物质来抵御紫外辐射的伤害,主要在细胞膜系统间进行转运,最后集中分布于合成部位、功效部位或贮藏部位的液泡、细胞壁中,因此,黄酮的积累主要受光照与紫外辐射的调节[32-35]。青钱柳叶的组织化学分析显示,强光下的黄酮类化合物主要分布在青钱柳叶片的主脉微管组织和叶肉部分的表皮细胞中,腺毛中也有部分分布。可见黄酮类化合物大部分被贮藏于叶脉中,当太阳辐射作用于叶片时,黄酮就被运输至需要进行光保护的叶表皮部位,防御太阳辐射。因此,在黄酮类化合物的提取作业中应注意保留黄酮富集部位,如叶脉、表皮,叶柄中也可能有一定的黄酮分布。本研究结果显示,全光条件下青钱柳总黄酮和3 个黄酮单体的积累量最高,黄酮含量受遮荫影响显著下降。前人研究显示,强光可以使黄酮合成的酶基因表达上升,酶的含量与活性因此提高,从而增加了黄酮类化合物的含量,其中,Idris 等[36]研究说明了强光可以增加薇甘菊(Mikania micranthaKunth)和平卧菊(Tridax procumbensLinn.)中总黄酮的含量, Cheng 等[37]的研究显示银杏(Ginkgo bilobaLinn.)的黄酮的形成、生物合成和含量均受到光强的影响,这些均体现了光强对黄酮的合成和积累产生的重要影响,说明高强度的光照更有利于黄酮的积累,并印证了黄酮在光保护机制中发挥的作用。本研究中,山奈酚、槲皮素和异槲皮苷的含量有相同的变化趋势,随遮荫程度的加强均显著下降。其中,异槲皮苷是青钱柳重要的黄酮单体,山奈酚的含量相对稳定,遮荫对槲皮素的含量影响最大。在Ko 等的研究中,光照处理对洋葱(Allium cepaLinn.)中槲皮素含量产生了更显著的影响[38],这与本试验的结果一致,说明不同的黄酮单体的积累对光照条件的响应存在差异。另外,近年来有研究显示,槲皮素及其苷类化合物具有较强的光保护作用,可以有效清除茶叶受光胁迫时产生的活性氧[39],进一步证明了槲皮素对于青钱柳来说是更有效的光保护剂。同时,青钱柳总黄酮与黄酮单体产量随着遮荫程度的增加而显著降低,可见强光环境更有利于黄酮产量的积累。不同光照处理下槲皮素的产量最高,异槲皮苷的产量相对稳定,山奈酚产量被遮荫影响较大,产量也相对最小。因此,在青钱柳的栽培过程中,强光更有利于促进黄酮含量与产量的积累。总之,光照条件的变化能够影响黄酮类化合物的积累模式与积累量。
综上所述,遮荫对青钱柳的苗高和地径生长、叶片黄酮类化合物积累均有显著影响。一层遮荫促进了青钱柳的苗高生长,二层遮荫均对苗高和地径产生了抑制作用,黄酮类化合物的含量与产量在两种遮荫处理下均受到了抑制。同时,青钱柳有一定的耐荫能力,受到弱光胁迫时能够通过外部形态、光合机构的成分与结构的可塑性变化来获取更多的光照、提高自己的光能捕获与利用能力,适应弱光环境。因此,研究青钱柳苗的生长与次生代谢物积累对光照的响应,对改善药用青钱柳人工林栽培技术和提高药用青钱柳产量具有重要意义。
猜你喜欢
农业技术与装备(2020年7期)2020-08-18 12:16:00
中国计算机报(2019年45期)2019-12-27 04:08:22
中成药(2017年9期)2017-12-19 13:34:21
中成药(2017年6期)2017-06-13 07:30:35
中成药(2017年4期)2017-05-17 06:09:31
山西大同大学学报(自然科学版)(2016年2期)2016-12-12 03:19:15
中国病理生理杂志(2015年8期)2015-12-21 12:38:08
电子世界(2015年19期)2015-03-27 12:11:22
浙江林业(2015年7期)2015-02-24 06:06:09
食品科学(2013年13期)2013-03-11 18:24:43
