
1株狂犬病毒减毒株的毒力和分子特性研究
2022-10-19 06:18石磊泰李玉华俞永新
中国人兽共患病学报 2022年9期
石磊泰,邹 剑,李玉华,俞永新
进入21世纪后,狂犬病仍然是重要的公共卫生威胁,全世界每年因犬介导的狂犬病死亡的人数估计有59 000人,大多数死亡发生在亚洲(59.6%)和非洲(36.4%)。印度为当前狂犬病疫情最严重的国家,印度狂犬病死亡的人数在亚洲(占狂犬病死亡人数的59.9%)以及全世界均居首位(占狂犬病死亡人数的35%)[1-2]。第二位是中国,2007年狂犬病疫情高峰时,发病人数为3 300例。从2004年到2016年,狂犬病病死数量一直位居国内传染病死亡人数前三位。可见,我国狂犬病疫情形势严峻[3]。
狂犬病一旦发病,病程进展迅速,几乎100%死亡,潜伏期可以从数天到数年,目前为止尚无特效治疗药物,有效预防手段只有对动物和人开展疫苗接种。控制和消灭人类狂犬病的根本措施中,对犬类进行大规模免疫接种是最根本有效的措施。在西欧、中欧、加拿大和美国大部分地区应用减毒活疫苗通过诱饵投放的方式口服免疫野生动物,已成功消除当地的狐狂犬病并达到减少和控制人狂犬病的效果[4-6]。同时,世界卫生组织(WHO)极力鼓励研究和发展安全有效的犬用诱饵口服疫苗用于家犬,并已研究制定了现场应用口服犬用疫苗指南[7],一些高度减毒的减毒株如SAG2和VRG株已被WHO建议用于流浪犬的口服免疫[8-9],并已在很多国家广泛应用[10-12]。目前,狂犬病减毒活疫苗的研究仍在不断进展中,并已取得良好的结果,一些高度减毒、免疫原性良好的新减毒株已被筛选出来。
本实验室早在20世纪80年代就开始了口服狂犬病减毒活疫苗的研究,在早期阶段,培养了一种毒性较弱的减毒株CTN-181,该毒株对4周龄小鼠没有致病性,但对3周龄及以下小鼠仍有致病性[13],其遗传稳定性较差[14]。
为了进一步筛选出毒力更低、减毒特性更稳定、免疫原性更好的疫苗株,我们将CTN-181在豚鼠颌下腺中进行3次传代,然后通过噬斑进行纯化,获得1株对3周龄小鼠脑内接种无致病力、对乳鼠脑内致病力明显减弱、免疫原性良好、遗传稳定性高的减毒株,命名为CTN181-3[14]。
本文对CTN181-3株的表型和基因型特性及其遗传稳定性进行全面的研究,为开发一种可用于动物口服和人体肌内接种的减毒活疫苗提供科学依据。
1 材料与方法
1.1 毒种和细胞 狂犬病毒CTN181-3株为本文研究用毒株;CTN-1株为CTN-181株的亲本株,CTN-181株为CTN181-3的亲本株,均由本室筛选并保存[15-16];Vero细胞和BHK21细胞由ATCC提供,BSR细胞由巴斯德研究所提供。
1.2 实验动物 昆明乳鼠和小鼠,Hartley豚鼠,SPF级,由中国食品药品检定研究院动物繁育场提供;金黄地鼠由北京维通利华实验动物技术有限公司提供,动物实验均按我国动物福利要求完成。
1.3 病毒毒力测定 将CTN181-3病毒分别脑内接种于2周龄和3周龄小鼠,0.03 mL/只;口腔接种3周龄小鼠和8周龄金黄地鼠,0.1 mL/只,观察21 d,记录动物的发病死亡结果,同时以空斑法(PFU/mL)测定病毒滴度(PFU/mL)。
1.4 病毒空斑(PFU)滴定 在六孔板中制备单层BHK21细胞,将待测病毒稀释液稀释成10-1~10-7系列,每个稀释度做复孔,每孔加0.2 mL;用病毒稀释液作空白对照。轻轻摇匀,置于5% CO2、37 ℃培养箱中,吸附1 h后每孔加甲基纤维素 4 mL 覆盖,在 5% CO2、35 ℃培养箱中继续培养,7 d后弃掉覆盖物,每孔加入2 mL结晶紫染料,染色15 min后计数,计算病毒的噬斑形成单位(PFU/mL)。
1.5 CTN181-3株的传代稳定性
1.5.1 乳鼠脑内传代毒力稳定性 将CTN181-3株脑内接种1~3 d龄乳鼠,待乳鼠发病濒死时无菌取脑,研磨后制成病毒悬液,脑内接种相同日龄乳鼠连续传5代,分别测定每代病毒材料的lgPFU和lgLD50,LD50测定采用3周龄小鼠脑内接种法,0.03 mL/只。
1.5.2 豚鼠颌下腺传代毒力稳定性 将CTN181-3株颈部皮下注射2只豚鼠,每只接种1 mL,注射后2 d收取豚鼠颌下腺,研磨制成病毒悬液,病毒悬液在BHK21细胞上增殖一代后以同法接种豚鼠传下一代,共在豚鼠颌下腺连续传4代,分别测定每代病毒材料的lgPFU和lgLD50,LD50测定采用3周龄小鼠脑内接种法,0.03 mL/只。
1.5.3 细胞传代后毒力稳定性 将CTN181-3株分别在BSR细胞和Vero细胞上连续传10代,用3周龄小鼠对传代后毒株进行脑内(0.03 mL/只)和肌内(0.1 mL/只)毒力测定。
1.6 病毒全基因序列测定
1.6.1 引物设计及合成 根据GenBank中登录号为FJ959397的CTN-1株设计8对引物,用于CTN181-3株全基因组分段扩增和序列测定,见表1。

表1 狂犬病病毒CTN克隆株全基因组分段扩增所用引物Tab.1 Primers used for CTN complete genome amplification
1.6.2 病毒RNA的提取及病毒基因组的分段PCR扩增 用QIAamp病毒RNA提取狂犬病病毒CTN181-3的基因组RNA。使用GoScript逆转录系统将RNA逆转录为cDNA,使用表1中的引物进行PCR分段扩增。反应体系共50 μL:10 μL 5×Phusion HF缓冲液、1 μL 10 mmol/L dNTP、2.5 μL上游和下游引物、1.5 μL DMSO、0.5 μL DNA聚合酶、2 μL cDNA和去离子水。反应条件:在98 ℃下初始变性30 s;98 ℃持续10 s,53 ℃持续30 s,72 ℃持续90 s,总共30个循环;在72 ℃下延伸5 min。PCR产物用1%琼脂糖凝胶电泳进行鉴定,通过QIA快速凝胶提取试剂盒进行纯化和回收。
1.6.3 PCR扩增产物的克隆、序列测定及序列拼接与分析 将PCR纯化产物与pGEM-T载体系统连接后转化感受态DH5α,挑取阳性克隆,菌落PCR鉴定无误后测序。每个片段至少应送3个阳性克隆进行双向测序验证。测序结果通过DNAStar和MegAlign软件进行拼接分析。
2 结 果
2.1 CTN181-3株对实验动物的毒力 以5.10 lgPFU/只的高滴度病毒脑内接种3周龄小鼠均无致病性;对2周龄小鼠的致病性亦明显降低,如以4.10 lgPFU/只病毒接种时,只有20%小鼠死亡。以5.60 lgPFU/只的病毒量口腔感染3周龄小鼠和8周龄地鼠,观察21 d均无动物出现发病死亡,见表2。

表2 CTN181-3对小鼠和金黄地鼠的毒力Tab.2 Pathogenicity of CTN181-3 in mice and golden hamsters
结果显示,CTN181-3株对3周龄小鼠脑内接种无致病性,对2周龄小鼠脑内接种呈现低毒力;口腔感染3周龄小鼠和8周龄金黄地鼠后实验动物无发病死亡。
2.2 CTN181-3的传代稳定性
2.2.1 在乳鼠脑内和豚鼠颌下腺传代后的毒力稳定性 CTN181-3通过乳鼠脑内连续传5代,病毒滴度可达7.10 lgPFU/mL,第1、3、5代乳鼠脑悬液对3周龄小鼠脑内接种未见发病。另通过豚鼠颌下腺连续传4代,病毒滴度最高可达7.63 lgPFU/mL,分别用4个代次豚鼠颌下腺病毒悬液接种到3周龄小鼠的脑内,没有发病,这表明CTN181-3保持了减毒病毒的稳定性,并且没有出现减毒特性的回复,结果如表3所示。
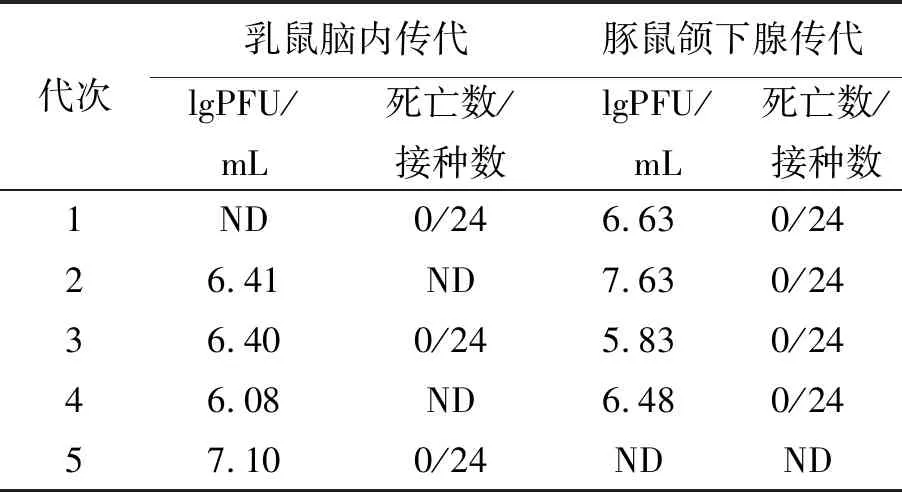
表3 CTN181-3在乳鼠脑内和豚鼠颌下腺培养传代后病毒的毒力稳定性Tab.3 Neroattenuation stability of CTN181-3 after serial passages in brains of suckling mice and submandibular glands of guinea pigs
2.2.2 细胞传代毒力和基因稳定性 CTN181-3株在BSR细胞和Vero细胞上各传10代后,分别得到CTN181-3 B10和CTN181-3 V10,两传代株病毒的神经毒力经3周龄小鼠脑内和肌内测试,结果小鼠均未发病和死亡,见表4。结果显示,CTN181-3株经乳鼠脑内、豚鼠颌下腺和两种细胞培养多代后,未见病毒毒力返祖,说明CTN181-3株遗传稳定性为十分稳定。

表4 CTN181-3 B10株和V10株对3周龄小鼠的致病性Tab.4 Pathogenicity of the B10 strain and V10 strain in 21-day-old mice
2.3 CTN181-3株基因型特性及其减毒基因的分析
2.3.1 CTN181-3的基因特征 CTN181-3株的基因全序列长度为11 924 bp,已被纳入GenBank,登录号为KU946961。其基因组组成与GenBank上发表的其他狂犬病病毒基因组一致。从3′端到5′端,先导RNA、5个蛋白质结构基因(N、P、M、G、L)和非编码尾部结构依次排列。其中3′先导区包括59个核苷酸,5′非编码区包括71个核苷酸。各结构基因皆包括3′端AACA和5′端的polyA尾结构。N基因位于 60~1 484 nt,ORF为72~1 424 nt,编码450个氨基酸;P基因位于1 487~2 475 nt,ORF为1 516~2 409 nt,编码297个氨基酸;M基因位于2 481~3 283 nt,ORF为 2 496~3 104 nt,编码202个氨基酸;G基因位于3 289~5 355 nt,ORF为3 316~4 890 nt,编码524个氨基酸;L基因位于5 380~11 853 nt,ORF为5 407~11 793 nt,编码2 128个氨基酸。各结构基因间隔区分别为2、5、5、24 个核苷酸。
2.3.2 基因同源性比较 从GenBank挑选国内外已公开发表的狂犬病疫苗株和街毒株的基因组序列与我国的狂犬病毒减毒株进行全基因组的同源性分析。
狂犬病毒减毒株CTN181-3与国内外疫苗株和街毒株的全基因组同源性在81.4%~93.3%,与国内4个分离株HN10、BD06、DRV和MRV的同源性为82.9%~93.3%,而与国外分离株的同源性仅为81.4%~84.4%。见表5。说明CTN181-3株与国内分离株在遗传进化过程中亲缘关系较近;而与国外其他疫苗株和街毒株的同源性则较低,亲缘进化关系较远。

表5 CTN181-3与其他株狂犬病病毒全长核苷酸和氨基酸同源性比较(%)Tab.5 Comparison of full-length-sequence nucleotide and amino acid similarity among CTN181-3 and other rabies virus strains (%)
2.3.3 CTN181-3的减毒基因分析 与致病性较高的CTN-1株相比,CTN181-3株在P基因5′端的mRNA polyA尾终止信号A7区段有1个A残基的丢失。CTN181-3与CTN-1的同源性为99.8%,与CTN-181的同源性为99.9%。CTN181-3与其亲本株CTN-181、CTN-1的所有核苷酸和氨基酸差异位点见表6。

表6 CTN-1、CTN-181和CTN181-3核苷酸和氨基酸位点差异Tab.6 Summary of nucleotide and amino acid differences among CTN-1,CTN-181 and CTN181-3
结果显示,CTN181较其母株CTN-1发生了8个氨基酸位点(N、M、G、L区各发生1、1、4、2个)的突变,其中[G276 L(TTG)→V(GTG)和L1496 M(ATG)→W(TGG)]2位氨基酸突变在CTN181株未发生,为CTN181-3株特有。由此我们认为,CTN181-3的毒力高度减弱、遗传稳定性高度稳定的分子基础为上述8个位点氨基酸突变的共同作用,而G276和L1496位氨基酸突变则导致CTN181-3株的毒力进一步降低。
3 讨 论
本文对实验室培育筛选到的CTN181-3减毒株的致病性、分子特性和遗传稳定性进行了系统研究。CTN181-3株对实验室狂犬病敏感动物无论是脑内还是口腔感染均无致病性,在同等病毒量的前提下,第1代病毒CTN-1株对4周龄及以上小鼠有致病性;第2代病毒CTN-181株对4周龄小鼠无致病性,但对4周龄以下小鼠有致病性[13];而第3代病毒CTN181-3株对3周龄小鼠无致病性;三代病毒对小鼠的致病性呈下降趋势,毒力逐代降低减弱。毒力稳定性方面,CTN181-3株病毒通过乳鼠脑内、豚鼠颌下腺、不同的细胞株多代传代,病毒的致病性没有发生改变,减毒基因也保持不变,均无回复突变发生。因此,用CTN181-3株制备口服活疫苗可以避免病毒对外传播引起任何动物致病的危险性。以上结果达到了WHO和国外用于野生动物和犬口服疫苗的基本要求,如在全球广泛应用的SAG2株,以6.5 lgPFU的剂量脑内注射6周龄成年小鼠不致病,乳鼠脑内回传5代毒力未返祖[10]。
与国内外疫苗株和街毒株进行全基因组同源性对比分析,狂犬病毒减毒株CTN181-3株与国内外疫苗株和街毒株的全基因组同源性在81.4%~93.3%,与国内4个分离株HN10、BD06、DRV和MRV的同源性为82.9%~93.3%,而与国外分离株的同源性仅为81.4%~84.4%。说明CTN181-3株与国内分离株在遗传进化过程中亲缘关系较近;而与国外其他疫苗株和街毒株的同源性则较低,亲缘进化关系较远。
从CTN181-3株与其亲本株CTN181以及CTN-1株[17-18]的全基因序列分析结果来看,与狂犬病毒毒力密切相关的G333位点[19],已经由CTN-1株的精氨酸变为CTN181株和CTN181-3株的谷氨酰胺,毒力已经大幅降低;而与毒力相关的另一位点G194[20-21],无论是CTN-1株、CTN181株还是CTN181-3株,该位点均为天冬酰胺,而不是会导致毒力升高的赖氨酸。就CTN181-3株而言,这2个毒力关键位点已经显示出低毒力的分子特征。
与CTN181株相比,CTN181-3株有4个氨基酸位点发生变化,分别是G266位点由N(AAT)变为D(GAT),G276位点由L(TTG)变为V(GTG),G299位点由K(AAA)变为R(AGA),L1496位点由M(ATG)变为W(TGG)。其中G276(V)和L1496(W)为CTN181-3株特有。CTN181-3株除保留CTN181株的减毒基因外,另发生2个氨基酸的突变。因此,可认为这2个氨基酸是CTN181-3株毒力进一步减弱的分子基础。
综上所述,CTN181-3株的神经毒力高度减弱,毒力十分稳定。其实验室指标与被WHO认可并推广应用近30年的口服疫苗株SAG2相当[10],是一株良好的狂犬病口服活疫苗候选株,目前对犬和其他大型动物的实验以及田间试验正在计划之中。
利益冲突:无
引用本文格式:石磊泰,邹剑,李玉华,等.1株狂犬病毒减毒株的毒力和分子特性研究[J].中国人兽共患病学报,2022,38(9):790-795,801.DOI:10.3969/j.issn.1002-2694.2022.00.114
猜你喜欢
浙江临床医学(2022年8期)2022-11-22
畜牧兽医学报(2022年7期)2022-08-05
中国兽医学报(2022年4期)2022-06-17
天津农业科学(2022年5期)2022-05-31
江苏农业科学(2019年14期)2019-09-23
农家致富顾问·上半月(2019年7期)2019-09-10
阅读(低年级)(2019年6期)2019-08-27
农家科技下旬刊(2017年7期)2017-08-22
山东农业科学(2016年4期)2016-11-19
读写算·小学低年级(2016年6期)2016-05-14
