
酿酒葡萄抗寒相关基因的研究进展
2022-10-04 05:04:02刘牧青孙雨航代玲敏
酿酒科技 2022年9期
马 岩,刘牧青,孙雨航,钟 轲,韩 宁,代玲敏
(齐鲁工业大学(山东省科学院)生物工程学院,山东济南 250353)
葡萄酒近年来在中国越来越被广大人民接受,但葡萄酒的原料——欧洲葡萄品质虽好,但抗寒性较差。我国国土面积广大,因气候原因,北方地区冬季温度过低导致葡萄植株受损而质量下滑。随着气候与土壤条件的不断变化以及一些植物的跨地域栽培,植物对环境的适应变得极其重要。在一些极端条件下,植物为了适应环境不断进化出一套防御体系。Jiang 等通过实验发现,对低温有一定抵抗能力的葡萄光合作用及与糖生物合成相关酶的活性比较强,这是植物开始适应低温环境的一种本能反应;范高韬从欧亚种玫瑰香中分离得到一种基因并命名为,经低温诱导及荧光定量PCR检测后发现,该基因表达量在低温诱导前期没有显著性变化,24 h 后显著上调,这表明该基因可能被其他冷相关基因诱导表达。由此看出,基因的表达并不是独立,而是一个甚至多个调控网络。
随着分子生物学等的不断发展,利用分子生物学等方法探索在低温条件下抗寒基因的表达途径。第二信使分子在细胞内的信号传递过程中扮演着重要的角色,钙离子与脱落酸(Abscisic Acid,ABA)比较常见。信号传递一般分为两类,不依赖ABA途径与依赖ABA途径。
不依赖ABA 信号传导途径:在低温条件下,植物细胞膜发生相变,这种刺激信号使Ca内流,细胞内的Ca受体蛋白激酶被磷酸化,CRLK 作为连接CRPK 与MAPK 级联途径的桥梁,低温信号通过该途径被传递并逐渐放大。Xu 等从山葡萄中克隆出两种ICE 基因(、),当细胞感受到低温信号后,在Ca信号作用下,基因编码MYC 型转录因子并激活基因的表达,基因编码bHLH 型转录因子以激活基因的表达,随后转录因子与抗寒相关基因启动子上的CRT/DRE 顺式作用元件结合,启动下游基因(、,基因相比于基因特异性调控基因)的表达。依赖ABA 信号传导途径:当细胞感受到低温信号后,ABA 浓度升高,在ABA 受体PYL 上合成AREB 蛋白,该蛋白与CE 元件偶联结合到启动子片段的ABRE 顺式作用元件上启动下游基因的表达(见图1)。
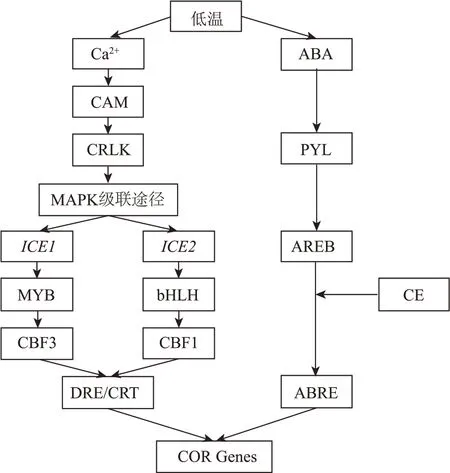
图1 抗寒基因表达通路
低温是限制植物生长发育及存活的逆境胁迫,不但会改变酶的生存条件,使酶活性降低甚至是丧失,还会改变细胞膜的状态。当膜系统遭受冷冻伤害,为了适应低温条件,细胞膜的不饱和脂肪酸比例升高,由于抗氧化酶活性受到影响,不饱和脂肪酸容易被氧化成脂氢过氧化物。脂氢过氧化物不仅会分解成丙二醛,还会均裂产生脂性自由基,这种物质既会不断损伤细胞膜,又会氧化蛋白质从而使蛋白质发生聚集。低温还会降低与光合作用相关的酶类活性:1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco)、磷酸核酮糖激酶(PRK)、3-磷酸甘油醛脱氢酶(GAPDH)等,减少糖类的合成。王依等[选取4种酿酒葡萄以筛选低温适应性强的葡萄品种,低温处理后发现过氧化氢酶活性以及可溶性脂肪含量升高,脯氨酸、可溶性糖及超氧化物歧化酶活性先升高后下降,过氧化物酶活性降低,电导率及丙二醛含量升高。2018年,贺兰山地区发生低温灾害,导致90%一年生葡萄幼株、30%~40%两年生葡萄幼株受冻死亡。低温条件会触发植物自身的抵抗机制来抵御胁迫,但仍会对其造成极大的损害。因此我们需要采取并对比一切可以提高葡萄抗寒性的合理措施以使葡萄安然越冬。
1 提高葡萄抗寒性的措施
提高葡萄越冬能力的方法多种多样:(1)埋土。埋土作为广泛采用的能使葡萄安然越冬的方法,从结果来看是极其有效的。葡萄在越冬期间,一年生枝干以及地表附近的根系容易受到损伤,土埋增加了根系周围的土壤厚度,可有效提高根系周围温度,保护根系免受损伤,在来年可使其有效吸收水分及营养。但其存在的问题也比较明显:首先,土埋比较费时费力,取土也存在一定风险,只从葡萄株周围取而不回填可能会人为对根系造成损伤,取土会造成取土区土壤厚度下降,在冬季对其造成二次伤害;其次,在葡萄成熟期以及采摘期,如果操作不得当,还会使腐烂的葡萄颗粒留在土壤中使其感染病害。(2)选择抗寒的砧木嫁接。不同砧木的抗寒性、抗旱性、抗盐碱能力及抗病性存在差异,需要根据葡萄品种的特性选择合适砧木。对5种抗寒性较好的砧木(5BB、5C、SO4、140R、kangzhen3)综合抗寒力进行比较,发现5BB 抗寒性最高。(3)灌水和熏烟。该方法需要及时关注天气情况,在霜冻来临前5 h 灌水,或点燃碎木屑,利用燃烧产生的热量增加株间温度。(4)建设保温棚。胡悦等发现保温棚可以有效提高棚内地下20 cm 以内的温度,空气湿度、土壤湿度都能有效提高,但是大棚的建设成本比较高,费时费力,适合小规模使用。(5)建设防护林。在冬季风向的垂直方向建造防护林,使冷空气沿树体爬升。(6)喷施壳聚糖。壳聚糖分解后产生低聚合度的碱性寡糖可引起葡萄植株的应激反应,增加植株内渗透调节物的含量,降低冰点,提高抗寒性。(7)选择合适的葡萄园,尽可能避开冷空气聚集的低洼区。(8)通过基因工程的手段,定向提高葡萄的抗低温能力。目前主要是通过转入调控基因或者目的基因来改善葡萄的抗寒性。因为基因的插入位点随机,需要筛选抗性优良的转基因植株以稳定遗传。目前发现低温转录因子(、等)、脱水素家族、单锌指结合蛋白家族和泛素/26S 蛋白酶体系的表达会显著提高植物的抗寒性。外源基因在植物的有性生殖中具有遗传稳定性,可避免每年投入大量的人力物力。从长远角度考虑,利用分子生物学及遗传学方法与技术定向提高葡萄的抗寒性具有很光明的前景。
2 抗寒相关基因
2.1 抗寒相关转录因子
CBF 转录因子属于AP2/EREBP 型转录因子,是植物低温胁迫中主要依赖的一条抗寒途径。在拟南芥中发现的CBF1、CBF2、CBF3 和CBF4 基因,前三者位于四号染色上,后者位于五号染色体上且被发现是四类CBF 基因中唯一一个与ABA 调控途径互作的基因。当植物遭受低温胁迫,ICE(inducer of CBF expression)基因被诱导表达,该基因结合到CBF 基因的启动子区域直接驱动CBF 基因的表达。CBF 蛋白随后结合到COR 基因启动子的CRT/DRE 顺式作用元件上启动下游基因的表达。从山葡萄中克隆得到VaCBF1 基因,经胁迫处理及荧光定量PCR 检测后发现,该基因受低温诱导表达,不受山梨醇和氯化钠诱导。其他转录因子,如、、、及等都可以提高植物抗寒性。
2.2 抗寒功能基因
2.2.1 脱水素基因
脱水素(Dehydrin,DHN)是一种晚期胚胎发育丰富蛋白(late embryogenesis abundant protein,LEA),属于LEA 蛋白的第二家族,分子量在9~200 KDa 之间,在种子发育后期和逆境条件下大量积累。Malik发现在非生物胁迫下,脱水素基因在转录和翻译水平均上调且植物的抗逆性提高。脱水素由3 个保守片段:K 片段、S 片段和Y 片段组成,Mota还发现富含极性氨基酸的、不保守的φ片段参与形成SKn 型大豆脱水素。K 片段位于C末端,可形成双亲性的α-螺旋结构,K 片段与阳离子抗菌多肽性质极度相似,脱水素表达会抑制大肠杆菌的生长。S 片段由一系列丝氨酸组成,易发生磷酸化,与核定位有关的信号肽结合,进入细胞核,与脱水素定位到细胞核并帮助肽链初步折叠行使分子伴侣功能有关。还有一些S 片段包含LHR 序列(GXGGRRKK),能够与K 片段连接,也可发生磷酸化。Y 片段((V/T)DEYGNP)位于N 末端,其氨基酸序列与植物和细菌的分子伴侣的核酸结合位点的氨基酸序列具有同源性。Y 片段富含甘氨酸,与组织特异性有关,亚细胞定位发现含Y 片段的脱水素在种子中大量表达,其他不含Y片段的脱水素没有组织特异性。脱水素含有的3个保守片段数量和排列方式被分为五类,不同类型脱水素响应的胁迫也存在区别(见表1)。

表1 不同类型脱水素的性质与响应模式
其中,YSK型脱水素以其双亲性的特点,通过静电作用与细胞膜的脂质分子结合,在细胞失水条件下可以防止膜聚集而保护膜结构。KS 型脱水素可以直接结合金属离子或间接清除活性氧自由基,降低氧化损害。SK和K型脱水素在低温胁迫下有明显积累。脱水素最早在研究棉花子叶中的转运蛋白时被发现,这类蛋白因在逆境胁迫下大量累积被广泛关注。脱水素的功能主要体现在以下几个方面:(1)K 片段赋予脱水素的双亲性结合膜结构,防止膜发生聚集;(2)结合细胞内的金属离子维持细胞内抗氧化酶的活性,降低由金属离子产生的活性氧自由基;(3)具有分子伴侣的功能。脱水素会在植物中选择性表达避免浪费机体资源,如亚洲白桦树为适应冻害在冬季表达大量脱水素蛋白但在夏季不会表达。第一次表明脱水素具有抗冻活性的研究是从桃中分离到的PAC60 基因,它既可以防止细胞内冰晶形成,又可以产生热滞效应,使植物在一定程度上免受冰晶造成的物理损伤,提高植物的耐低温性。过表达榴莲脱水素基因的拟南芥经低温处理,发现脱水素可以提高转基因拟南芥的抗寒性。将梅花脱水素基因转入烟草后发现转基因烟草的叶片相对含水量、可溶性糖含量都明显升高,抗寒性也随之升高。Hincha认为脱水素对一些关键酶产生的保护作用与它本质上无规则的结构密切相关,脱水素维持酶的活性有两个可能:(1)依赖其不规则的结构形成物理障碍,阻碍蛋白质的聚集变性;(2)脱水素可以防止蛋白质错误折叠从而维持酶活性。Sánchez 等从仙人掌中分离得到一个SK3 型脱水素基因,发现该基因的表达产物在植物体内可以自身相互作用,K 片段与富含组氨酸的基序在自身互作中起着关键性作用。脱水素分子量大小与其对乳酸脱氢酶的保护能力呈正相关。葡萄中的脱水素基因比较少,共分离出4种 并 命 名 为、、、,其 中受低温、高温、干旱和白粉病诱导表达,受高温、低温诱导表达,后两者仅在胚胎发育后期的种子中表达。
2.2.2 DNA结合单锌指蛋白基因
DNA 结合单锌指(DNA binding with one zinc finger,Dof)蛋白是植物中特有的一类蛋白,单锌指结构由4个半胱氨酸与一个锌离子结合组成。
该蛋白一般由200~400个氨基酸组成,N末端是52 个氨基酸组成的具有保守性的功能结合结构域,可结合DNA 及蛋白质以相互作用;C 末端含有不保守的转录调控域,赋予Dof 蛋白多样性。AAAG 基序是Dof 蛋白结合域的核心结合位点,Dof 蛋白还会与细胞内的可与DNA 结合的未磷酸化的HMG 蛋白互作,增加Dof 蛋白的结合能力。Dof 蛋白还参与植物中的种子萌发、碳氮代谢、开花调控及光调节等。Zhang 等从苹果中分离出60 个Dof 蛋白基因,大多数响应低温胁迫。Dof 蛋白是一种多功能转录因子,Su 等发现,GhDof 在低温条件下可被诱导表达,并提高细胞内、、基因的表达及种子含油量;Ma等发现白菜基因在低温条件下被上调;李成慧在葡萄当中鉴定出25 个基因,其中11 个基因参与响应生物和非生物胁迫;代伟娜发现基因响应激素诱导,对MeJA 响应最为明显,MeJA 通常与植物抵御病虫害等生物胁迫息息相关。
2.2.3 泛素/26S蛋白酶体基因
泛素是一类由76 个氨基酸组成的高度保守的小分子蛋白质。N 端的1 个甲硫氨酸及7 个赖氨酸可以与其他泛素分子结合形成泛素链以调节生物体不同的生命活动。泛素/26S 蛋白酶体系是真核生物体内降解蛋白的有效途径,可降解生物体内80%~85%的蛋白质。该体系包含泛素、泛素激活酶(E1)、泛素结合酶(E2)、泛素连接酶(E3)、蛋白酶体。泛素在有ATP 供应下被E1 激活,随即与E2 结合形成E2-泛素结合体,再与E3 连接并将泛素直接或间接转移到蛋白上,使蛋白泛素化以被蛋白酶体识别并降解。泛素连接酶可被分为四类:U-box、HECT、RING 和F-box,前三者属于单亚基类,后者属于多亚基类。U-box 与RING 相似度较高,二者较明显的区别在于U-box 类连接酶靠盐桥和氢键维持结构的稳定性,RING 类连接酶靠半胱氨酸和锌离子构成的锌指结构维持稳定性。Ubox 拥有4 类结构域:ARM、Ser/Thr kinase、WD40和MIF4G。ARM 位于U-box 类型泛素连接酶的C末端,可与其他蛋白互作。Yao 等发现中国野生华东葡萄泛素连接酶基因经低温诱导后在葡萄原生质体中大量表达,通过酵母双杂技术发现,与互作,低温条件下促进VpICE1 蛋白积累,抑制VpHOS1 蛋白对VpICE1 蛋白的降解作用。在过表达的拟南芥中,低温处理后经RT-PCR 检测发现不止该基因上调,与低温胁迫有关的、、等基因也被上调。另一种U-box 类E3 泛素连接酶从左山一号被分离出并被命名为,该基因在低温条件下在各个组织中大量表达,在拟南芥中过表达该基因后,发现与低温相关的/、/及等冷相关基因被上调,但/基因被下调,该现象说明响应低温胁迫的同时也会影响其他冷相关基因的表达。
3 总结与展望
提高欧洲葡萄抗寒性的方法多种多样,具体可划分为两类:(1)改变葡萄生存的外部环境;(2)研究抗寒相关基因,并通过基因工程的手段改变葡萄自身以适应不利生存条件。本文阐述重要抗寒调控基因和抗寒功能基因的功能及其对植物抗寒性的影响,为研究葡萄抗寒基因并定向改良葡萄抗寒性提供参考。ICEs 类转录因子在遭受低温胁迫时率先表达,作为一种“诱导剂”诱导CBFs 类转录因子的表达,最终该类型转录因子结合到抗寒相关基因启动子区域的顺式作用元件CRT/DRE 上启动下游基因表达。脱水素基因表达的蛋白主要对细胞内的膜结构起到保护作用,起到分子伴侣的功能,保护细胞内关键酶的活性,保护能力与脱水素的大小呈正相关关系;单锌指蛋白在植物遭受低温胁迫时,通过调节光合作用合成生命活动所必需的物质;泛素/26S 蛋白酶体则通过降解蛋白以维持细胞内与抗寒相关蛋白的含量,提高植物抗寒性。三个类型的基因在低温条件下都会引起其他大部分冷相关基因上调及小部分基因下调,说明基因的表达受到一系列小分子/大分子物质调控。目前国内外还存在抗寒基因及表达方式认知不足的问题,因此我们还需不断挖掘抗寒基因,探索它们的表达方式以缓解低温给植物生长造成的压力。
猜你喜欢
今日农业(2021年20期)2021-11-26 01:23:56
河北果树(2020年4期)2020-11-26 06:04:28
新疆农垦科技(2016年2期)2016-08-21 13:50:26
广西林业科学(2016年2期)2016-03-20 05:53:26
中国果菜(2016年9期)2016-03-01 01:28:40
中国马铃薯(2015年5期)2016-01-09 06:11:11
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15 12:47:42
中国医学科学院学报(2015年5期)2015-03-01 04:03:46
园艺与种苗(2015年10期)2015-02-27 11:27:51
现代检验医学杂志(2015年2期)2015-02-06 02:01:01
