
湖北省下三叠统牙形石生物地层序列与研究展望
2022-10-20 10:31崔亚圣杨良哲李红军余浩宇邹亚锐叶思遥
资源环境与工程 2022年5期
吴 奎, 崔亚圣, 刘 辉, 杨良哲, 李红军, 余浩宇, 邹亚锐, 叶思遥, 王 伟
(1.湖北省地质科学研究院,湖北 武汉 430034; 2.中国地质大学(武汉) 生物地质与环境地质国家重点实验室,湖北 武汉 430074;3.资源与生态环境地质湖北省重点实验室,湖北 武汉 430034; 4.远安县自然资源和规划局,湖北 宜昌 444200;5.合肥工业大学 资源与环境工程学院,安徽 合肥 230009)
湖北省地处华中地区,在构造单元上位于华南板块中北部边缘,其北部以秦岭—大别构造带与华北板块相接。在古、中生代之交,华南与华北板块之间尚有勉略洋相隔[1-2],勉略洋以南有扬子碳酸盐岩台地的存在(图1),使得湖北省在此阶段沉积了大量的海相地层[3-4](图2-A)。横贯东西的青峰—襄广断裂将湖北省分为两个地层区,其北为普遍接受了区域变质作用改造的秦岭区,其南为以沉积岩为主的扬子区(图2-B)。受到印支运动的影响,整个早三叠世时期湖北省范围内的海平面逐渐下降,从大冶组泥质灰岩、泥晶灰岩为主逐渐过渡到以嘉陵江组岩溶角砾岩、白云岩、白云质灰岩为主的沉积[4]。在早、中三叠世之交,随着印支运动的进一步启动,湖北省的海相沉积逐渐演变为巴东组滨海相碎屑岩[1-2]。近年来在南盘江及川西等地报道了一套富钾长英质火山灰,其富含黄绿色的豆状石英颗粒,因而被称为“绿豆岩”,往往被认为是下、中三叠统的岩石学标志,而这套“绿豆岩”的层位与当前早、中三叠世界线的标准化石Chiosellatimorensis的首现点接近[1,5]。最近,有学者在湖北省也发现了这套“绿豆岩”的存在[6]。
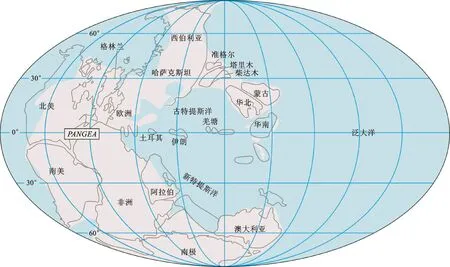
图1 古、中生代之交华南与华北板块相对位置图
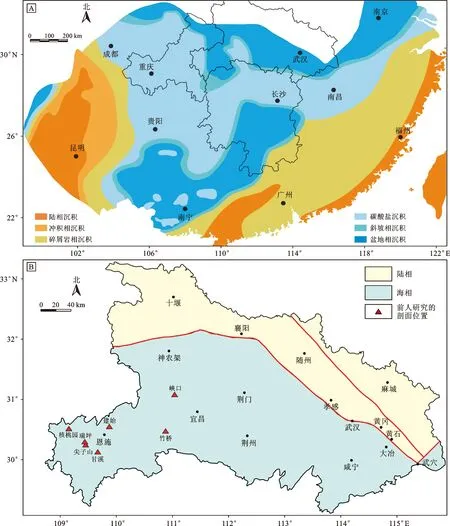
图2 华南早三叠世古地理图和湖北省早三叠世古地理图及前人研究剖面位置图
在二叠纪—三叠纪之交,发生了显生宙以来最大的生物大灭绝事件,造成了海洋中超过90%以上的物种消失[7-8]。这之后的海洋生态系统复苏过程,也是历次生物大灭绝发生以后最为漫长和复杂的[9-10]。期间反复出现的海水高温、缺氧、酸化等恶劣条件[11],导致全球范围内的海洋古生物的迟缓复苏[9];同时受到印支运动的影响,华南板块北部盆地逐渐变浅,成为岩溶角砾岩及白云岩发育的潮间带沉积,即古环境和古地理急剧变化的双重打击,使得该地区下三叠统中保存的古生物化石相对较少,这也体现在湖北省的下三叠统化石记录上。但是相较于其他地区,该段地层中依然有一些重要的化石保存,例如黄石地区的鱼化石,尤其是南漳—远安地区的海生爬行动物化石,更是属于全球独一无二的化石资源[12]。因此,湖北省广泛发育的下三叠统,是恢复二叠纪—三叠纪之交生物大灭绝之后海洋生态系统复苏进程的重要窗口之一。为了对这个过程进行详细的解译,就必须建立详细、统一的地层对比方案。作为古、中生代之交最重要的标准化石之一,牙形石(牙形刺)是建立该时期生物地层对比最重要的工具之一,可以为地层界线划分与对比、生物与环境协同演化等提供重要的信息[13]。
本文系统总结20世纪70年代以来前人在湖北省进行的下三叠统牙形石研究成果,为湖北省下三叠统牙形石的生物地层划分与对比提供系统性的意见,也提出一些值得进一步研究的相关问题。
1 湖北省下三叠统牙形石研究历史简介
相关三叠纪牙形石的研究报道最早出现于20世纪50年代[14]。中国关于三叠纪牙形石的研究始于1976年,王成源、王志浩首次报道了珠穆朗玛峰地区的三叠纪牙形石,为中国三叠纪牙形石研究打下了基础[15-16]。紧接着,湖北省下三叠统牙形石相关研究也陆续有了报道,拉开了湖北省早三叠世牙形石研究的序幕[16]。但之后的近20年里,湖北省地区下三叠统牙形石的研究进入了停滞状态。随着二叠纪—三叠纪之交层型剖面的建立,学者又将研究视线重新关注到二叠纪—三叠纪之交的生物大灭绝事件,尤其是本次生物灭绝事件之后的生物复苏过程。由于牙形石是这段地层中可以进行生物地层对比的最有效工具,湖北省下三叠统牙形石生物地层学的研究也重新被重视;另外,由于在建立二叠纪—三叠纪界线“金钉子”时,国内外学者对属于华南的上扬子、下扬子以及南盘江地区均做了一定的研究,而位于中扬子地区的湖北省被忽略,这也引起了学者对湖北省下三叠统牙形石研究的重视[17-18]。
2 湖北省早三叠世牙形石序列
当前,湖北省不同地区的下三叠统可以识别出9~12个牙形石带(包括亚带,如图3),包括(受到相变的影响,不同剖面的牙形石带并不完全一致,因此部分牙形石带的时间非严格按照自下而上顺序):Hindeodusparvus带、Isarcicellastaeschei带、Isarcicellaisarcica带、Hindeoduspostparvus带、Hindeodussosioensis带、Neoclarkinakrystyni带、Neoclarkinadiscreta带、Clarkinacarinata带、Sweetognathuskummeli带、Neospathodusdieneri带(包括Ns.dieneriM1亚带、Ns.dieneriM2亚带、Ns.dieneriM3亚带)、Neospathoduspakistanensis带、Novispathoduswaageni带(包括Nv.w.eowaageni亚带及Nv.w.waageni亚带)、Euryganthoduscostatus带、Parachiroganthus-Pachycladina带、Novispathoduspingdingshanensis带及Triassospathodushomeri带。
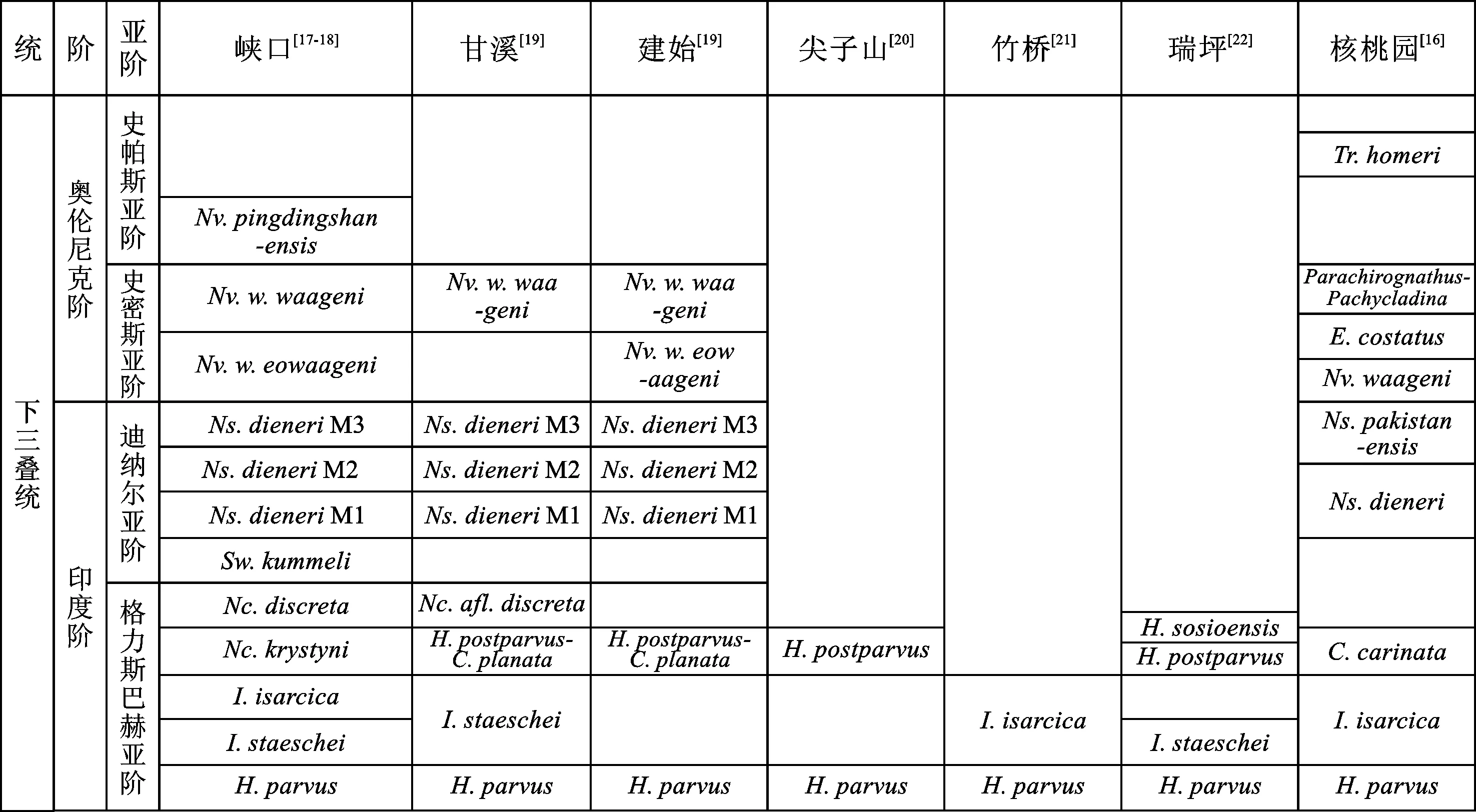
图3 湖北省不同剖面下三叠统牙形石带
2.1 Hindeodus parvus带
此牙形石带是国际范围内三叠系的第一个牙形石带,其在浙江长兴煤山D剖面的27层的底界标志着全球二叠系—三叠系界限标准[23]。在Hindeodusparvus的最前部位置发育有一个明显比其他细齿大2倍以上的主齿[24]。截至目前,在中国广泛分布的三叠纪海相沉积(包括湖北省全部海相下三叠统,如图1)、海陆过渡相沉积乃至全球范围内均有此牙形石带的报道[3,25]。值得注意的是,受到物种迁移速率、生物对环境的选择、化石保存概率等影响,不同剖面牙形石的首现点可能并不完全等时[12-18,23-27]。因此,有关二叠系—三叠系界线与H.parvus的首现点之间的关系,也吸引了许多学者的关注[3,28-29]。通过整体相关带对比表明,在华南范围内H.parvus最早出现的位置可能位于南盘江地区台地边缘向盆地过渡的边阳—明塘一带,而这也可能与当时该地区处于中层海水相区,受到了较小的缺氧与高温的影响有关[3,30]。
2.2 Isarcicella staeschei带
当前该牙形石带在全球众多地区都有一定的报道,包括克什米尔、阿尔卑斯南部、伊朗、阿曼、斯洛文尼亚、马来半岛以及中国的华南和西藏等地区[25],在湖北地区也有若干剖面有此牙形石带的报道(图3)。在Isarcicellastaeschei的基腔上方单侧,可见一明显垂直发育的细齿,这也是该种最明显的鉴定特征。在部分剖面,由于并未发现这一特殊的牙形石种,但可以识别出以与其形状类似却并未发育基腔单侧细齿的牙形石I.lobata建立的牙形石带,而这两个牙形石带也被认为是等时的[31-32]。在湖北地区,部分剖面也未识别出该牙形石带,这个现象可能与采样间距或者该牙形石属种对环境的选择有关(图3)。
2.3 Isarcicella isarcica带
当前,该牙形石带也有着较广泛的分布,包括巴基斯坦、克什米尔、印度、意大利、伊朗、斯洛文尼亚以及中国的华南地区[32]。在湖北省,此牙形石带也有报道(图3)。本带与Isarcicellastaeschei带相似,在部分剖面中有但并没有被发现,可能也是由于采样间距不够或者该牙形石属种的分布有着一定的局限性。
2.4 Hindeodus postparvus带
当前此牙形石带在斯洛文尼亚、印度以及中国的西藏和华南地区均有报道[20,32]。值得注意的是,Jiang et al.[33]曾经在位于南盘江碳酸盐岩台地相区的打讲剖面也发现了该牙形石种,但并未建立该牙形石带,而是在其首现点之上以Hindeodussosioensis的首现点建立了H.sosioensis带。
2.5 Hindeodus sosioensis带
该牙形石带由Kozur[34]首次在意大利西西里岛的Sosio峡谷地区建立,随后其也因Hindeodussosioensis和H.postparvus的共同出现而建立组合带[35]。在中国华南以及阿曼等地区,也有此牙形石带的相关研究报道[32]。
2.6 Clarkina carinata/Clarkina planata带
Clarkinacarinata首次在美国内华达地区的下三叠统中被报道,以齿体平台宽、细齿向上直立但较低矮、主齿两侧的平台不发育为特征。Orchard et al.[36]研究过C.carinata的系统发育特征,他们发现其自幼年期平台后边缘就不太发育,随着发育程度增加,齿体最后部的生长主要限制于主齿周围,因此呈现出尖锐的后部。Clarkinaplanata最突出的识别特征在于其具有一个宽阔而平坦的平台,最宽点位于中间,主齿位于相对圆的后边缘且其后方可见一窄的后边缘[37]。由于这两个牙形石种具有相近的地层分布特征[38],因此它们既常被单独用来建立牙形石带[25],也常被用来建立组合带[28]。在湖北省,这两个牙形石种均有相关报道(图3)。
2.7 Neoclarkina krystyni带
Neoclarkinakrystyni由Orchard和Krystyn[37]首先报道于印度Spiti地区格里斯巴赫亚阶的顶部。当前,该牙形石种亦被报道于加拿大、阿曼以及中国的湖北、贵州、浙江等地区[39]。Nc.krystyni最显著的鉴定特征为其平台相对较窄,后部末端尖,细齿高度逐渐增加,主齿明显[37]。在系统发育的过程中,该种的特征几乎保持稳定,只有少量变化,如细齿逐渐融合、后部细齿变得逐渐明显[39]。
2.8 Neoclarkina discreta带
此牙形石带最早由Orchard和Krystyn[37]在印度Spiti地区的Mikin组中报道,其属于早三叠世格里斯巴赫亚期晚期。最近的研究表明,在个体发育的过程中,Neoclarkinadiscreta在齿体中部位置两侧的凸起会逐渐变得明显,但是始终只占据极少部分位置,同时细齿的数量逐渐增多,并且逐渐更加融合在一起[39]。目前,该牙形石带在中国湖北峡口、浙江煤山、贵州甲戎、明塘以及印度的Spiti等地区均有报道[40]。
2.9 Sweetognathothus kummeli带
此牙形石带最早由Sweet[41]1970年报道于巴基斯坦盐岭地区,其属于早三叠世迪纳尔亚期的第一个牙形石带[25,39]。此牙形石种最显著的特征为整个齿体呈梳状,齿体长度约为其高的两倍,平台或侧脊极其狭窄[41]。最近,有学者通过大量的标本研究过Sweetognathoduskummeli的系统发育学特征和过程,他们发现该种整个生长阶段可以分为三个过程:早期细齿数量较少、主齿不明显,有时主齿后方还发育有其他细齿,最典型的特征为齿体不发育侧脊;中期细齿数量变多且更加融合在一起,主齿后方常发育有1~2枚细齿,最典型的特征为开始出现较为明显的侧脊;晚期细齿数量增加不明显,主齿后方可能发育3枚细齿,且主齿附近可能出现增生现象,呈近直角状,侧脊进一步加深,甚至可形成平台状[39]。当前,此牙形石带受到广泛报道,包括巴基斯坦、尼泊尔、印度(Spiti、Guryul Ravine)、美国(爱达荷、犹他、内华达)、加拿大以及中国华南和西藏等地区[25]。
2.10 Neospathodus dieneri带(包括Ns. dieneri M1亚带、Ns. dieneri M2亚带、Ns. dieneri M3亚带)
此牙形石种最早由Sweet[42]在1970年报道于巴基斯坦盐岭地区,随后在全球有大量的报道,包括印度的Spiti、克什米尔地区、美国西部的内华达和犹他地区、俄罗斯的Primorye南部地区、加拿大的极地和阿尔伯塔地区、日本的东南地区以及中国的藏南、广西、贵州、四川、湖北、江苏、安徽、浙江等地区[25]。根据中国安徽巢湖地区丰富的下三叠统牙形石分子,学者将Neospathodusdieneri细分为三个形态类型,即Ns.dieneriM1、Ns.dieneriM2和Ns.dieneriM3,这三个形态类型的主要鉴定特征类似,区别在于主齿相对于其后面相邻细齿的大小[43]。当前,在湖北省峡口、恩施地区及广西作登地区,已经有这三个亚带详细的报道[18-19,32]。最近,一项来自克什米尔地区的Guryul Ravine剖面的详细牙形石研究表明,该地区也有大量这三个形态种,但是由于获得的部分牙形石标本主齿和部分细齿残缺,因此只建立了后两个亚带[25]。
2.11 Neospathodus pakistanensis带
此牙形石带由Sweet[41]在1970年首次报道于巴基斯坦盐岭地区Mianwali组的Mittiwali段,处于Ns.cristagalli带和Nv.waageni带之间。当前,该牙形石带已经在世界范围内多次被报道,包括巴基斯坦、挪威斯瓦尔巴群岛、尼泊尔、加拿大极地和西部地区、美国西部、俄罗斯Primorye南部、澳大利亚西部、克什米尔、印度Spiti、越南、日本、中国华南和西藏等地区[25]。
2.12 Novispathodus waageni带(包括Nv. w. eowaageni亚带及Nv. w. waageni亚带)
此牙形石带由Sweet[41]于1970年首次在巴基斯坦地区报道。几十年来大量的相关研究表明,该牙形石种在世界范围内有广泛的分布,包括克什米尔、印度Spiti、阿曼、美国西部、加拿大极地地区、哥伦比亚、马来西亚、越南、澳大利亚、俄罗斯Primorye南部、日本以及中国的藏南、甘肃、广西、贵州、湖北、安徽、浙江、江苏、江西等地区[44]。在经过大量的样本研究后,Zhao et al.[43]于2007年将Novispathoduswaageni进一步划分为三个亚种,包括Nv.w.waageni、Nv.w.eowaageni及Nv.w.posterolongatus。Nv.w.waageni与Nv.w.eowaageni的不同之处在于,前者的齿体更厚,中部有加粗现象,细齿数量更少且弯曲,最高的细齿位于后部,基部轮廓更加弯曲且后部翘起明显[43-45]。由于Nv.w.posterolongatus具有明显较长的齿体和较多的细齿数量,现在学者更认同其属于Novispathodus属的一个种,即Novispathodusposterolongatus[25,43,45]。当前,Nv.w.waageni和Nv.w.eowaageni这两个亚带已经在澳大利亚西部、中国湖北大峡口及建始剖面和贵州甲戎剖面被识别[25]。目前,该种在中国安徽巢湖地区的平顶山西剖面的首现点,被认为是奥伦尼克阶底界的候选“金钉子”[46]。
2.13 Euryganthodus costatus带
此牙形石种由Staesche[47]于1964年首次在意大利Dolomites地区发现,随后在世界范围内许多地区均有报道,包括美国内华达、马来西亚、克什米尔、印度Spiti、日本、哥伦比亚、俄罗斯Primorye南部、阿拉伯、阿曼、意大利、克罗地亚、斯洛文尼亚、越南以及中国的甘肃、广西、云南、安徽巢湖、江苏、贵州、湖北、四川等地区[48]。系统发育学及生物演化学研究表明,Euryganthoduscostatus可以被划分出四个生长阶段:第一阶段该种发育有7~13个横脊状的细齿,细齿顶部呈尖圆状,高度近一致,侧视时某种程度上具有与Neospasthodus属或Sweetospathodus属类似的轮廓;第二阶段,横脊状的细齿数量比上一阶段略少,为5~12个,但可见细齿明显变宽,反口视时可见齿体后部更圆,基腔可延续到前部,细齿之间的横脊的高度降低,同时齿体轮廓由上一阶段的对称变得不对称;第三阶段,其横脊的数量变化不大,为7~12个,但除最中部横脊状细齿外其余横脊发育成角状;第四阶段,平台极度发育,甚至超过基部面积,横脊状的细齿几乎全发育成角状[48]。本属除Euryganthoduscostatus外,还有另外一特征明显的种,即整体形状、轮廓类似,但齿体上表面并不发育细齿或横纹的Euryganthodushamadai。相较于前者,Euryganthodushamadai的地理分布以及地层延限均较窄。最近来自甘肃益哇沟剖面的研究表明,Euryganthoduscostatus可以被作为识别早三叠世印度期—奥伦尼克期界线的重要化石,但是由于该种在高纬度地区以及冷水区并未有报道,因此并不能用其来取代Novispathoduswaageni的“金钉子”化石地位[49]。
2.14 Parachiroganthus-Pachycladina组合带
当前,此牙形石组合带的建带分子已经在低纬度地区的古特提斯洋、泛大洋以及美国西部地区有大量报道,中国范围内则包括四川、贵州、广西、秦岭以及西藏等地区[49-51]。在中国安徽巢湖地区,学者虽然没有在Nv.waageni带之上、Nv.pingdingshanensis带之下建立该组合带,但是该地区的平顶山西剖面也有这两个属的相关报道,并且从和龙山组顶部之下约1 m之处开始出现[52]。之前的研究认为,早三叠世这两个牙形石属主要分布于潮间带[50,53-54]。值得注意的是,在属于下斜坡相的贵州甲戎剖面,学者认为该剖面中此组合带的牙形石均为来自潮间带而后再沉积的分子[50]。
2.15 Novispathodus pingdingshanensis带
此牙形石带首次报道于华南板块的巢湖地区[43],现有研究表明该牙形石带在全球范围内也有较广泛的分布,包括美国西部、巴基斯坦盐岭、越南北部、加拿大西部、日本西南部、克什米尔、阿曼以及中国的贵州、广西、江苏、安徽等地区[25]。在详细研究平顶山西剖面和龙山组和南陵湖组的牙形石序列后,Liang et al.[52]提出建议,将Novispathoduspingdingshanensis作为史密斯亚阶—史帕斯亚阶界线的标准化石。
2.16 Triassospathodus homeri带
此牙形石种由Bender[55]在1967年首次报道于爱琴海地区的希俄斯岛,当前已经在世界范围内有广泛的报道,包括俄罗斯、日本、越南、泰国、马来西亚、克什米尔、印度Spiti、尼泊尔、新西兰、美国、加拿大、挪威斯瓦尔巴德、罗马尼亚、希腊、阿尔巴尼亚、斯诺文尼亚以及中国的安徽、江西、江苏、湖北、四川、青海、云南、贵州、广西、西藏等地区[32]。在建立该种的时候,Bender[55]并没有给出其关键的识别特征,只提到该种在其齿体后部1/3位置发育有椭圆形的基腔。在经历了几番厘定之后,该种现在比较被认可的鉴定特征为:齿体相对较长,前部细齿略倾斜,后部细齿倾斜程度增加且与基部呈45°角,下部轮廓的前方较平直,往后可见末端下折,基腔后半部分逐渐扩展,最宽点位于后半部分之前[56]。
3 湖北省早三叠世地层界线划分
根据当前国际年代地层表划分方案,早三叠世被分为两个期,分别为印度期和奥伦尼克期。其中,印度期和奥伦尼克期又进一步被划分为格里斯巴赫亚期、迪纳尔亚期及史密斯亚期、史帕斯亚期。根据上述牙形石带,可以将湖北省下三叠统划分出四个阶段。
在早三叠世,受到海平面逐渐降低的影响,湖北省的沉积地层主要以泥灰岩、灰岩、白云岩、白云质灰岩、角砾白云岩为主,省内岩性较为统一。根据《中国区域地质志湖北志》[57]总结,湖北省下三叠统主要为大冶组和嘉陵江组,这两套岩石地层单位以白云岩或角砾白云岩的出现为区分依据。
3.1 印度期
在湖北省下三叠统大冶组底部,普遍发育了与下伏上二叠统吴家坪组的含硅质团块的生物碎屑灰岩或大隆组的硅质岩、硅质泥岩整合接触的薄层状灰岩或页岩,可见大量的Clararia和Ophiceras化石。在这套代表了浅海陆棚盆地沉积环境的地层中,各剖面底部普遍可见若干层2~8 cm厚的白色凝灰质黏土层,以位于兴山县峡口镇的大峡口剖面和恩施市的甘溪剖面最为典型,这两个剖面二叠系—三叠系之交的火山灰均可达7层。详细的牙形石生物地层学研究表明,三叠系底界的“金钉子”化石Hindeodusparvus在大峡口剖面中大冶组最底部的中薄层状灰岩的中部开始出现[18],这也与三叠系底界界线层型剖面——煤山D剖面的表现一致,即Hindeodusparvus在煤山D剖面的首现点位于殷坑组最底部中薄层状灰岩的中部[23]。因此,在湖北省境下三叠统大冶组,二叠系—三叠系界线位于其最底部灰岩的中间。值得注意的是,在二叠纪—三叠纪之交,全球范围内许多地区都发育了一层厚度不一的微生物岩,在鄂西南以及东南地区也有这一套特殊的沉积物[58]。从岩性上来看,这类微生物岩呈厚层状,与下伏的吴家坪组岩性更加类似,与上覆的薄层状泥岩、泥灰岩、灰岩呈现出更截然的岩性变化。大量研究表明,三叠系底界的“金钉子”化石Hindeodusparvus在这一类微生物岩的下部、中部甚至最底部开始出现,表明这一套在生物大灭绝之后出现的微生物岩至少有相当一部分属于早三叠世最早期[20,59]。
当前,比较认可的格里斯巴赫亚期—迪纳尔亚期界线的划分标准是Sweetognathoduskummeli的首次出现[3]。根据这一划分标准,湖北省下三叠统格里斯巴赫亚阶—迪纳尔亚阶界线位于大峡口剖面大冶组底部之上约20 m处的薄层状灰岩中,属于大冶组第一段或第二段[18]。
3.2 奥伦尼克期
自20世纪90年代起,学者便开启了国际范围内奥伦尼克阶底界“金钉子”的研究工作,其中安徽省巢湖地区的平顶山西剖面更是被列为该界线的候选层型剖面[43,45-46]。近来,围绕该界线附近的牙形石研究表明,Novispathoduswaageni作为该界线的标准化石是最有优势的,同时作为特征明显的化石——Eurygnathoduscostatus也是识别该界线的重要化石辅助标志[44,48-49]。根据湖北省牙形石生物地层序列,基本可以将奥伦尼克阶的底界确定在大冶组三段的中薄层状灰岩中,在兴山大峡口剖面位于大冶组底部之上约35 m[18],在恩施建始剖面位于大冶组底部之上约40 m[49],在利川核桃园剖面位于大冶组之上约200 m[16]。
早三叠世是二叠纪—三叠纪之交生物大灭绝之后重要的生态系统恢复期,在生物逐渐复苏的同时,也发生了一些重要的次级灭绝事件,其中一次便发生在史密斯亚期—史帕斯亚期界线附近[11,60]。当前,比较认可的划分该界线的标准是牙形石Novispathoduspingdingshanensis的首次出现[50,52,61]。根据此划分标准,湖北省下三叠统史密斯亚阶—史帕斯亚阶的界线位于大冶组四段,在大峡口剖面则位于大冶组底部之上约40 m[18]。
4 结论
湖北省出露的下三叠统包括以灰岩为主体的大冶组和白云岩、角砾、灰岩为主体的嘉陵江组。其中,大冶组中可以识别出Hindeodusparvus带、Isarcicellastaeschei带、Isarcicellaisarcica带、Hindeoduspostparvus带、Hindeodussosioensis带、Neoclarkinakrystyni带、Neoclarkinadiscreta带、Clarkinacarinata带、Sweetognathuskummeli带、Neospathodusdieneri带(包括Ns.dieneriM1亚带、Ns.dieneriM2亚带、Ns.dieneriM3亚带)、Neospathoduspakistanensis带、Novispathoduswaageni带(包括Nv.w.eowaageni亚带及Nv.w.waageni亚带)、Euryganthoduscostatus带、Parachiroganthus-Pachycladina带、Novispathoduspingdingshanensis带,嘉陵江组中可以识别出Triassospathodushomeri带。湖北省大冶组跨越了早三叠世格里斯巴赫亚期、迪纳尔亚期、史密斯亚期以及部分史帕斯亚期,嘉陵江组主要沉积于史帕斯亚期。
猜你喜欢
中国科技纵横(2022年17期)2022-10-25
汉语世界(The World of Chinese)(2021年1期)2021-02-22
——工程地质勘察中,一种做交叉剖面的新方法
中国设备工程(2021年2期)2021-01-28
数学物理学报(2020年1期)2020-04-21
青海政报(2017年8期)2017-07-31
北京航空航天大学学报(2016年6期)2016-11-16
海峡姐妹(2016年7期)2016-02-27
学习月刊(2015年19期)2015-07-09
地震研究(2014年3期)2014-02-27
城市道桥与防洪(2013年8期)2013-03-11
