
宁夏地区番茄种质资源核心种质构建策略
2022-10-07 09:16郑福顺王晓敏李国花李洪磊刘珮君胡新华付金军
浙江农业学报 2022年9期
郑福顺,王晓敏,2,3,4,*,李国花,李洪磊,刘珮君,胡新华,付金军
(1.宁夏大学 农学院,宁夏 银川 750021; 2.宁夏现代设施园艺工程技术研究中心,宁夏 银川 750021; 3.宁夏优势特色作物现代分子育种重点实验室,宁夏 银川 750021; 4.宁夏设施园艺(宁夏大学)技术创新中心,宁夏 银川 750021; 5.宁夏巨丰种苗有限责任公司,宁夏 银川 750021)
番茄()原产于南美洲,是我国人民主要食用蔬菜之一。番茄是严格的自花授粉作物,长期的人工选择会导致其遗传背景逐渐变窄,广泛地收集番茄种质资源是有效解决这一问题的手段之一。随着种质资源的材料份数日益增多,种质资源的保存与利用也逐渐困难。
核心种质的概念是澳大利亚学者在1984年首次提出的,即以最小的种质资源数量最大限度地保留遗传多样性。核心种质的构建将有效解决种质资源数量多且保存困难等问题。近年来,园艺作物核心种质的构建也被相关领域的研究者所关注。核心种质构建策略的展开一般从遗传距离、抽样比例、抽样方法和系统聚类法4个因素考虑。牛玉等对219份樱桃番茄种质7个性状的基因型值,按照30%的抽样比例,采用2种遗传距离、8种聚类方法和3种抽样方法进行樱桃番茄核心种质构建策略研究,并评价不同方法构建樱桃番茄核心种质的优劣。贺滉等对185份大果番茄种质资源7个性状的基因型值,分别采用2种遗传距离法、8种不同的聚类方法、3种不同的抽样方法,按照30%的抽样比例对大果番茄核心种质的构建策略展开研究,均旨在为核心种质构建提供方法和理论依据。但是牛玉等和贺滉等都是对遗传距离、系统聚类法和抽样方法依次比较筛选,缺少各条件之间的比较,无法准确地比较出不同条件之间组合后构建核心种质的优劣。
本研究拟通过对前期调查的480份番茄种质资源的20个表型性状数据,依次对最短距离法、最长距离法、中间距离法、重心法、类平均法、可变类平均法、可变法、离差平方法共8个系统聚类法和10%、15%、20%、25%、30%共5个抽样比例分别进行对比,并对不同方法的优劣进行排序;在此基础上对2个遗传距离、排名前二的抽样比例、3个抽样方法和排名前二的系统聚类法进行组合试验,并对所构建的24个核心种质进行代表性评价,最终确定最佳的构建策略。以期为番茄核心种质构建提供方法,为番茄种质资源新品种选育提供便利。
1 材料与方法
1.1 试验材料
试验材料为宁夏大学农学院蔬菜课题组和宁夏巨丰种苗有限责任公司共同收集的480份番茄种质资源,材料类型与来源详见文献[10]。
1.2 试验方法
1.2.1 性状调查
依照《番茄种质资源描述规范和数据标准》,具体调查的性状为首花序节位、单花序果数、商品果纵径、商品果横径、果梗洼大小、果梗洼木栓化大小、单果重、硬度、可溶性固形物含量、果肉厚、心室数、畸形果率、花序类型、成熟果色、叶色、绿肩、生长势、叶片着生状态、裂果性和坐果性共20个表型性状。
1.2.2 系统聚类法对比
利用QGA Station 2.0软件,利用欧氏距离、10%的抽样比例、随机取样法和8个系统聚类法(最短距离法、最长距离法、中间距离法、重心法、类平均法、可变类平均法、可变法和离差平方和法)进行不同方法的对比,并根据核心种质代表性评价对聚类方法进行排序。
1.2.3 抽样比例对比
利用QGA Station 2.0软件,采用欧氏距离、随机取样法、最短距离法和5个不同的抽样比例(10%、15%、20%、25%和30%)进行不同方法对比,根据核心种质代表性评价对抽样比例进行排序。
1.2.4 核心种质最佳构建方法的筛选
利用QGA Station 2.0软件,对2个遗传距离、排名前二的抽样比例、3个抽样方法和排名前二的系统聚类法进行组合试验,如表1所示,进行构建核心种质并筛选出构建核心种质的最佳方法。

表1 核心种质不同构建方法信息表
1.2.5 核心种质代表性评价
Hu等提出的4个评价参数分别为均值差异百分率、方差差异百分率、极差符合率和变异系数变化率。核心种质的代表性评价按照均值差异百分率<20%、极差符合率>80%,变异系数变化率和极差符合率越大就越能够代表原种质群体的遗传多样性。利用均值检验和方差检验,检验核心种质与原群体的差异,均值差异越小,方差和变异系数变化率越大,则说明核心种质的代表性越好。
2 结果与分析
2.1 系统聚类法的对比
8个系统聚类法构建的8个核心种质的差异百分率信息如表2所示。各核心种质的极差符合率均>80%,除CX4的均值差异百分率为40.00%>20%,不符合核心种质构建原则外,其他核心种质的均值差异百分率均<20%。将符合核心种质构建原则的7个核心种质的均值、方差和变异系数与原群体进行差异比较,结果如表3所示。CX8、CX7、CX2和CX6的均值与原群体基本保持一致,分别有5、4、4和3个性状的方差与原群体存在显著或极显著性差异;CX3的果梗洼木栓化大小的均值与原群体存在显著性差异,6个性状的方差与原群体呈显著或极显著性差异;CX5和CX1虽然符合核心种质构建原则,但是在均值检验中,分别有7个和2个性状的均值与原群体存在差异,核心种质的代表性较差;7个核心种质的20个性状的变异系数大部分比原群体有所提高;对所构建的核心种质代表性评价结果表明,各核心种质构建方法的排名顺序为离差平方和法>可变法>最长距离法>可变类平均法>中间距离法>最短距离法>类平均法。

表2 不同系统聚类法构建的核心种质与原始群体差异百分率

表3 不同系统聚类法构建的核心种质与原始群体差异比较

续表3 Continued Table 3
2.2 抽样比例对比
5个抽样比例所构建的5个核心种质的差异百分率信息如表4所示。各核心种质的极差符合率均>80%,均值差异百分率<20%。将符合核心种质构建原则的5个核心种质的均值、方差和变异系数与原群体进行差异比较,结果如表5所示。CC2的首花序节位、生长势和畸形果率3个性状的方差与原群体存在显著性差异,畸形果率的均值与原群体呈显著差异;CC5有8个性状的方差与原群体存在显著性差异;CC5、CC4、CC3和CC1均有2个性状的均值与原群体存在显著性差异;5个核心种质的20个性状的变异系数大部分比原群体有所提高。所构建的核心种质代表性评价排名顺序为15%>30%>25%>20%>10%。

表4 不同抽样比例构建的核心种质与原始群体差异百分率

表5 不同抽样比例构建的核心种质与原始群体差异比较
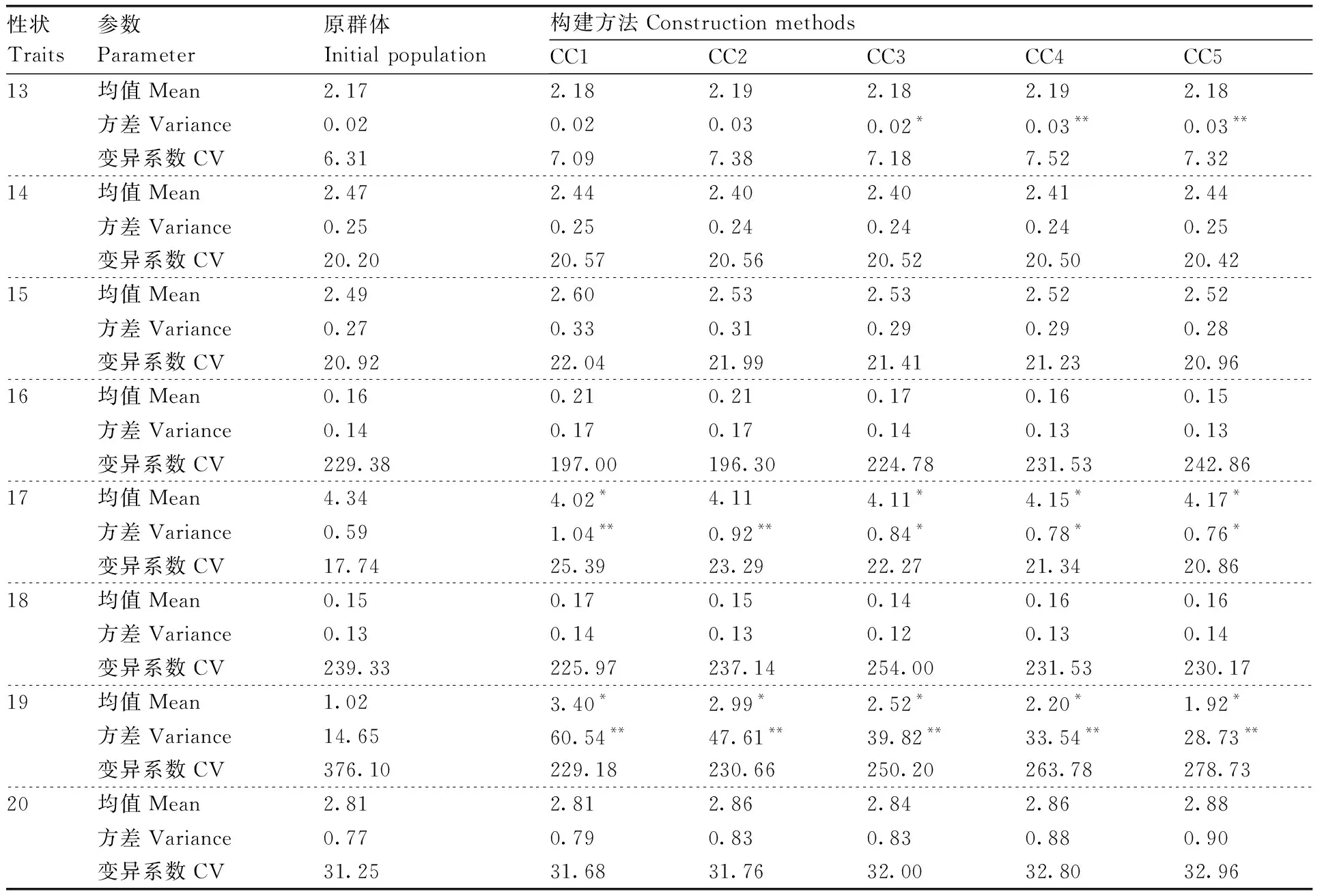
续表5 Continued Table 5
2.3 核心种质最佳构建方法的筛选
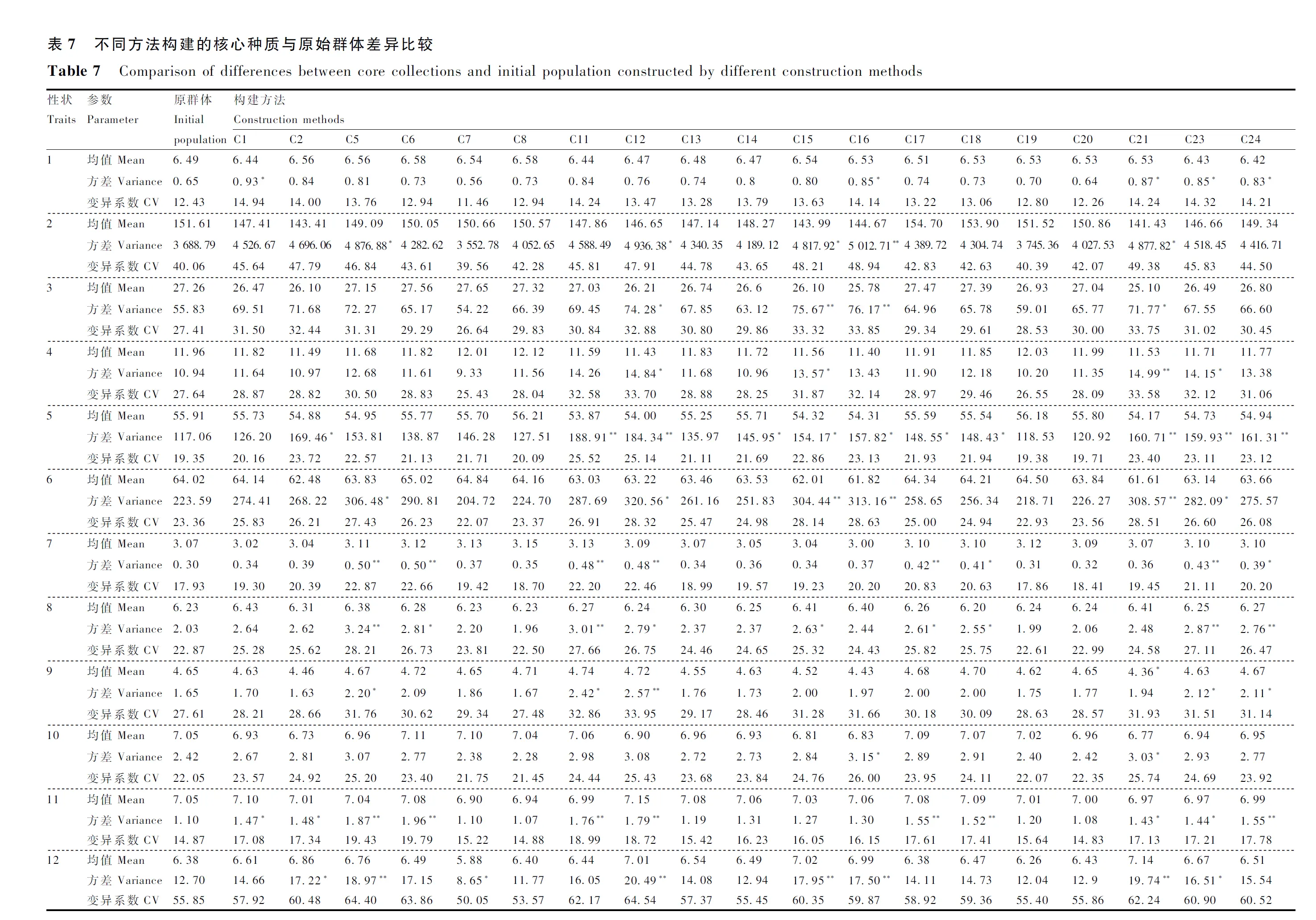
对2个遗传距离、2个最佳的抽样比例、3个抽样方法和2个最佳的系统聚类法进行组合试验构建核心种质,结果如表6所示,各核心种质的极差符合率均>80%,除C3、C4、C9、C10和C22的均值差异百分率分别为60.00%、50.00%、45.00%、60.00%、40.00%,均>20%,不符合核心种质构建原则,其他核心种质的均值差异百分率均<20%。将符合核心种质构建原则的20个核心种质的均值、方差和变异系数与原群体进行差异比较,结果如表7所示。所有核心种质20个性状的均值与原群体基本保持一致,C12共15个性状的方差与原群体存在显著性差异,其中,9个性状的方差与原群体呈极显著性差异;C23有14个性状的方差与原群体存在显著性差异;C24、C21和C5分别有12、14、10个性状的方差与原群体呈显著或极显著差异;C5、C11、C12、C15、C16、C17、C21、C23和C24的方差差异百分率较高,表明各核心种质获得了一定的变异;极差符合率均>95.00%,表明各核心种质对特殊种质有所保留;各核心种质大部分性状的变异系数比原群体有不同程度的提高,表明核心种质具有较好的异质性。综上所述,本研究中C12为最优核心种质构建方法,即遗传距离为马氏距离、抽样比例为15%、抽样方法为偏离度取样法和系统聚类法为离差平方和法。

表6 不同构建方法构建的核心种质与原始群体差异百分率

3 讨论
核心种质资源能够以最少的种质材料代表原群体所包含的全部或绝大部分遗传变异,有效解决种质资源数量庞大和保存困难的问题。表型数据可以有效直观地表现出种质资源的表型特征,而且保存种质资源的基本出发点是对其各表型性状进行利用,因此,对核心种质的构建和评价过程中不应该缺失表型数据。邓学斌等研究表明,基于表型构建的核心种质表型变异代表性较好,基于基因型构建的核心种质群体结构代表性较好。而构建核心种质时表型变异代表性和群体结构代表性同等重要,因此,利用表型构建核心种质仍然十分重要。
前人研究表明,在构建核心种质时应该首先对构建策略进行筛选。由于种质资源的收集过程、保存年份、遗传变异程度等存在差异,因此,不同作物构建核心种质时的取样比例大小也不尽相同,但取样比例大小的选择应以原群体种质资源数量而确定。雷刚等基于表型数据对603份辣椒进行核心种质构建研究,当抽样比例为15%时,各项指标均最佳。郝晓鹏等从663份普通菜豆中筛选出152份核心种质材料,最终确定20%的抽样比例最佳。张欢等利用161个水青树种质的叶表型性状对核心种质构建方法进行了探索,筛选出72份核心种质材料,即45%的抽样比例最佳。这些学者研究结果均表明,当构建核心种质的原群体样本量越大时,抽样比例应越小,反之,抽样比例应越大。本研究供试材料480份,最终筛选抽样比例为15%最佳,符合这一规律。徐盛春等研究表明,在取样比例较小时,随着取样比例的升高,核心种质的极差符合率急剧升高。当达到一定取样比例以后,极差符合率和变异系数变化率的变化开始减小。本研究取样比例为10%时,极差符合率为93.46%,当取样比例为15%时,极差符合率急剧升高至96.08%,当取样比例分别为20%、25%、30%,即大于等于20%时,极差符合率分别为96.75%、97.86%、97.91%,三者之间变化不大,符合上述规律。
郑福顺等根据牛玉等、Hu等、邓学斌等核心种质构建策略中筛选的最佳方法,确定欧氏距离、10%的抽样比例、3种抽样方法和类平均法成功构建出2个核心种质。然而在对核心种质的构建策略进行研究时发现,刘子记等、牛玉等和贺滉等都是固定抽样比例后,先对遗传距离进行比较,筛选出最佳的遗传距离后,对系统聚类法进行比较,筛选出最佳的系统聚类法后,再对抽样方法比较,筛选出最佳的抽样方法,单因素筛选最佳方法,缺少了不同因素组合条件下的对比,无法准确地比较出不同条件之间组合后构建核心种质的优劣。为探求构建种质资源核心种质的最佳策略,本研究旨在对构建策略中的4个因素进行组合试验。常见的8个系统聚类法和5个常用的抽样比例(10%、15%、20%、25%和30%),若直接设置组合后数据量太大。因此,本研究依次对8个系统聚类法和5个抽样比例分别进行对比与排序,在此基础上再对2个遗传距离、排名前二的抽样比例、3个抽样方法和排名前二的系统聚类法进行完全组合试验,共构建24个核心种质,与原种质群体对比后,明确C12为最优核心种质,即遗传距离为马氏距离,抽样比例为15%,抽样方法为偏离度取样法,系统聚类法为离差平方和法。郑福顺等以与本研究相同的试验材料,不同构建策略构建的2个核心种质虽成功构建了宁夏地区番茄种质资源核心种质,但R1仅有2个性状的方差与原群体存在显著或极显著差异,D1有8个性状的方差与原群体存在显著或极显著差异,R1、D1的变异系数绝大多数高于原群体;而本研究中C12共有15个性状的方差与原群体存在显著性差异,其中9个性状的方差与原群体呈极显著性差异,表明本研究筛选的最佳方法构建的核心种质更具有原群体的代表性。本研究的组合试验增大了可比性,同时也增大了覆盖度,组合结果更有说服力。最终确定的最佳方法不仅能最大限度的保留原群体的遗传信息,而且还具有良好的异质性。但是单一的表型数据尚不能完全说明问题,因此,本研究下一步将结合SNP分子标记技术,基于基因型数据进行核心种质构建的策略研究,以期更加准确地保证核心种质的代表性,为后续工作奠定基础,为宁夏地区番茄种质资源相关研究提供理论依据。
猜你喜欢
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
安徽农业科学(2022年9期)2022-05-17
中国蔬菜(2022年2期)2022-03-10
汽车实用技术(2022年4期)2022-03-07
华东师范大学学报(自然科学版)(2019年5期)2019-11-11
初中生世界·九年级(2017年10期)2017-11-08
电子技术与软件工程(2016年23期)2017-03-06
中学生数理化·八年级数学人教版(2016年5期)2016-08-23
中学生数理化·八年级数学人教版(2016年5期)2016-08-23
