
‘马家柚’果实发育期有机酸含量变化及柠檬酸代谢相关基因的表达分析
2022-10-08 12:04:34易明亮张王妮匡柳青刘德春
江西农业大学学报 2022年4期
易明亮,张王妮,2*,杨 莉,匡柳青,刘德春,刘 勇,胡 威*
(1.江西农业大学 农学院/江西省井冈蜜柚繁育与栽培技术工程中心,江西 南昌 330045;2.陕西枫丹百丽生物科技有限公司,陕西 宝鸡 721199)
【研究意义】‘马家柚’[Citrus maxima(Burm.)Merr.,Rutaceae]是江西名优柑橘品种,在上饶市广丰地区广泛种植,其果肉呈红色、果味清香、风味独特,享有江西省酸柚类第一名的美誉[1]。有机酸含量的高低是柚类果实独特品质形成的重要指标之一[2-3],对果实口感具有决定性作用。因此研究‘马家柚’果实发育过程中各有机酸含量的变化规律及柠檬酸合成、降解、转运相关基因的表达变化对探究其果实品质形成机制具有重要价值。【前人研究进展】果实中不同的有机酸比例形成其独特的风味品质,不同柑橘品种的柠檬酸代谢机制也存在差异。高阳等[4]研究发现‘靖安椪柑’果实有机酸各组分中柠檬酸含量最高,奎宁酸、酒石酸次之,而苹果酸最低。柑橘果实发育过程中有机酸变化与柠檬酸代谢密切相关,柠檬酸的代谢大致分为合成、转运、降解3个环节。在细胞质内,柠檬酸的合成始于磷酸烯醇式丙酮酸经羧化酶(phosphoenolpyruvate carboxylase,PEPC)羧化,而后经柠檬酸合成酶(citrate synthase,CS)的催化合成柠檬酸[5]。林琼[6]的研究结果表明CitPEPCs和CitCSs基因的上调表达导致了露地栽培的椪柑柠檬酸含量显著高于温室栽培果实。柠檬酸在线粒体中合成后,转运到植物细胞液泡中储存,在液泡膜上,柠檬酸的转运受到质子泵的调控[7],其中液泡膜上的质子泵H+-ATP酶(V-ATPase)和H+-焦磷酸化酶(V-PPase)通过水解ATP 释放能量并不断的泵入H+以维持液泡中的酸性环境[8-9]。Yao 等[10]在苹果上分别鉴定出VATPase 和V-PPase 的编码基因MdVHP1和MdVHA-A,同时证明这两个基因都与苹果中有机酸的积累有关;Li 等[11]在柑橘中也发现了液泡膜质子泵编码基因CitVHA-c4与柑橘果实柠檬酸积累密切相关。另外,CHX(cation/h+exchanger)作为阳离子和氢离子的交换蛋白,可以通过增加液泡膜内的电化学梯度来调控柑橘果实中的柠檬酸积累[12],同时有研究表明CitCHX和二羧酸转运载体基因(dicarboxylate carrier gene,CitDIC)参与柑橘类水果中热空气激活的柠檬酸盐的降解[13]。柠檬酸降解始于从液泡中释放后在细胞质内经顺乌头酸酶(Aconitase,Aco)转化为异柠檬酸,而后经NADP-异柠檬酸脱氢酶(NADP-isocitrate dehydrogenase,NADP-IDH)生成谷氨酸,谷氨酸在谷氨酸脱羧酶(glutamate decarboxylase,GAD)或谷氨酰胺合成酶(glutamine synthetase,GS)作用下经谷氨酰胺或γ-氨基丁酸(γ-aminobutyric acid,GABA)途径进行降解。陈明[14]的研究结果显示CitAco3、CitGAD4/5、CitGS2的转录丰度在低酸早熟椪柑果实中均显著高于高酸的普通椪柑。曹淑燕等[15]研究砧木对黄果柑有机酸的影响,表明通过嫁接可显著提高黄果柑NADP-IDH 活性,降低果实中有机酸含量。此外,柠檬酸还可能在ATP-柠檬酸裂合酶(ATP citrate lyase,ACL)的作用下,经乙酰辅酶A 降解途径生成草酰乙酸和乙酰辅酶A,然后参与类黄酮等次生代谢物合成[16]。罗丽娟等[17]研究表明低温处理可显著降低温州蜜柑ACOs和ACLβ1基因的表达水平,同时提高汁胞中柠檬酸含量。【本研究切入点】目前,‘马家柚’前期研究多集中在气象因子、土壤环境以及栽培管理措施等外在因素对果实品质的影响[18-20],而关于‘马家柚’果实发育阶段有机酸各组分含量变化规律及其与柠檬酸代谢相关基因表达量变化的内在联系和机制研究鲜有报道。【拟解决的关键问题】为了解‘马家柚’果实发育过程中柠檬酸代谢的分子机制,本研究对‘马家柚’果实发育阶段的有机酸各组分及柠檬酸代谢相关基因的表达进行分析,以期鉴定其柠檬酸代谢的关键基因,为良种‘马家柚’的栽培管理及品质调控提供理论指导依据。
1 材料和方法
1.1 试验材料
‘马家柚’果实于2018年6月开始采自江西省上饶市广丰区芦林镇果园,选定6棵长势相同且树龄在10年以上的成年结果树,2棵为一次生物学重复,在盛花后60(S1),75(S2),90(S3),105(S4),120(S5),135(S6),150(S7),165(S8),180(S9),195(S10),210 d(S11)直至果实成熟,每个时期分别采集6个果实,选取成熟度和大小均一、无病虫害无机械损伤的正常果实,剥取赤道处果肉用液氮速冻,于-80 ℃低温中保存。
1.2 测定指标及方法
1.2.1 有机酸的提取与含量测定 果实有机酸的提取参考陈明[14]的方法,采用高效液相色谱法(HPLC)测定有机酸各组分含量。有机酸测定的色谱条件为:Water C18 柱(4.6 mm×250 mm),柱温为25 ℃,流动相为0.05 mol/L的磷酸氢二氨水溶液(pH=2.7),流速为0.5 mL/min;二极管阵列检测器检测。总有机酸含量是柠檬酸、苹果酸、酒石酸、奎尼酸含量之和。
1.2.2 RNA 的提取与逆转录 果肉RNA 的提取采用华越洋快速通用植物RNA 提取试剂盒,采用琼脂糖凝胶电泳法对提取的RNA 进行检测,RNA 逆转录采用TaKaRa公司(型号为Cat.#RR047A)反转录试剂盒合成第一链cDNA。
1.2.3 荧光定量 使用Bio-RAD 荧光定量PCR 仪,日本TaKaRa 公司的SYBR Premix EX TaqTM 荧光定量酶进行q-PCR 基因的定量表达分析,其中内参基因(Actin)与柠檬酸相关基因引物设计参考Lin 等[13]、Guo等[21]的设计(表1),引物序列委托生工生物工程股份有限公司合成,PCR反应体系采用Jiang等[22]的设计,每个样品设置3个生物学重复,基因相对表达量分析采用2-△△CT方法。

表1 荧光定量PCR引物序列Tab.1 Primer sequences of real-time fluorescence quantitative PCR
1.3 数据统计与分析
数据统计及整理采用Office365 Excel 2019软件,数据分析利用IBM SPSS 软件进行Duncan 式方差分析以及LSD检验差异显著性(P<0.05),最后采用Origin2018软件进行图形的制作。
2 结果与分析
2.1 ‘马家柚’果实发育过程中有机酸含量的变化
从图1可观察到‘马家柚’果实从幼果到成熟过程中的变化,S1~S9时期(花后60~180 d)为果实膨大期,果实横纵径及单果质量逐渐变大,之后果实不再膨大进入成熟期,同时S7~S10时期果皮由绿转黄的同时果肉也逐渐变红。在发育过程中‘马家柚’果实中柠檬酸含量在膨大期呈上升趋势,在S1~S5 时期上升较为缓慢,之后在S6~S9 时期即膨大后期快速上升。成熟初期柠檬酸含量继续上升,至S10 时期时达到最大值5.85 mg/g,之后开始显著下降(图2)。从图2中可以看到柠檬酸含量变化趋势与总有机酸基本相同,在果实膨大期不断上升而在果实成熟期后开始下降。从‘马家柚’果实整个发育期来看,柠檬酸的积累可以分为缓慢增长期(S1~S5)、快速增长期(S5~S10)和下降期(S10~S11)。而苹果酸、酒石酸和奎宁酸含量在发育期间变化量不大。‘马家柚’果实中柠檬酸含量始终最高,占总有机酸含量的45.51%~81.47%,苹果酸含量占总有机酸含量的3.07%~24.56%,酒石酸含量占总酸含量的5.65%~27.88%,奎尼酸占总有机酸含量的5.62%~23.6%,由此表明,‘马家柚’果实发育至成熟期间主要的有机酸是柠檬酸,总有机酸变化趋势主要由柠檬酸含量决定。

图1 ‘马家柚’发育过程中果实变化Fig.1 Fruit and cross section during development of‘Majia’pomelo

图2 ‘马家柚’果实发育期有机酸含量Fig.2 Organic acid content of‘Majia’pomelo during fruit development
2.2 ‘马家柚’果实发育期柠檬酸合成相关基因的表达
对柠檬酸积累的3 个阶段具有代表性的S3、S5、S7、S9、S10、S11 时期进行柠檬酸代谢相关基因的表达分析。磷酸烯醇式丙酮酸羧化酶(PEPC)和柠檬酸合成酶(CS)参与了柑橘柠檬酸的合成。由图3a 可知在‘马家柚’整个果实发育过程中,CmPEPC1在S3时期表达量相对较高,在S5~S11时期保持较低水平稳定表达且无显著变化。由图3b 可知,CmPEPC2基因相对表达量在S3~S11 整个时期相对稳定且无明显变化,两个基因的表达趋势与柠檬酸积累趋势并不一致。

图3 ‘马家柚’果实发育期柠檬酸合成相关基因的表达量Fig.3 Expression of genes related to citric acid synthesis during fruit development of‘Majia’pomelo
由图3c 可知,CmCS1基因相对表达量在S3~S7 时期持续下降,在S9~S10 时期又重新上升,到S11 时CmCS1基因相对表达量则显著下降至0.51。由图3d 可知,CmCS2基因的相对表达量在S3~S7 期间无明显变化,之后一直呈上升趋势,在成熟期S10~S11时期相对表达量急剧上升,达到最高值2.43(图3)。由此表明CmCSs基因的表达量变化对柠檬酸含量没有影响。
2.3 ‘马家柚’果实发育期柠檬酸转运相关基因表达
质子泵可以为柠檬酸转运到液泡贮藏而提供动力,对果实中有机酸积累具有正向调节作用。质子泵基因CmVHP2的相对表达量在S3~S10 时期整体呈下调表达趋势,在S10 时期达到最低相对表达量0.43,S11 时期显著上升至最大值1.35,其变化趋势与柠檬酸变化趋势并不一致(图4a)。质子泵基因CmVHA-c4在果实发育期S5~S9 呈上升趋势,在S9 时期达到最高值后缓慢下降,S10 时期虽有下降但并不显著(图4b),因而其变化趋势与柠檬酸的积累模式类似,可能对柠檬酸的积累起重要作用。
H+反向转运蛋白基因CmCHX相对表达量呈先降低后上升再缓慢降低再上升的波动趋势,与果实柠檬酸含量的变化规律没有联系(图4c)。二羧酸载体基因CmDIC在S3时期相对表达量最高,至S5时期时显著下降,并维持相对低水平表达,而此时从S5 时期开始果实柠檬酸积累刚好进入快速增长期,至S10时期开始后进入下降期,而S11 时期CmDIC基因表达量相对于S10 时期显著上调(图4d)。由此表明柠檬酸含量受到CmDIC基因的负调控,其对‘马家柚’果实柠檬酸积累具有重要作用。

图4 ‘马家柚’果实发育期有机酸转运相关基因表达Fig.4 Expression of organic acid transport related genes during fruit development of‘Majia’pomelo
2.4 ‘马家柚’果实发育期柠檬酸降解相关基因表达
在‘马家柚’果实发育阶段CmACLα1基因的表达是先下降后上升,在S7时期相对表达量最小,在S11时期时达到最大值(图5a)。CmACLβ1基因在S3 时期表达较高,随后下降,在果实发育S5~S11 时期表达水平较低,且保持相对稳定(图5b),S5时期开始柠檬酸进入快速增长期,因此CmACLβ1基因可能是马家柚果实膨大期调控柠檬酸积累的重要因素。CmAco3基因表达量在S3~S7 时期保持较低水平,之后开始上调表达,至S11时期时表达量显著升高至最高值(图5c),而此时柠檬酸含量下降,因此认为CmAco3可能是调控果实成熟期降解‘马家柚’柠檬酸的重要基因之一。
从图5f可知谷氨酸合酶基因CmGS2的表达量在柠檬酸含量较低的S3~S7时期保持较高水平表达,而在柠檬酸含量较高的S9~S11时期维持较低水平表达,因此柠檬酸含量极可能是由CmGS2基因的表达进行负调节。在果实生长发育过程中,CmNAD-IDH2基因相对表达量在S3~S5时期有下降趋势,之后在柠檬酸上升期却呈上升趋势,并且从S9 时期开始其表达量并无显著差异(图5d);CmNAD-IDH3基因的表达在S3~S7时期相对稳定,在S9时期达到最大值,随后表达量又逐渐下降,其表达量变化对柠檬酸含量没有影响(图5b)。
谷氨酸脱氢酶基因CmGAD5的相对表达量在S3~S9 时期无显著变化,在S10 时表达量最低,随后在S11时期达到最大值(图5g);γ-氨基丁酸转氨酶基因CmGABA-T的相对表达量在S3~S10时期相对稳定,在S11 时显著上调达峰值(图5h),因此认为CmGAD5和CmGABA-T基因在果实成熟期的显著上调表达也是柠檬酸降解的重要因素。
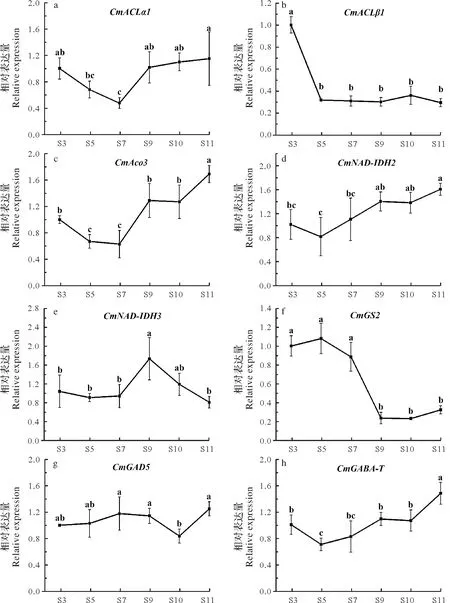
图5 ‘马家柚’果实发育期有机酸降解相关基因表达Fig.5 Expression of organic acid degradation-related genes during fruit development of‘Majia’pomelo
3 结论与讨论
柑橘果实在发育过程中有机酸逐渐积累,其中柠檬酸为主要有机酸,呈先增加后降低的趋势,而其他有机酸组分含量因品种不同而存在差异[23]。研究发现,‘马家柚’果实中有机酸以柠檬酸为主,苹果酸、酒石酸次之,奎尼酸含量最低,柠檬酸的变化趋势为先上升后下降。温州蜜柑、早香柚[24]以及‘靖安椪柑’[4]在果实发育期,其有机酸含量均呈先上升后下降趋势,柠檬酸含量均在总有机酸含量中占比最大,温州蜜柑占比56%以上,早香柚占比53%以上,而靖安椪柑更是达到71%以上。
前人对柑橘柠檬酸积累与柠檬酸合成相关基因表达之间的研究很多,大部分研究认为柠檬酸的积累与其合成相关基因无直接联系。如易明亮等[25]研究表明,温州蜜柑完熟栽培过程中柠檬酸合成相关基因CitPEPCs和CitCSs的表达与果实有机酸积累并没有联系。罗安才等[26]和Chen等[27]研究结果也表明柠檬酸合成相关酶活性以及相关基因的表达量也均与柠檬酸含量无显著相关性。在本研究中CmPEPC1、CmPEPC2、CmCS1和CmCS2基因的相对表达量变化与柠檬酸含量变化规律不一致,他们之间无显著相关性,说明‘马家柚’果实发育过程中柠檬酸的积累并不是由柠檬酸合成基因决定。
柠檬酸合成后需要通过载体运输到液泡中进行保存,其降解同样需要通过载体从液泡中运输至细胞质中进行降解反应。质子泵基因促进柠檬酸运输至液泡贮存,对提高果实柠檬酸含量具有积极作用。Guo 等[21]的研究表明V 型质子泵基因CsVHA、CsVHP和p 型质子泵基因CsPH8在低柠檬酸突变品种‘红暗柳’橙中的表达水平明显低于‘暗柳’橙;李绍佳[28]的研究表明CitVHA-c4基因的表达模式与椪柑果实发育过程中柠檬酸的积累呈正相关关系,同时在烟草叶片中瞬时过表达CitVHA-c4基因可显著提高叶片中柠檬酸含量。并且在不同柑橘品种中各运输载体基因在柠檬酸积累的过程中发挥的作用不尽相同[24]。有机酸转运相关基因CHX和DIC可将柠檬酸从液泡中运输至细胞质中促进其降解,Lin 等[13]的研究证实低酸的温州蜜柑中CitCHX和CitDIC基因表达水平显著高于高酸品种‘高橙’,同时在烟草叶片中瞬时超表达CitCHX和CitDIC基因可显著降低烟草叶片中柠檬酸含量。在‘马家柚’果实发育期CmCHX和CmVHP2基因表达量变化与柠檬酸含量变化趋势并不一致;而柠檬酸含量受CmDIC基因负调控,与此同时CmVHA-c4基因的表达模式与柠檬酸的变化趋势相近,认为CitDIC和CmVHA-c4基因在‘马家柚’果实柠檬酸的积累过程中发挥着重要作用。
柑橘果实发育过程中柠檬酸的积累与柠檬酸降解基因的表达相关,柠檬酸可以通过多个途径进行降解,主要有谷氨酰胺途径、γ-氨基丁酸(GABA)和乙酰辅酶A 3 个途径。Terol 等[29]研究了甜橙、橘、柠檬等多个柑橘种类果实发育过程中柠檬酸的变化规律及CcACOs基因的表达水平变化,结果表明在除柠檬外的大部分品种中CcACOs基因的表达能促进果实中柠檬酸的降解。在椪柑叶片及果实中瞬时超表达CitAco3基因或调控CitAco3基因的转录因子基因CitNAC62和CitWRKY1时均能显著降低其柠檬酸含量[30],同时Chen 等[31]研究发现热处理能促进柑橘果实CitAco3基因的上调表达导致柠檬酸下降。以上研究均说明ACO基因在促进柠檬酸降解的途径中发挥着重要作用。另外,有研究报道在脐橙果实生长发育过程中,CitAco3-CitIDH1-CitGS2的级联反应可能参与了不同环境下柠檬酸的降解,而CitGS2基因是谷氨酰胺途径中的关键酶基因[27]。Katz 等[16]发现果实柠檬酸含量下降过程中GAD 酶活性持续升高,同时Lin 等[32-33]证实在椪柑的果实成熟过程中,以及在低温诱导下柠檬酸的降解主要与CitGAD4基因的表达密切相关,Sheng 等[34]的研究表明HB 柚采后储存过程中CitGABA-T基因高表达导致了柠檬酸的降解,而CitGAD4和CitGABA-T基因均是GABA 途径上关键的酶基因。在柑橘不同品种果实发育过程对转录本的分析表明乙酰辅酶A 途径的关键基因ACLs对果实柠檬酸含量的调控作用根据品种的不同而有所差异,同时仅有CitACLβ1基因可以受干旱和ABA 诱导下调表达造成柠檬酸含量的显著上升[35],在温州蜜柑和纽荷尔脐橙果实中也发现编码ACL 的基因表达升高导致柠檬酸含量的降低,说明CitACL基因在柠檬酸降解过程中发挥着重要作用[36]。本研究的结果显示CmACLβ1基因在‘马家柚’果实发育中后期低水平表达是柠檬酸积累的重要原因;CmAco3基因在柠檬酸下降时显著上调表达,因此认为CmAco3基因在果实成熟期柠檬酸降解过程中发挥着关键作用。而柠檬酸前期缓慢积累后期大量积累也受降解基因CmGS2前期的高表达以及后期的低水平表达所控制,说明谷氨酰胺途径是‘马家柚’果实发育前期柠檬酸降解的重要途径之一。CmGAD5和CmGABA-T基因的表达量在果实发育前中期表达相对稳定,而在后期显著上调表达,说明CmGAD5和CmGABA-T基因是‘马家柚’果实在成熟期柠檬酸降解的关键基因,且在‘马家柚’果实发育后期(果实成熟期)柠檬酸降解主要以GABA 途径为主;而CmACLβ1基因可能是马家柚果实膨大期调控柠檬酸积累的重要因素,其下调表达直接导致柠檬酸的快速积累,由此说明乙酰辅酶A 途径可能是果实发育前期柠檬酸降解的重要途径。因此‘马家柚’果实发育期间柠檬酸降解在不同时间段主要通过不同的代谢途径进行,而不同柑橘品种在果实发育期柠檬酸降解途径是有差异的,如早熟温州蜜柑在果实发育期其柠檬酸降解是通过谷氨酰胺和GABA 途径进行[14],普通温州蜜柑在果实发育期柠檬酸降解则主要通过谷氨酰胺途径进行[6],而‘纽荷尔’脐橙则主要通过GABA 途径降解柠檬酸,CsAcos、CsIDH1/3与CsGAD4表达水平决定果实柠檬酸含量[37]。
本研究明确了‘马家柚’果实有机酸在整个发育期的变化规律,其有机酸以柠檬酸为主,柠檬酸积累呈先上升后下降的趋势。在整个果实发育期柠檬酸的积累并不由柠檬酸合成相关基因决定,而是由柠檬酸转运载体基因以及降解基因决定。在果实发育前中期,柠檬酸的积累主要受CmDIC、CmVHA-c4、CmGS2和CmACLβ1基因的调控,其代谢主要是通过谷氨酰胺途径和乙酰辅酶A 途径进行;而在果实发育后期柠檬酸的积累在受CmVHA-c4基因调控的同时也受CmAco3、CmGAD5和CmGABA-T的调控,其降解主要通过GABA 途径进行;因此本研究丰富了柑橘类果实发育期柠檬酸积累及代谢规律。
致谢:江西省柑橘产业技术体系项目(JXARS-07)同时对本研究给予了资助,谨致谢意!
猜你喜欢
江苏调味副食品(2021年1期)2021-04-01 12:51:12
河北果树(2020年4期)2020-01-09 16:06:08
家庭医学(下半月)(2019年9期)2019-10-12 08:03:56
中成药(2018年5期)2018-06-06 03:12:15
上海农业学报(2016年5期)2016-02-10 06:52:51
应用化工(2015年2期)2015-07-13 03:12:26
中国茶叶加工(2015年3期)2015-02-27 07:55:30
应用化工(2014年1期)2014-08-16 13:34:08
长江大学学报(自科版)(2014年4期)2014-03-20 13:20:35
河南城建学院学报(2014年4期)2014-02-27 07:08:52
