
土壤动物对柠条灌丛枯落物分解和土壤碳含量的影响
2022-10-21 03:46吉雪茹刘任涛赵文智蒋嘉瑜常海涛
草业科学 2022年9期
吉雪茹,刘任涛,赵文智,蒋嘉瑜,常海涛
(1. 宁夏大学生态环境学院, 宁夏 银川 750021;2. 西北退化生态系统恢复与重建教育部重点实验室, 宁夏 银川 750021;3. 中国科学院西北生态环境资源研究院, 甘肃 兰州 730000)
柠条(Caragana korshinskii)为豆科锦鸡儿属灌丛植物,因其根系较为发达,有较强的抗旱性和适应性,同时具有很好的涵养水源、保持水土以及改良林地土壤等作用,在荒漠草原水土保持和固沙造林实践中被大面积栽植[1-2]。柠条植物枯落物作为半干旱区土壤碳含量的主要来源之一[3],通过分解向土壤中释放营养元素,维持植物正常生长,是连接地上地下生态系统物质循环与能量流动的重要生态界面[4]。枯落物分解过程中碳元素释放已成为柠条灌丛“肥岛”形成的重要来源和内在机制。
枯落物分解过程中的质量损失主要由环境因素和生物因素共同决定[5],因而枯落物质量损失和养分释放会随着微生境变化引起的环境和生物因素改变而发生显著变化。柠条灌丛冠幅大小直接影响枯落物积累量及分解过程[6],其引起的资源条件变化吸引土壤动物前来生存、定居,为极端环境条件下土壤动物存活提供了良好的避风港[7]。土壤动物是灌丛群落内物质循环过程中重要的消费者和特殊的分解者,其通过生长、繁殖等生命活动直接影响有机质的降解速率和营养元素的周转量,其次通过摄食和分泌排泄物等方式改变微环境,间接促进微生物的生长和对养分元素的利用,以此直接或间接促进灌丛微生境下枯落物分解[8],从而导致土壤碳收支和土壤肥力也随之变化[9]。因此,探究基于不同发育阶段的灌丛微生境枯落物分解对土壤碳的影响及土壤动物的分解功能,对于深入理解半干旱区灌丛的固碳作用机理及碳循环过程具有重要意义。
已有研究表明,土壤动物参与枯落物分解会促进土壤碳库含量的增加[10];刘瑞龙[11]通过研究土壤动物对枯落物质量损失和有机碳分解的影响,结果表明土壤动物对枯落物木质素、纤维素降解具有显著影响且加速了枯落物有机碳的分解;廖姝等[12]通过研究土壤动物对油樟(Cinnamomum longipaniculatum)、马尾松(Pinus massoniana)、麻栎(Quercus acutissima)、柳杉(Cryptomeria japonica)枯落物的影响,结果表明土壤动物显著影响枯落叶分解过程中可溶性碳、氮动态。徐国良等[13]通过13C 同位素标记技术证实跳虫(Folsomia candida)可以短时间内同化大量枯落物新碳,加快碳库存量的增长。但是,目前关于柠条灌丛微生境土壤动物参与的枯落物分解对土壤碳含量影响的研究较少,尤其是有关半干旱区灌丛生长过程中枯落物分解对土壤碳含量的影响及土壤动物分解功能的研究甚少。
本研究以宁夏盐池县荒漠草原柠条灌丛枯落物为研究对象,采用网孔分解袋法限制土壤动物进入,分析不同大小灌丛微生境下枯落物分解和土壤碳含量分布特征,结合土壤动物变化,阐明土壤动物参与的柠条枯落物分解对土壤碳含量的影响规律,旨在揭示柠条灌丛微生境土壤碳含量变化和枯落物分解及土壤动物生态功能的相互关系,为半干旱荒漠草原区植被建设及生态系统恢复提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于宁夏回族自治区吴忠市盐池县,县境由东南至西北为广阔的干草原和荒漠草原,地势南高北低,北接毛乌素沙地,南靠黄土高原,属典型的过渡地带。研究样地位于该县东北方向10 km处,地理坐标为107°27′30.18″ E,37°49′12.90″ N,海拔1 416 m。日照长,温差大,四季分明,属典型的温带大陆性季风气候;年均气温8.5 ℃,年降水量281.7 mm,其中全年72%的降水集中在7 月-9 月,年际变化率大;年蒸发量2 136 mm,年无霜期为165 d,年平均风速为4.89 m·s-1。土壤结构松散,肥力较低,质地多为沙壤土、轻壤土和沙土。主要植被类型为人工灌丛-草地镶嵌复合系统,包括柠条锦鸡儿柠条锦鸡儿(C. koushinskii)、油蒿(Artemisia ordosica)、牛枝子(Lespedeza bicolor)、花棒(Hedysarum scoparium)等灌丛及猪毛蒿(Artemisia scoparia)、苦豆子(Sophora alopecuroides)、中亚白草(Pennisetum centraticum)等草本植物。
1.2 研究方法
1.2.1试验设计
于2016 年10 月,在宁夏盐池县杨寨子村进行试验布设,设置不同大小柠条灌丛和不同体径的土壤动物类型两个处理因子。首先,按照灌丛大小(高度 × 冠幅)选取生长健硕旺盛的柠条锦鸡儿灌丛作为研究对象[6],分别为大型(A,341 cm × 223 cm)、中型(B,145 cm × 155 cm)、小型(C,519 cm × 122 cm)和极小型(D,115 cm × 71 cm);每种类型柠条灌丛选择3 株作为重复,并对样株进行标记。
其次,在研究样地内收集新鲜柠条锦鸡儿叶片(带叶柄),带回实验室自然风干。根据地表柠条锦鸡儿枯落物的分布特征及标准株下样方面积,分别称取6 g 柠条枯落物干样装入3 种分解袋中(15 cm ×15 cm),表面孔径依次为4 mm (允许所有土壤动物和微生物进入)、2 mm (仅允许中小型土壤动物和微生物进入)和0.01 mm (仅允许微生物进入),贴地面层网孔大小均为0.01 mm (避免枯落物损失和地下土壤动物的干扰)[14]。在分解袋上标好编码,并做记录。在4 类灌丛下分别布设3 种规格的分解袋,分解袋距灌丛根部< 15 cm;每两个分解袋之间相距10 cm;每株灌丛下均匀布设18 个(3 种规格 × 6 次取样)分解袋。
然后,用铁丝弯钩固定分解袋,埋藏深度为0-2 cm,使分解袋与土壤紧密接触。在2016 年12 月初完成分解袋布设,共布设分解袋216 个(即3 种规格 × 3 个重复 × 4 类灌丛 × 6 次取样)。于0 (2016 年12 月)、120 (2017 年4 月)、150 (2017 年5 月)、210(2017 年7 月)、510 (2018 年5 月)和570 d (2018 年7 月)分别取回枯落物样本进行分析处理。
1.2.2枯落物取样与分析
取样时,在每株灌丛下取3 袋样品,每次取出36 袋(3 种规格 × 3 个重复 × 4 类灌丛)。将取出的枯落物样品装入干净的信封袋,放入保温箱中带回实验室。
首先,用刷子清除枯落物表面附着的泥沙和杂质,置于烘箱(65 ℃,48 h),烘干至恒重,记录枯落物样品干重。然后,烘干后的枯落物样品用于枯落物养分含量的测定。其中,酸性洗涤纤维素(acid detergent fibers,ADF)采用改进的Van Soest 方法[15]测定;酸性洗涤木质素(acid detergent lignin,ADL)用范式法[15]测定;全氮采用元素分析仪(意大利DK6,UDK140分析仪)测定[16];全磷采用氢氧化钠熔融-钼锑抗比色法测定[16]。
柠条灌丛枯落物养分初始含量为:全磷0.22%,全氮2.12%,酸性洗涤纤维34.85%,酸性洗涤木质素9.66%。
1.2.3土壤取样与处理
每次取出分解袋的同时,用铁铲取出枯落物分解袋下0-10 cm 的土壤样品装入密封袋中,带回实验室自然风干,过2 mm筛,去除其中的杂质,用于测定土壤pH 和电导率、土壤粒径分析和土壤全碳含量(表1)。然后用铝盒取剖面土壤,用于土壤含水量的测定。

表1 4 类柠条灌丛生境的土壤初始理化性质Table 1 Initial physical and chemical properties of soil in four Caragana korshinskii shrub microhabitat types
土壤含水量采用烘干称重法测定(105 ℃,24 h)。土壤pH (2.5 ∶ 1 水土比浸提液)和电导率(5 ∶ 1 水土比浸提液)分别采用酸度计法和电极法进行测定。土壤粒径组成采用Mastersizer3000 激光衍射粒度分析仪进行其粒径体积百分含量的测定。土壤全碳通过元素分析仪(意大利DK6,UDK140 分析仪)来测定[16]。
1.3 数据处理与统计分析
1.3.1各孔径内枯落物质量损失率[17]

式中:W0为枯落物袋埋置前干枯落物质量;Wt为当次取样时各孔径内干枯落物残留量。
1.3.2土壤动物对枯落物分解的贡献率[18]

式中:Cf为全部土壤动物对枯落物分解的贡献率;Cma为大型土壤动物对枯落物分解的贡献率;Cme-mi为中小型土壤动物对枯落物分解的贡献率;Df为全部土壤动物引起的枯落物损失,即4 和0.01 mm 孔径中枯落物质量损失率之差;Dt为整个过程中生物因素与非生物因素引起的枯落物质量损失,即4 mm孔径枯落物袋损失率;Dma为大型土壤动物引起的枯落物损失,即4 和2 mm 孔径中枯落物质量损失率之差;Dme-mi为中小型土壤动物引起的枯落物损失,即2 和0.01 mm 孔径中枯落物质量损失率之差。
1.3.3数据处理
采用SPSS 20.0 软件进行数据统计与分析。采用单因素方差分析和多重比较法分析不同数据组间的差异。采用多因素方差分析网孔、分解时间和灌丛大小及其交互作用对土壤碳含量的影响。采用Pearson 相关系数分析不同指标间的相关性。显著水平为0.05。
2 结果与分析
2.1 土壤动物对柠条枯落物分解的平均贡献率
全部土壤动物参与枯落物分解时,B 类灌丛在分解至210 和570 d 时土壤动物贡献率表现为负效应(图1)。A 类和D 类灌丛在分解至570 d 时土壤动物贡献率均表现为负效应,在其他分解阶段均表现为正效应。C 类灌丛在整个分解阶段土壤动物贡献率均表现为正效应。
仅大型土壤动物参与枯落物分解时,A 类和D 类灌丛在分解至120 d 时土壤动物贡献率均表现为正效应,在其他分解阶段均表现为负效应(图1)。B 类灌丛在分解至120、150 和210 d 时土壤动物贡献率表现为正效应,在其他分解阶段均表现为负效应。C 类灌丛在分解至150 和210 d 时土壤动物贡献率表现为正效应,在其他分解阶段均表现为负效应。
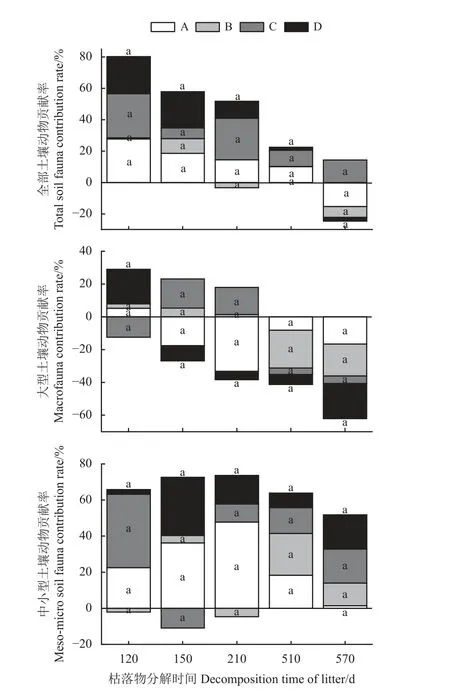
图1 不同微生境下不同体型土壤动物随枯落物分解的贡献率Figure 1 Contribution of different sizes of soil fauna to litter decomposition in different microhabitats
中小型土壤动物参与枯落物分解时,B 类灌丛在分解至120 和210 d 时土壤动物贡献率表现为负效应,在其他分解阶段均表现为正效应(图1)。C 类灌丛在分解至150 d 时土壤动物贡献率表现为负效应,在其他分解阶段表现为正效应。A 类和D 类灌丛在整个分解阶段土壤动物贡献率均表现为正效应。
在枯落物分解的整个过程中,4 类灌丛微生境土壤动物贡献率均无显著差异(P> 0.05)。
2.2 土壤动物参与柠条枯落物分解中土壤碳含量变化
柠条灌丛微生境土壤碳含量既受灌丛大小和分解时间的极显著影响(P< 0.001),亦受两者交互作用的显著影响(P< 0.05) (表2)。

表2 土壤动物参与柠条灌丛微生境枯落物分解过程中碳含量的方差分析Table 2 Analysis of variance between soil carbon content and experimental treatment
全部土壤动物参与枯落物分解时,当分解至120、210 和510 d,土壤碳含量表现为A 类灌丛显著高于C 类和D 类灌丛(P< 0.05) (图2)。但是,当分解至150 和570 d 时,土壤碳含量表现为4 类灌丛间均无显著差异(P> 0.05)。

图2 3 种网孔内不同大小灌丛下土壤碳含量随时间变化Figure 2 Changes in soil carbon content with time for three-mesh litter bags across different shrub sizes
仅大型土壤动物参与、仅中小型土壤动物参与以及仅有微生物参与枯落物分解时,土壤碳含量均表现为4 类柠条灌丛间无显著差异(P> 0.05) (图2)。
2.3 土壤碳含量与枯落物养分和土壤动物贡献率间的相关性
土壤碳含量与枯落物木质素含量呈极显著正相关关系(P< 0.01),但与土壤动物贡献率无显著相关性(P> 0.05) (表3)。

表3 土壤碳含量与枯落物养分和土壤动物贡献率间的相关性Table 3 Correlation coefficient between soil carbon content and litter nutrients and soil fauna contribution rate
3 讨论
3.1 柠条灌丛微生境土壤动物对枯落物分解的影响
土壤动物作为连接地上、地下物质循环和能量流动的纽带,对地上枯落物分解过程、结构和功能起着重要的反馈调控作用,是地下生态系统最活跃的部分[19]。本研究发现,全部土壤动物参与分解时,枯落物分解至120、150 和510 d,4 类柠条灌丛微生境土壤动物贡献率均为正值且无显著差异。这是因为该阶段柠条灌丛均处于非生长季,土壤冻融,气温回升,土壤动物休眠期结束,竞争压力小,且灌丛微生境下食物资源丰富,养分元素充足,吸引大量的土壤动物至灌丛下生存、繁衍,促进了枯落物的分解[20]。
但在枯落物分解至210 d,中型柠条灌丛微生境土壤动物贡献率为负值,其他3 类灌丛均为正值,且4 类灌丛间无显著差异。这是因为该时期柠条灌丛处于生长季,研究区降水较多,中型灌丛因冠幅较大,可以有效截留降水,从而抑制了枯落物的分解[21]。而其他3 类灌丛,枝叶相对稀疏,导致降水所产生的穿透雨量和树干茎流量加大,灌丛微生境下淋溶作用加强,从而加速枯落物的分解。但由于干旱区气候干旱少雨,所以降水的影响不足以引起灌丛间土壤动物贡献率有显著差异[22]。
当枯落物分解至570 d 时,小型灌丛土壤动物贡献率表现为正效应,而其他3 类灌丛均表现为负效应。这是因为小型灌丛处于生长初期,还没有建立起较为完整的食物网结构。随着灌丛增大,食物网逐渐完整和稳定,微生物的种类和数量增多,加速难分解物质的分解进程[23],从而导致土壤动物的作用相对减弱。
分解至510 和570 d 时,在仅有大型土壤动物参与下,土壤动物贡献率均为负值;在仅有中小型土壤动物参与时,土壤动物贡献率均为正值,即中小型土壤动物对枯落物分解的贡献率大于大型土壤动物贡献率。相关研究发现,中小型土壤动物为干旱区枯落物分解的主要参与者,其在分解过程中能够调节微生物种群、改造大型土壤动物粪便,并对枯落物分解起关键作用的微生物活性产生显著影响,从而间接影响枯落物的分解进程[24]。
3.2 土壤动物参与柠条枯落物分解对土壤碳含量的影响
本研究显示,全部土壤动物参与分解时,随柠条灌丛增大土壤碳含量相对增加,这与崔静等[25]、曲卫东等[26]的研究结果存在差异,可能是因为本研究灌丛间年限差异较小,时间周期短,不足以说明整个生长年限的变化。此外,枯落物分解至120 和510 d,土壤碳含量表现为小型和极小型灌丛低于大型和中型灌丛,而大型和中型灌丛下土壤碳含量差异较小。可能是由于小型和极小型灌丛处于生长初期,根系快速吸收养分和无机盐,间接改变了土壤的理化性质,影响了土壤动物和微生物的生长繁殖,消耗了地表大量有机质,导致两者微生境内枯落物分解所产生的土壤碳含量始终处于相对较低的水平。中型灌丛处于生长盛期,柠条灌丛枝叶茂盛,光合作用能力强,对土壤表层的碳元素吸收强度较小,致使地表有机质储量增多,碳含量升高[27]。大型灌丛处于老化期,生态功能逐步弱化,养分吸收的能力降低,导致地表土壤有机质的积累量相对较高[25]。当分解至570 d 时,4 类柠条灌丛下土壤碳含量均无显著差异。一方面是因为该时期是柠条灌丛的生长期,枯落物经过长时间的淋溶作用,可溶性物质持续减少;另一方面,土壤动物聚集,微生物酶活性提高,两者的共同作用可以加快土壤养分循环速率[28]。
当仅有大型土壤动物、仅有中小型土壤动物以及仅有微生物参与分解时,4 类灌丛下土壤碳含量间均无显著差异。且仅有大型土壤动物参与时,随分解时间的变化土壤碳含量呈不规则增加或减少,这是因为部分大型土壤动物通过取食、混合土壤及调节有机质等对土壤碳的固存和矿化产生影响。仅有中小型土壤动物参与时,随分解时间的变化大型和小型灌丛土壤碳含量均为正值,主要是因为中小型土壤动物在分解过程中的主要作用是调节微生物种群和分解大型土壤动物粪便[29]。并且,大型灌丛因生长年限较长,有更加完整的食物网和分解机制,小型灌丛因生长需要,物质循环加快,中小型土壤动物作用加强,因此两类灌丛下土壤碳含量均为正值。
本研究表明,土壤碳含量受枯落物袋网孔的影响较小,且与土壤动物贡献率相关性较小。这是因为枯落物分解的复杂性和野外环境的多变性等因素可能会引起本研究中土壤动物作用不明显[30],且土壤碳的形成和累积是一个长期的过程,短期枯落物的分解和土壤动物的作用并不能引起土壤碳含量发生显著变化。
4 结论
1)中小型土壤动物是枯落物分解的主要参与者,对枯落物分解的贡献率随分解时间变化正效应逐渐减弱,而大型土壤动物对枯落物分解的贡献率在510 d 后转变为负效应。
2)土壤碳含量受灌丛大小与分解时间交互作用的显著影响,且全部土壤动物参与枯落物分解在120、210 和510 d 时,土壤碳含量随灌丛大小的增加而增加。
3)土壤动物参与能显著提高枯落物的分解速率,但土壤碳的形成和累积是一个长期的过程,因而短期内枯落物分解和土壤动物作用并不能引起土壤碳含量发生显著变化。
猜你喜欢
农业与技术(2022年15期)2022-08-15
种子(2022年2期)2022-03-21
当代陕西(2019年7期)2019-04-25
湖南饲料(2018年2期)2018-12-05
领导文萃(2017年11期)2017-06-12
中国科技纵横(2016年15期)2016-12-29
高教探索(2015年10期)2015-10-29
绿色科技(2014年9期)2014-10-30
现代营销·经营版(2014年5期)2014-07-08
草原(2011年5期)2011-11-21
