
濒危植物血皮槭花部特征和交配系统*
2022-10-15 08:36孙佳伟郑勇奇于雪丹夏新合赵玉贤武宇霞张川红
林业科学 2022年6期
孙佳伟 郑勇奇 于雪丹 夏新合 赵玉贤 武宇霞 张川红
(林木遗传育种国家重点实验室 国家林业和草原局林木培育重点实验室 中国林业科学研究院林业研究所 北京 100091)
20世纪80年代以来,学者们从遗传多样性、种群生态学、生殖生物学等多个角度对植物濒危机制进行了探讨(Fiedleretal., 1992a; 1992b),普遍认为物种濒危是多因素综合作用的结果,其中许多植物因生殖力、存活力和适应力低下等内在因素导致濒危。有性生殖对植物生命延续和种群数量扩大具有决定性作用,该过程中任何一个环节出现障碍,都会导致生殖失败,如濒危植物羊角槭(Aceryangjuechi)存在胚珠败育现象(许小连,2012),风箱果(Physocarpusamurensis)(殷东生等,2016)和夏蜡梅(Sinocalycanthuschinensis)(Lietal., 2020)等因自然条件下传粉媒介种类和数量少、访花频率极低等因素导致结果率或结实率低,进而引起物种濒危。
血皮槭(Acergriseum),无患子科(Sapindaceae)槭属(Acer)落叶乔木(Harringtonetal., 2005),是我国特有种、世界濒危种,集中分布于我国中部地区,包括湖南、湖北、河南、山西、陕西、四川、重庆和甘肃等省(市),生长于海拔1 500~2 000 m疏林中,其天然群体数量少,呈片段化分布(Gibbsetal., 2009; 陈朋等,2013)。血皮槭树干呈赭褐色,树皮纸状卷曲剥落,秋季叶色红艳,是世界著名彩干兼彩叶树种,可作为庭园主景树(van Gelderenetal., 1994)。1993年血皮槭曾获得英国皇家园艺学会园艺奖,2013年被列入《北京市主要常规造林树种目录》(乔红娟,2018),同时也被收录于国家林业和草原局发布的《中国主要栽培珍贵树种参考名录(2017版)》中。在国内血皮槭苗木市场需求量大,但由于其为濒危种,且濒危原因尚不清楚(汪松等,2004),故目前市场上苗木稀缺,供需矛盾突出。孙圣(2014)研究认为血皮槭天然群体遗传多样性水平较高,认为遗传因素不是导致其濒危的主要原因。陈朋等(2013)实地调查发现,血皮槭野外植株结实率低,天然更新困难,实生幼苗生境恶劣,种间竞争激烈,有些群体表现为衰退趋势(叶学敏,2017),上述因素可能是导致其濒危的原因。对血皮槭天然群体和人工栽培植株均存在结实率低的原因,目前尚无相关文献报道。早年国外学者对植物园内种植的少量血皮槭性别观察有争议,有学者认为是雌雄同株,有学者认为是雌雄异株,美国学者de Jong(1976)将血皮槭花划分为雌花、雄花Ⅰ型和雄花Ⅱ型。血皮槭结实率低是否与花部特征和交配系统有密切关系尚需研究。鉴于此,本研究以血皮槭人工栽培植株为材料,探究其花部特征和交配系统类型,从生殖生物学角度寻找血皮槭结实率低的原因,以期为缓解目前苗木供需矛盾、制定有效措施奠定理论基础。
1 研究区概况
研究区位于北京市中国林业科学研究院内(40°0′34″,116°15′4″),平均海拔43.5 m,典型北温带半湿润大陆性季风气候,夏季高温多雨,冬季寒冷干燥,春、秋短促。全年无霜期180~200天,年均气温约12.9 ℃。降水季节分配不均匀,全年降水的80%集中在夏季6、7、8月。
2 研究与方法
2.1 试验材料
以血皮槭为实生苗人工栽培植株为试验材料,共计8株开花母树,树龄15年,平均胸径9.6 cm,平均树高5.1 m,无病虫害,生长状况良好,已进入正常开花结实年龄。
2.2 试验方法
2.2.1 开花动态和花形态特征观察 分别于2019和2021年4月观测血皮槭植株开花情况。参考Dafni(1992)标准,开花物候划分以下乎个阶段: 1)始花期——第一个植株开花的日期; 2)盛花期——50%以上个体开花,群体进入盛花期; 3)末花期——75%以上个体已经谢花,视为进入末花期,直至花全部凋零(最后一朵花凋落)视为花期结束。雌花成熟期指柱头开始具有可授性到失去可授性的时期,雄花成熟期指从花粉开始释放到释放完毕的时期。统计各物候期的持续时间,同时在血皮槭盛花期,于开花树体上、中、下3层,东、南、西、北4个方向采集花枝,统计花序类型以及雌雄花数量和比例。
随机选取健康的花蕾,挂牌标记,每天上午9∶00观察并拍照以记录开花特征和开放进程。同时采集完全开放的雌花和雄花分别进行解剖观测,采用游标卡尺(精度0.1 mm)测量其花纵径、花横径、花萼及花瓣数量、雄蕊数量及长度(花丝基部到花药顶端)等形态指标,雌花和雄花各测量60朵(60次重复)。通过便携式显微镜(SCALAR)拍照记录花形态。
2.2.2 花粉生活力和柱头可授性检测 采用离体萌发法检测花粉生活力。参照王续蕾(2016)方法配制含有蔗糖(10%)和硼酸(0.03%)的液体培养基。标记雄花若干,在花药开裂前,采集花药于硫酸纸袋中带回,室温干燥2 h后收集花粉并开始计时,分别于0、3、6、12、24 h取适量花粉进行生活力检测,每处理3次重复,探究花粉生活力的动态变化。蘸取少量花粉于凹面载玻片上,滴入适量培养基搅拌均匀,置于培养箱(GXZ光照培养箱)内15 ℃、黑暗条件下培养6 h,盖上盖玻片,于光学显微镜(Leica DM 2500,10×下观察3~5个视野,统计每个视野全部花粉数和萌发的花粉数。以花粉管伸长长度大于花粉粒直径视为花粉萌发,花粉生活力采用下式计算: 花粉生活力(%)=萌发花粉数/全部花粉数×100%。
采用过氧化氢法(Dafnietal., 1998)检测柱头可授性。根据花的不同发育阶段选取柱头各10枚,浸入反应液(3%H2O2)中,置于体视镜(Leica S8AP0)下观察,反应越剧烈,气泡数量越多,表明柱头可授性越强。
2.2.3 花粉胚珠比和异交指数估算 采取即将开放而花药尚未开裂的雄花10朵,每朵花取全部花药,用镊子挤碎,放入盛有适量HCl(1 mol·L-1)溶液的玻璃瓶中,置于60 ℃水浴锅内水解1 h去除花药壁,蒸馏水定容制成5 mL花粉粒悬浮液,用移液枪吸取5 μL滴在凹面载玻片上,置于光学显微镜下(10倍左右)观察统计花粉数量,重复取样观察3次,计算单花花粉量和平均单花花粉量。取雌花子房在解剖镜下用解剖针划开心皮,观测并记录其胚珠数,根据单株雌雄花比例分配权重,用单花花粉量除以单花总胚珠数计算花粉胚珠比(pollen-ovuleratio,P/O)。
参照Dafni(1992)标准进行交配系统评估。1) 花朵直径<1.0 mm记为0; 1.0~2.0 mm记为1; 2.1~6.0 mm记为2; >6.0 mm记为3。2) 花药开裂时间与柱头可授期之间的时间间隔: 同时或雌蕊先熟记为0; 雄蕊先熟记为1。3) 柱头与花药的空间位置: 同一高度记为0; 空间分离记为1。三者之和为异交指数(out crossing index,OCI),评判标准为: OCI=0,为闭花受精; OCI=1,为专性自交; OCI=2,为兼性自交; OCI=3,为自交亲和,有时需要传粉者; OCI≥4,为部分自交亲和,异交需要传粉者。
2.2.4 人工控制授粉试验 为检测交配系统类型,随机选取发育状况良好的花朵分别进行以下5种处理: 1)自然对照组——不套袋,自由传粉; 2)同株异花授粉——套纸袋,采用同株异花花粉进行人工授粉; 3)异株花授粉——套纸袋,采用不同单株花粉进行人工授粉; 4)开花前套纸袋,不授粉,检测是否存在单性结实现象; 5)套网袋,检测有无风媒传粉。每处理不少于60朵花,于60天后检测统计结实状况。具种仁可育的种子称为饱满种子。结果率和结实率计算公式如下:
结果率(%)=(翅果数量÷心皮数量)×100%;
结实率(%)=(具饱满种子的翅果数量)÷心皮数量×100%。
3 结果与分析
3.1 花形态特征与雌雄花比例
3.1.1 花形态特征 血皮槭花性别表达复杂,花基于功能可分为雄花和雌花2类,其中雄花根据雌蕊退化程度又分为雄花Ⅰ型和雄花Ⅱ型。雄花Ⅰ型仅具有雄蕊(图1A),雌蕊退化消失,花药正常开裂散粉,形态与功能表达性别一致。雄花Ⅱ型(图1B)具有雄蕊和退化雌蕊,退化雌蕊无子房仅有2裂柱头,雄蕊长于雌蕊,花药正常开裂散粉,为功能雄性。雄花Ⅰ型和Ⅱ型大小没有差别,但Ⅰ型较常见,Ⅱ型较少见。雌花(图1C)形态完整,具有雌蕊和雄蕊,雌蕊柱头2裂,柱头高于花药,仅基部联合,子房上位,密被白色绒毛; 雄蕊退化,花丝较短,花药从雌花开放至结束始终不开裂散粉,雌花为假两性花,功能雌性。雌花和雄花均具有结构相似的蜜腺,呈肉质环状,位于花瓣与雌雄蕊之间(图1)。从单花形态比较发现,雌花雄蕊长度小于雄花Ⅱ型,雌花柱头高度明显高于雄花Ⅱ型(花柱异型),即雄花和雌花的雌蕊与雄蕊长度不等长,单花存在雌雄异位现象。另外,雌雄异花,也是雌雄异位的表现。

图1 血皮槭花形态Fig. 1 Morphology of floret in A. griseumA: 雄花Ⅰ型(fi: 花丝, an: 花药, ne: 蜜腺); B: 雄花Ⅱ型(pi: 雌蕊); C: 雌花(ov: 子房).A: Male floret Ⅰ(fi: filament, an: anther, ne: nectary); B: Male floret Ⅱ(pi: pistil); C: Female floret(ov: ovary).
由表1可知,雌花花高、花直径皆大于雄花,二者花萼、花瓣数量和雄蕊数量无差异,皆为5或6枚。雌花花萼和花瓣数量多为5枚,少数为6枚; 花萼和花瓣为5或6枚时,对应雄蕊数量为10或12枚。
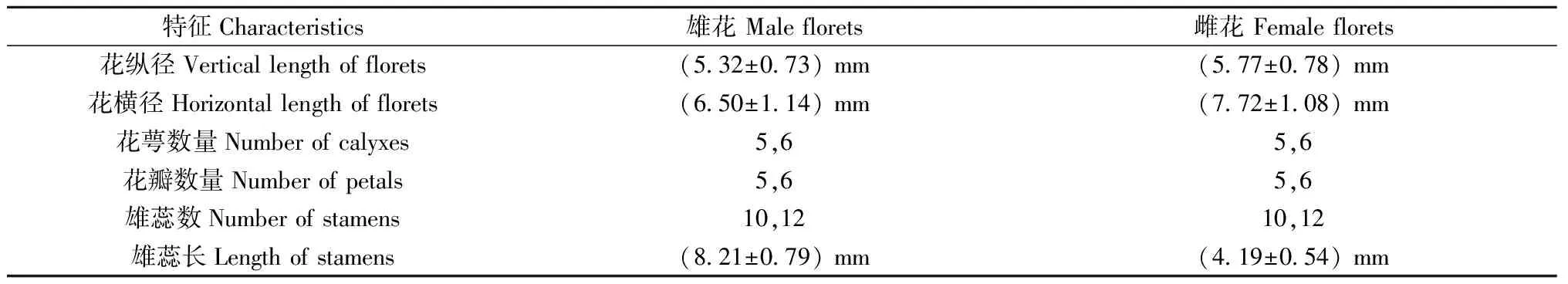
表1 血皮槭花形态特征(平均值)①Tab.1 Morphological characteristics of florets in A. griseum (average)
3.1.2 花序类型和雌雄花比例 血皮槭花芽为混合芽,多着生于小枝顶端。雄花与雌花一般共生于1个聚伞花序,常由3小花组成,每个花序着生于2枚复叶之间。不同花序中花性别和数量不同,可将发现的花序划分为3雄型(3)、3雌型(3♀)、2雄1雌型(21♀)、1雄2雌型(12♀)和1雄1雌型(11♀)。由表2可知,3小花型花序数量最多,占98.63%,其余类型花序仅占1.37%。
血皮槭为雌雄异花同株。从单株来看,雌花百分比平均78.77%,雄花21.23%,单株雌雄花比例平均约3.71∶1(表2),血皮槭单株雌花数量接近4倍于雄花。本研究中8株样株共观察花数量2 402个,平均每株300.25个,其中雌花数量1 853个,占总数量为77.14%,雄花数量549个,占总数量为22.86%,整体雌雄花比例约3.37∶1。
3.2 花开放动态与花物候
3.2.1 花开放动态 从花开放过程来看,雄花和雌花均经历以下5个阶段。 雄花:1)花蕾期,花芽膨大,发育为椭球状花蕾(图2A);2)萌动期,花蕾进一步发育膨大,花撑开芽鳞片(图2B);3)露白期,花药露出(图2C);4)开放期,花梗伸长,花突破芽鳞片,雄花花丝明显伸长,花药伸出花瓣,随后花药开裂散粉; 同一花序内雄花散粉不同步,位于中间的雄花先散粉,两侧雄花后散粉; 同一朵雄花内花药开放也不同步(图2D-F);5)凋零期,花药散粉完毕后连同花丝、花瓣、花梗迅速干枯、脱落(图2G-H)。雄花从开放到脱落平均耗时7.6天。雌花: 花蕾期萌动期同雄花(图3A-B);露白期,雌花柱头露出,已张开且轻微反卷,具有可授性(图3C);开放期,花梗伸长,花序露出,花瓣张开幅度小,随着子房膨大,花瓣被逐渐撑开,同时随着开放柱头卷曲程度不断加大(图3 D-G);凋零期,随着子房膨大到一定程度,花瓣、雄蕊(不开裂散粉)等逐渐干枯脱落,同时柱头干枯,最终形成果实(图3H)。雌花从开放到坐果(花期结束)平均耗时11.6天。图4所示为混合类型花序中雌雄花的开放顺序,雌花盛开时,雄花花药尚未开裂散粉,雌花较雄花先成熟开放,同时可知二者的成熟期有所重叠。
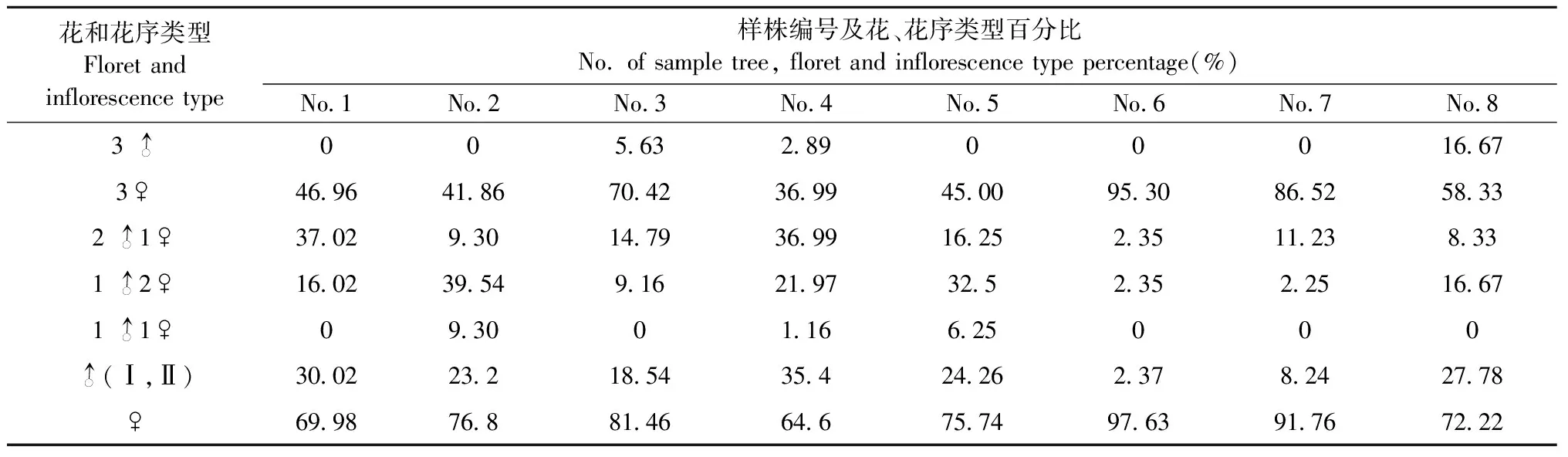
表2 血皮槭不同类型花序和雌雄花比例(2021年)①Tab.2 The percentage of female florets and male florets of A. griseum in 2021

图2 血皮槭3花序小花开放动态Fig. 2 Opening dynamics of female florets in 3 inflorescenceA: 花蕾; B: 花蕾萌动; C: 露白期; D, E, F: 开放, 散粉; G, H: 干枯, 凋零.A: Flower bud; B: Flower budding; C: Anther appearing; D, E, F: Full blooming, pollen dissemination; G, H: Floret withering.

图3 血皮槭3♀花序小花开放动态Fig. 3 Opening dynamics of female florets in 3♀ inflorescenceA: 花蕾; B: 花蕾萌动; C: 露白期; D, E, F, G: 开放; H: 柱头和花瓣枯萎并脱落, 形成果实.A: Flower bud; B: Flower budding; C: Stigma appearing; D, E, F, G: Full blooming; H: Stigmas and petals withered and fallen, fruit appearing.

图4 血皮槭混合类型花序小花开放动态Fig. 4 Opening dynamics of female floret and male florets in 12♀ inflorescenceA: 露白期, 雌花柱头伸出; B: 雌花开始开放; C, D: 雄花开放和散粉; E, F: 雄花脱落凋零, 果实形成.A: Stigma appearing exerting; B: Female florets begin to open; C, D: Male florets begin to open and disseminate pollens; E, F: Male florets withered and fallen, fruit appearing.
3.2.2 开花物候 血皮槭每年开花1次,4月初芽萌动、现花蕾,花先叶开放,单株树花开始开放时间平均比始展叶期早4.6天,一般进入盛花期时叶才开始伸展。血皮槭花开放较为集中,于4月上旬进入盛花期,4月中下旬进入末花期,花期长约19.5天(表3)。观察发现,血皮槭存在雌雄异熟现象,具体表现为雌花先熟或雄花先熟。2019年仅1株开花单株表现为雄花先于雌花开放,其余7株皆为雌花先开放。2021年则有2株树为雄花先于雌花开放,其余6株皆为雌花先开放,且同一单株在不同年份表现出相反的顺序。此外,雌花成熟期和雄花成熟期皆有重合。
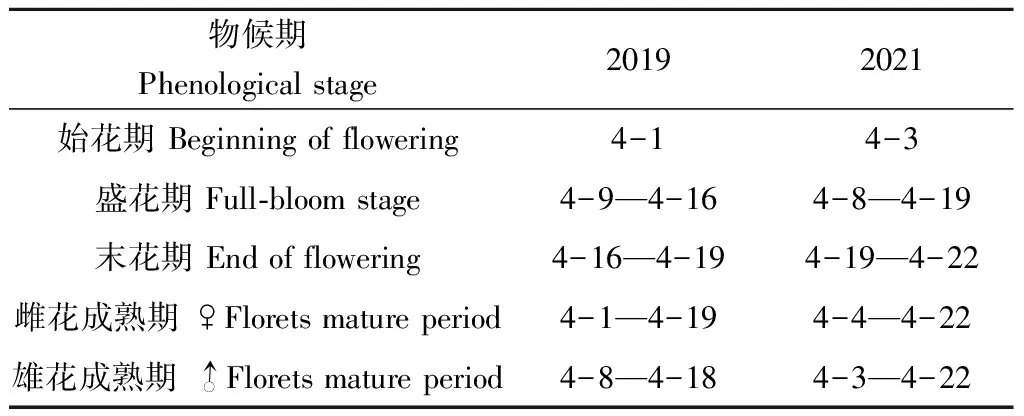
表3 血皮槭开花物候观测 (月-日)Tab.3 Observation of flowering phenology(month-date)
3.3 花粉生活力与柱头可授性
血皮槭花粉生活力的持续时间较短,如表2所示,收集到足量花粉时(0 h)花粉生活力为53.76%±2.46%,经3 h室温干燥保存后,花粉生活力最高为63.86%±5.35%,与0 h相比差异显著,12 h后花粉活力迅速降至21.12%±5.50%,24 h后降至1.03%±0.13%,近乎完全丧失生活力。
雌花不同发育时期的柱头可授性表现为: 1) 露白期,花柱未开裂,柱头顶端弱度反卷,产生气泡极少,可授性微弱或无(图5A); 2) 开放期,花柱开裂,柱头顶端轻度反卷,产生气泡的面积扩大,速度快,数量多,可授性较强(图5B); 柱头可授性与柱头姿态相关,随着柱头反卷程度加深,气泡产生速度加快,反应十分剧烈,气泡发生面积进一步扩大(图5C),可授性达到最强; 3) 凋零期,柱头顶端开始褐化干枯,柱头可授性微弱(图5D)。综上所述,柱头从露白期伸出花冠时已具有微弱可授性,随着柱头反卷程度不断加深,可授性逐渐加强,持续到花末期柱头褐化干瘪为止,可授性几乎贯穿于整个雌花花期。
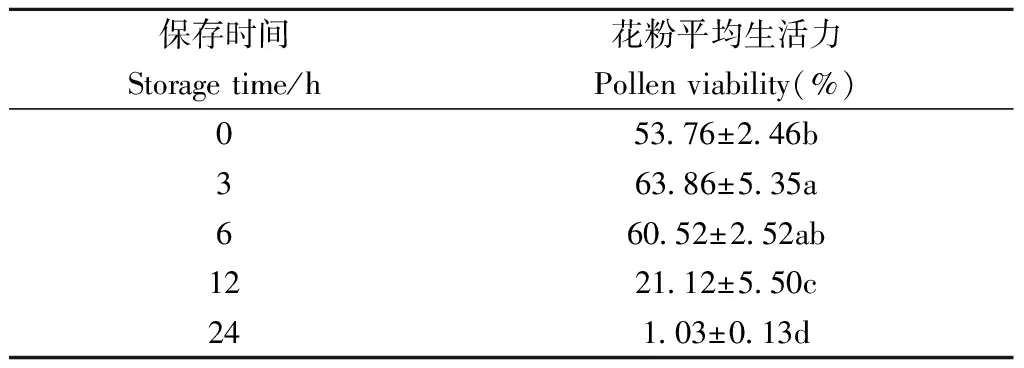
表4 室温自然条件下血皮槭花粉生活力变化①Tab.4 Pollen viability of A. griseum at room temperature
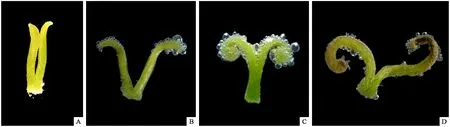
图5 雌花不同时期柱头可授性变化情况Fig. 5 Stigma receptivity on different period of female flowersA: 露白期, 花柱未开裂; B, C: 开放期, 花柱开裂, 柱头顶端开始反卷; D: 末花期, 柱头褐化干枯.A: Stigmas appearing, stigmas not dehiscent; B, C: Flowering, stigmas are dehiscent and curved; D: End of flowering, stigmas are brown and dried-up.
3.4 花粉胚珠比与异交指数
血皮槭花粉量较大,平均单花花粉量为45 900~58 900个; 单花雌蕊2心皮,2室,每室2个胚珠,平均胚珠数为4,因为雌雄异花,按照单株雌雄花比例,计算出P/O为3 093~3 969。依据Cruden(1977)标准,P/O为2 108.0~195 525.0时交配系统为专性异交,判定血皮槭交配系统为专性异交。
根据Dafni(1992)标准,血皮槭平均单花直径: 雄花为(6.50±1.14)mm,雌花为(7.72±1.08)mm,皆大于6 mm,记为3; 雌蕊柱头可授期间有雄花散粉,记为0; 柱头与花药的空间位置分离,记为1,故其OCI为4,由此判断血皮槭交配系统为异交,部分自交亲和,需要传粉者。
3.5 人工控制授粉试验结果
血皮槭人工控制授粉试验结果(表5)表明: 1)套网袋处理结实率极低,说明血皮槭风媒传粉可能性极低,以虫媒为主;2)自株异花授粉结实率(28.57%)低于自然授粉(38.95%);说明血皮睚交产生可育种子数少,存在自交亲和现象; 3)异株异花授粉结实率(67.19%)高于自然授粉(38.95%),说明自然条件下传粉效率低下,人工辅助授粉可提高结实率; 4)套袋不授粉结实率为100%,且结实率为0,说明血皮槭不经过受精过程产生空果,即存在强烈单性结实现象。
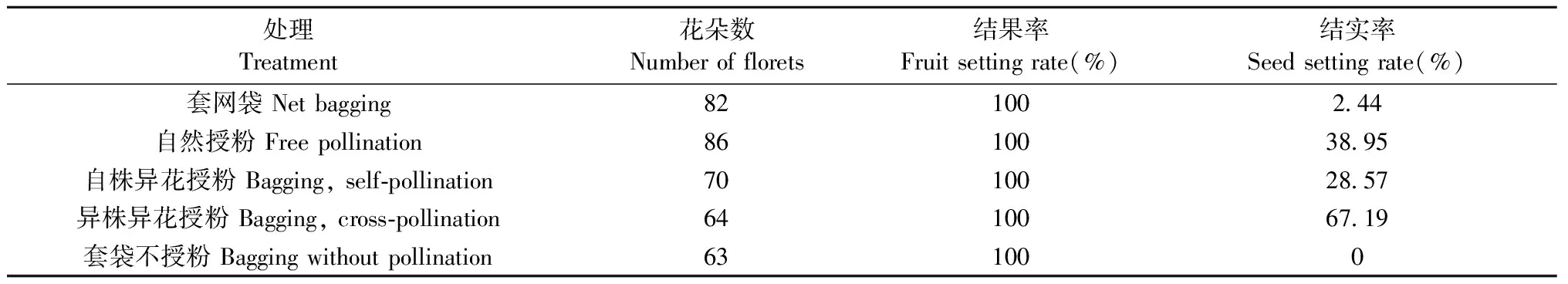
表5 控制授粉试验结实情况Tab.5 Seed setting from controlled pollinations of A. griseum
4 讨论
4.1 血皮槭交配系统类型及单性结实现象
花粉胚珠比结果表明血皮槭交配系统为专性异交,而异交指数结果则表明其交配系统为异交,部分自交亲和,需要传粉者,与人工控制授粉自交结实率28.57%相一致,由此推测血皮槭交配系统为异交,部分自交亲和,需要传粉者。血皮槭由于异交而维持群体内较高遗传多样性,与分子标记显示群体内存在部分自交的结论(孙圣,2014)吻合的。人工异株花授粉(67.19%)明显高于自然授粉结实率(38.95%),表明自然条件下传粉者数量不足或传粉效率低下可能是导致其结实率低的重要原因。同时观察发现,血皮槭植株附近(100 m内)同处花期(4月中旬)的植物有牡丹(Paeoniasuffruticosa)和海棠(Malusspectabilis)等,推测植物之间对传粉者的竞争可能影响结实率,或由于血皮槭在北京属于引进树种,其可能缺乏适合的传粉媒介。另外,本研究推测野外血皮槭单株结实率低是因传粉媒介的种间竞争导致,至于血皮槭的传粉媒介是什么,还需要对血皮槭的传粉生物学作进一步探究。
de Jong(1976)发现槭属植物普遍存在单性结实现象,但表现程度不同。单性结实不明显的树种如粗柄槭(A.tonkinense)和红花槭(A.rubrum)等,若雌花未受精,则子房会停止发育,进而脱落或果实小而干瘪; 挪威槭(A.platanoides)和红脉槭(A.rufinerve)等,若雌花未受精也能结实,但果实在发育过程中会部分脱落,单性结实较明显; 单性结实强烈的树种如梣叶槭(A.negundo)等,雌花无论受精与否均不会脱落,形成的空籽果实与正常含籽果实形态无差别,难以区分。本研究套袋不授粉试验结果率为100%,证实血皮槭存在强烈的单性结实现象,推测其为自然条件下栽培植株结实率低的原因之一。
4.2 血皮槭避免自交机制
槭属植物大部分物种表现为二重雌雄异型异熟,即植株雌雄功能按雄—雌—雄顺序分别表达,具有两期雄花(de Jong, 1976)。色木槭即表现出二重雌雄异型异熟,同时也具有雄先熟或雌先熟的现象(张灵芝等,2011)。本研究花物候观测结果表明,血皮槭开花进程中通常表现为雌先熟或雄先熟。血皮槭雌花雌蕊柱头可授期时间较长,贯穿开花始末,在雌花柱头可授期内,同一花序或同一单株雄花开放散粉,花粉生活力虽然时间短,但与柱头可授期之间存在一定重叠。在植物传粉过程中,有效的传粉是以大量的花粉、有效的媒介和具有可授性的柱头为前提(黄双全等,2000),观测中发现血皮槭在开花期通常存在大量发育时期不同的花,即血皮槭在花序水平上(混合花序)和单株水平上具有自交的可能性。综上表明,血皮槭为不完全雌雄异熟。
4.3 血皮槭单翅果单籽现象
本研究发现,血皮槭每个雌花每个心皮内含有2个胚珠,但通常只有1个胚珠发育正常,1对翅果由2个单翅果(每个单翅果由1个心皮发育而来)组成,即单翅果单籽现象。调查中偶尔观察到洛阳牡丹园内种植血皮槭仅有1个单株的有些翅果外形较大,解剖发现单翅果双籽现象,即心皮内的2个胚珠皆能正常发育,这说明多数血皮槭果实种胚发育过程中普遍存在败育现象,降低了结实率。峨眉山天然单株天全槭(A.sutchuenensesubsp.tienchuanense)地面多年残留的空果实也发现1个翅果内2个空洞存在,即单翅果双籽现象。其他槭树也偶尔发现单翅果双籽现象(van Gelderenetal., 1994)。血皮槭目前普遍为单翅果单籽,结实率很低,如果能提高单翅果双籽发生频率,则可获得更多的种子。因此,血皮槭单翅果单籽现象产生的原因及提高单翅果双籽频率的栽培措施还需要进一步研究。
5 结论
血皮槭花功能单性,雌雄异花同株,不完全雌雄异熟; 交配系统兼性异交,部分自交亲和,需要传粉者,具有强烈的单性结实倾向。单性结实和传粉效率低可能是引起血皮槭结栽培植株结实率低的重要原因。本研究对象为人工栽培植株,对血皮槭交配系统进行初步探讨,对于天然群体还需进一步通过分子标记方法结合传粉媒介的观察研究血皮槭天然群体的交配系统类型,从而揭示天然群体血皮槭结实率低的原因,同时花功能单性的形成机制以及种子发育过程中是否存在败育现象也需要进一步探究,以为制定合理的保护策略提供科学依据。
猜你喜欢
安徽农业科学(2022年18期)2022-10-13
麦类作物学报(2022年8期)2022-08-08
金沙江文艺(2022年4期)2022-04-26
江苏农业科学(2020年21期)2020-03-08
湖北农业科学(2019年20期)2019-12-11
中国瓜菜(2019年8期)2019-09-19
儿童故事画报·自然探秘(2019年10期)2019-01-14
现代农业研究(2017年10期)2018-01-02
Coco薇(2015年12期)2015-12-10
农村百事通(2009年8期)2009-05-25
