
大鳞鲃形态特征及其同工酶电泳分析
2022-10-02 11:00张涛周剑光张林甘金华何力
中国渔业质量与标准 2022年4期
张涛,周剑光,张林,甘金华,何力
(农业农村部水产品质量安全风险评估实验室(武汉), 农业农村部淡水鱼类种质监督检验测试中心,中国水产科学研究院长江水产研究所, 武汉 430223)
大鳞鲃(Barbuscapito)属于鲤形目(Cypriniformes)、鲤科(Cyprinidae)、鲃亚科(Barbinae)、鲃属(Barbus),主要分布于里海南部和咸海水系、乌兹别克斯坦、伊朗和土耳其等内陆河流,是一种具有较高经济价值的鱼类,具有耐盐碱、生长速度快、肉质鲜美、食性广、适应性强等特点[1]。中国约有4.6×107hm2低洼盐碱水资源,主要分布在东北、华北和西北地区,为开发和利用中国内陆盐碱水域,改变上述地区盐碱水域养殖品种单一、缺乏优质品种的现状,2003年中国水产科学研究院黑龙江水产研究所从乌兹别克斯坦引进大鳞鲃进入国内,目前已在黑龙江、内蒙古、天津等多个省市的淡、咸水池塘养殖,养殖产量和规模也逐渐在中国北方渔业中占据一席之地[2-3]。
目前有关大鳞鲃的报道主要集中在营养学特性[4-7]、发育生物学[8-9]、生理学特性[10-13]、细胞遗传学特性[14]、繁殖生物学[3, 15]及分子遗传学特性[16]等方面,有关大鳞鲃形态特征方面的研究仅见零星报道[17-19],并且多是与体质量相关的可量性状参数分析,主要为选育种提供参考依据。有关生化遗传特性方面的报道仅见葛彦龙等[20]的研究,该研究报道了大鳞鲃肝脏、肾脏和眼睛3种组织的同工酶,但关于大鳞鲃其他组织的同工酶尚未见相关报道。本研究通过形态学观察、可数可量性状测定,并结合聚丙烯酰胺凝胶电泳技术检测不同组织中的乳酸脱氢酶(LDH)以及苹果酸脱氢酶(MDH)表达情况,同时筛选出大鳞鲃种质的特征生化遗传参数,并与已有研究结果进行比较分析,旨在从形态特征和生化遗传角度进一步丰富大鳞鲃种质资源方面的研究内容,为其种质标准的制定提供理论依据。
1 材料与方法
1.1 实验材料
实验用大鳞鲃于2018年3月采自湖北省荆州市,为人工养殖群体,共30尾,体重范围为456.3~843.0 g,均值为(651.4±103.2) g;体长范围为33.2~40.2 cm,均值为(36.6±1.8) cm。
1.2 实验方法
1.2.1 形态测定
按照《养殖鱼类种质检验第3部分:性状测定》(GB/T 18654.3—2008)标准[21]的规定,对30尾样本鱼进行形态观察,并对可数可量性状进行测定。可数性状计数参数包括背鳍分支鳍条数、胸鳍分支鳍条数、腹鳍分支鳍条数、臀鳍鳍条数、尾鳍条数、侧线鳞数、侧线上鳞数、侧线下鳞数及脊椎骨数,其中脊椎骨的计数需通过解剖实验鱼后观察并计数。可量性状测量参数包括全长、体长、体高、头长、吻长、眼径、眼间距、尾柄长和尾柄高。全长及体长参数用自制量鱼板测量,其他参数用游标卡尺进行测量,并计算可量性状的比例值。
1.2.2 组织酶液的制备、电泳及染色
冰浴条件下剪鳃放血,趁实验鱼尚存活时取心脏、眼睛晶状体、肝脏、肾脏和脾脏组织样品,各组织样品经预冷生理盐水冲洗干净后用滤纸吸干表面水分。样品称重后置于洗净预冷的匀浆器中,按1∶3(m/v)比例加入预冷双蒸水,冰浴条件下反复研磨至浆状,4 ℃条件下12 000 r/min离心30 min,重复3次,收集上清液分装,-80 ℃保存备用。
采用聚丙烯酰胺凝胶垂直板电泳,电极缓冲液为pH 8.3的Tris-甘氨酸,恒压220 V电泳,LDH所用凝胶为不连续浓度胶,浓缩胶、分离胶浓度分别为7.5%、4.0%,MDH和EST所用凝胶浓度为9.0%连续浓度胶,电泳结束后取出凝胶板,室温下避光染色,待显现清晰条带后以去离子水漂洗凝胶板2~3次,然后将凝胶板平放在自制灯箱上采用尼康数码相机拍照,同工酶的染色参照张涛等[22]的方法。
1.2.3 模式图的绘制
采用Bandscan 5.0(Glyko, 美国)电泳图谱中的酶带进行灰度识别,并根据识别灰度绘制电泳图谱模式图。
1.2.4 酶的命名与分析
同工酶的命名和分析参考熊全沫等[23]的方法,以各酶带的相对迁移率(Rf)从小到大依次命名并顺序编号。
1.3 数据处理
所得可量性状数据采用SPSS 20.0(IBM公司,美国)进行分析,结果以(平均值±标准差)表示。
2 结果与分析
2.1 大鳞鲃形态描述及可数、可量性状
观测30尾大鳞鲃(图1),体呈梭形,头部较小,体修长,口亚下位,具有吻须和颌须各一对。背鳍起点位于腹鳍起点前端,尾鳍正尾形,背鳍起点至吻端距离比至尾鳍基距离短。身体被覆发光鳞片,鱼体背部呈银灰色,腹部银白色。

图1 大鳞鲃外观形态Fig.1 Morphological measurement of Barbus capito
大鳞鲃鳔2室,前室短,后室长。下咽齿3行,齿式为2·3·5/5·3·2。脊椎骨数为46~49。左侧第一鳃弓外侧鳃耙数为20~28。腹膜呈黑色。大鳞鲃可数、可量性状见表1,表中范围给出了所测各指标的上下限,均值反映了所测数据的集中程度。可数性状中,背鳍条数稳定,侧线鳞数较多,变化范围也较大。可量性状中,主要以体长和头长为参照,给出了吻长、眼径和眼间距等头部主要参数与头长的比例关系,也反映了体高、尾柄长和尾柄高等躯干部主要参数与体长的比例关系。
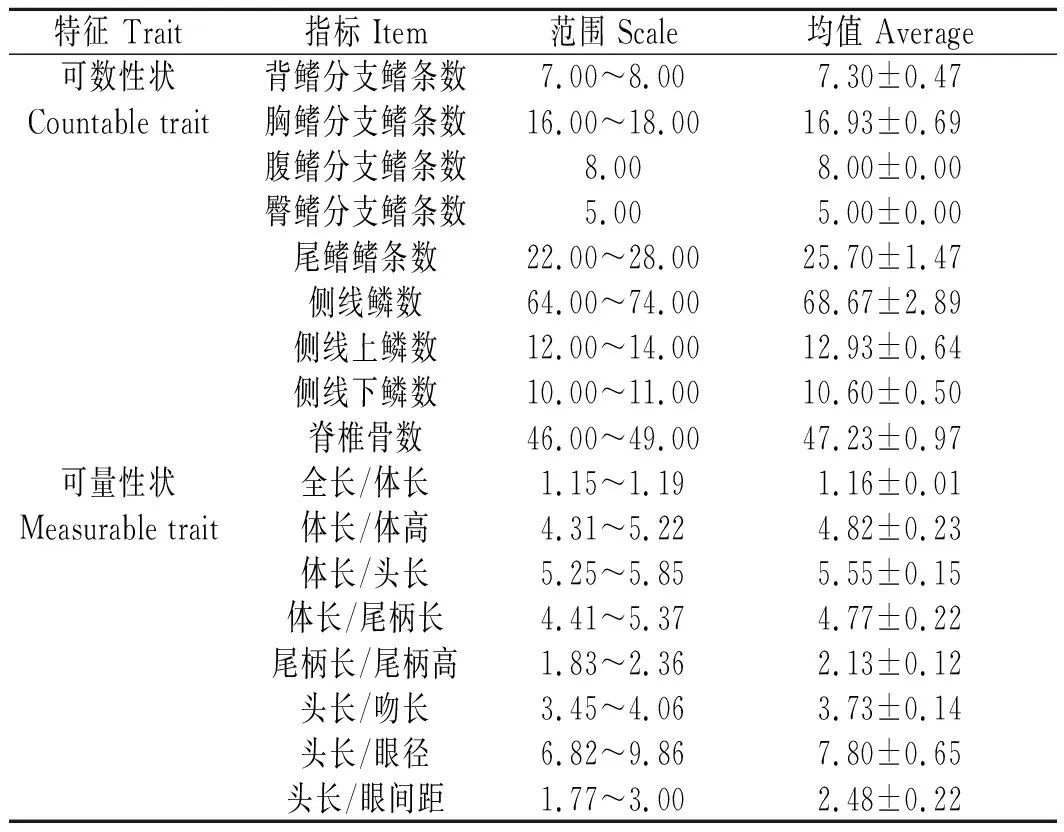
表1 大鳞鲃的可数性状和可量性状的均值与标准偏差Tab.1 The mean values and standard deviation of countable and measurable traits of Barbus capito n=30
2.2 大鳞鲃LDH的表达
LDH同工酶在大鳞鲃5种组织中的表达结果如图2所示。心脏组织中LDH同工酶的着色程度很深,可见其表达的活性很强,共检测到7条酶带(图2A),其中LDH1~LDH5表达活性较强,其余2条酶带表达活性相对较弱,并且只在1号、2号和4号鱼中有检出;LDH同工酶在眼睛晶状体组织中着色程度最浅(图2B),可见其在眼睛晶状体中表达的活性最弱,与其他4种组织相比,检出的LDH酶带数也最少,共检测出5条酶带,5号鱼表达活性太低或未表达,在酶谱图上未检出,其余4尾鱼都有5条LDH酶带检出,2号鱼和3号鱼LDH酶带的表达活性较强,4号鱼中LDH酶带的表达活性最弱;LDH同工酶在肝脏组织中表达的酶带数最多,共检测到13条LDH酶带(图2C),4号鱼表达活性太低或未表达,在酶谱图上未检出,LDH10~LDH13在电场作用下迁移的距离不远,离阴极较近,LDH9仅在2号和3号样本鱼中有检出,使得这2尾样本鱼中检测到的LDH酶带最多,LDH1和LDH2在5号样本鱼表达活性过低或未表达,没有检出,该样本鱼中检出的酶带最少,共检测到10条LDH酶带,LDH3~LDH5、LDH12和LDH13在1~3号样本鱼中表达的活性程度相对较强;LDH同工酶在肾脏组织中强烈表达,共检测到10条酶带(图2D),酶带的着色程度较深,LDH2~LDH5、LDH8和LDH9表达活性均较强,LDH1在1号样本鱼中表达活性较其他4尾样本鱼弱,4号样本鱼中各LDH酶带表达活性较其余样本鱼强;脾脏中LDH酶带表达活性程度也较强,共检测到8条酶带(图2E),5号样本鱼中LDH酶带表达活性程度相对其他4尾鱼弱,4号样本鱼LDH酶带表达活性最强,LDH4和LDH5表达活性程度较其他LDH酶带强,LDH2表达活性相对较弱。

图2 大鳞鲃LDH电泳图谱A、B、C、D、E分别表示心脏、眼睛晶状体、肝脏、肾脏和脾脏的LDH酶谱;1~5泳道分别表示不同个体的酶谱。Fig.2 Electrophoretogram of LDH isozymes in Barbus capitoA, B, C, D, E show electrophoretograms of LDH isozymes expressed in heart, eyes, liver, kidney and spleen respectively. 1-5 show the zymograms of different individuals.
2.3 大鳞鲃MDH的表达
大鳞鲃5种组织MDH同工酶的表达情况如图3所示,从MDH酶带在各组织中着色程度来看,眼睛晶状体着色最浅,肝脏着色最深,表明肝脏中MDH同工酶表达活性程度最强,而眼睛晶状体中最弱。心脏中共检测出5条MDH酶带(图3A),MDH4和MDH5表达活性程度较其余3条MDH酶带强。MDH酶带在眼睛晶状体中表达情况见图3B,1号样本鱼眼睛晶状体中MDH酶带表达活性较其余4尾样本鱼弱,MDH1和MDH2在1号样本鱼中过低表达或未表达,反映在酶谱上该酶带未检出。MDH3~MDH5酶带表达活性程度较其余2条MDH酶带强。MDH1和MDH2在各样本鱼中都较低表达。肝脏组织中5条MDH酶带表达活性程度都强(图3C),尤其2号样本鱼中MDH1和MDH5。MDH2酶带表达活性较其余4条MDH酶带弱, MDH1酶带在1号、4号和5号样本鱼中表达活性较弱。肾脏组织中也检测到5条MDH酶带(图3D),表达活性程度较强,其中MDH5在各样本鱼中都强烈表达,MDH3和MDH4表达活性次之,MDH1和MDH2表达活性相对较弱。MDH在脾脏中共有5条酶带检出(图3E),除MDH1和MDH2外,其余3条MDH酶带都较强表达,尤以MDH5明显,MDH1和MDH2在3~5号样本鱼中表达很弱。
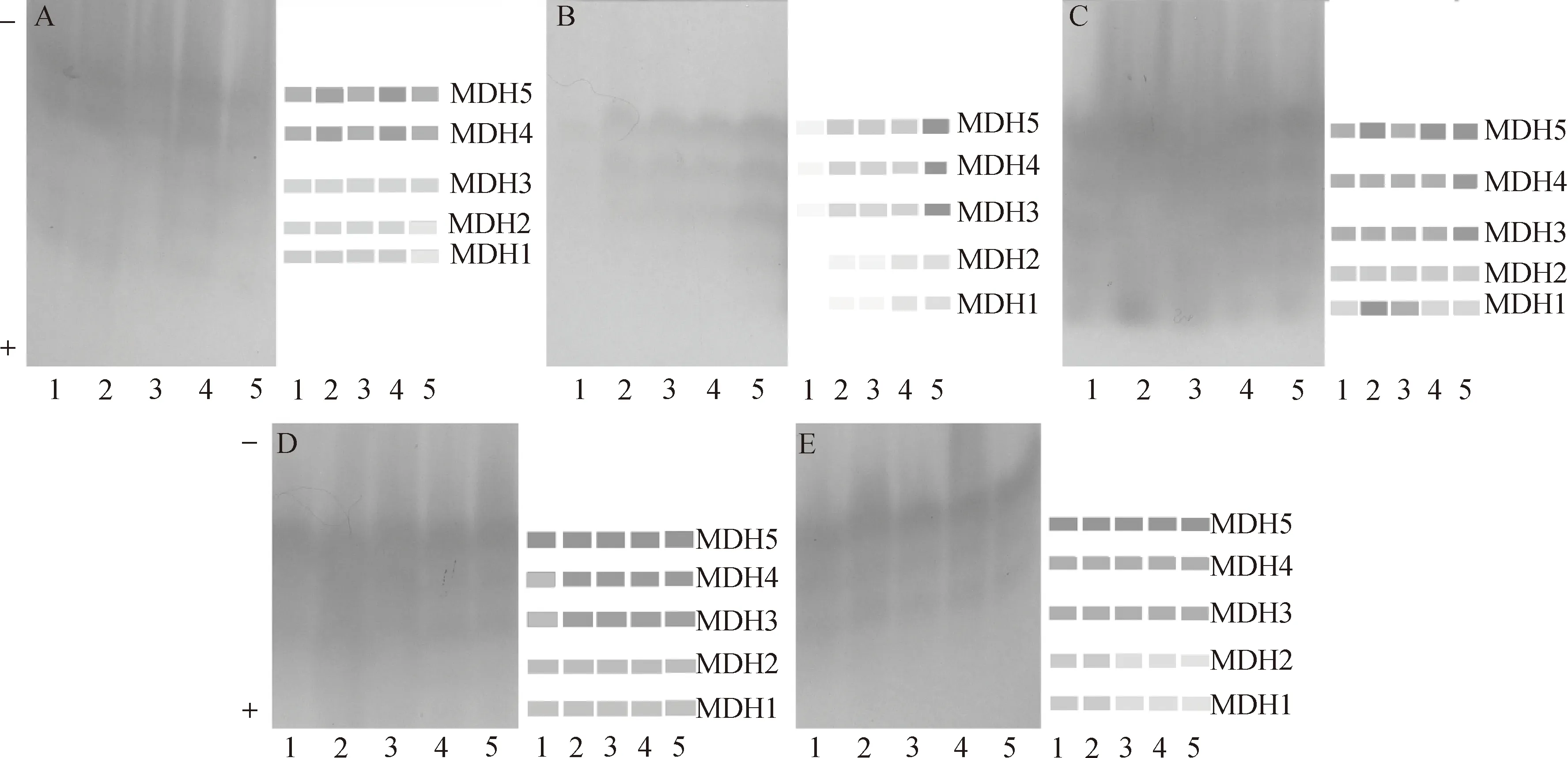
图3 大鳞鲃MDH电泳图谱A、B、C、D、E分别表示心脏、眼睛晶状体、肝脏、肾脏和脾脏MDH酶谱;1~5泳道分别表示不同个体的酶谱。Fig.3 Electrophoretogram of MDH isozymes in Barbus capitoA, B, C, D, E show electrophoretograms of MDH isozymes expressed in heart, eyes, liver, kidney and spleen respectively. 1-5 show the zymograms of different individuals.
2.3 大鳞鲃EST的表达
大鳞鲃EST同工酶的表达如图4所示,在大鳞鲃5种组织中肝脏EST酶带数最多,着色相对最深;眼睛晶状体EST酶带着色相对最浅,说明EST同工酶在大鳞鲃肝脏中强烈表达,在眼睛晶状体组织中表达活性弱。心脏组织中共检测到3条EST酶带(见图4A),EST1酶带表达活性相对比其他2条酶带强,尤其2号鱼EST1酶带强烈表达;5尾样本鱼眼睛晶状体中共检出5条EST酶带(见图4B),其中1号样本鱼EST酶带表达活性较其他4尾样本鱼弱,EST2酶带在1号鱼和2号鱼中表达活性过低或者未表达,酶谱上未检出。EST3~EST5酶带表达活性较其余2条酶带强。EST酶带在大鳞鲃肾脏中表达较强,共检测出5条酶带(见图4C),EST2在1号样本鱼中表达过低或未表达,酶谱上未检出,EST2和EST3在2号、3号和4号样本鱼中较强表达,尤以3号样本鱼明显。肝脏和肾脏中共有10条EST酶带检出(见图4D),5尾样本鱼两种组织中分别共检出10条、6条EST酶带,图4D中左边5条酶带示肝脏EST同工酶,右边5条酶带示肾脏EST同工酶。肝脏中EST9和EST10酶带着色最深,其表达活性程度较其余8条酶带强,EST1和EST3酶带仅在1~3号样本鱼中有检出,EST5酶带在4号样本鱼中未表达或过低表达,酶谱上未检出。肾脏中EST同工酶表达活性也较强。EST7、EST9和EST10酶带在肾脏中表达活性较其余几条酶带强,EST4和EST5酶带仅在1号和2号样本鱼肾脏中有检出。
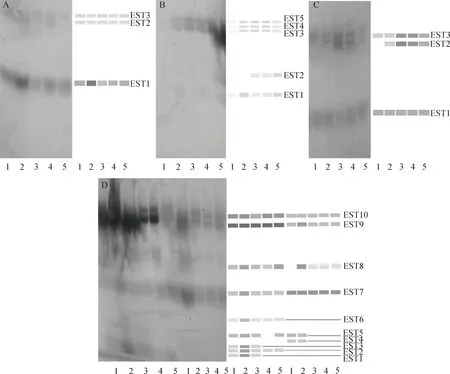
图4 大鳞鲃EST电泳图谱A、B、C分别表示心脏、眼睛晶状体和脾脏EST酶谱,D表示肝脏和肾脏EST酶谱;1~5泳道分别表示不同个体的酶谱。Fig.4 Electrophoretogram of EST isozymes in Barbus capitoA, B, C, show electrophoretograms of EST isozymes expressed in heart, eyes and spleen respectively, D show electrophoretograms of EST isozymes expressed in liver and kidney. 1-5 show the zymograms of different individuals.
3 讨论
3.1 大鳞鲃的形态学比较
关于大鳞鲃形态学特征的报道主要见蔺玉华[24]和安丽等[25]的研究,本研究中大鳞鲃的形态特征与上述已有报道基本一致。可量性状方面的差异主要体现在体长/尾柄长、头长/吻长和头长/眼径上,造成可量性状的差异可能与以下几方面的原因有关:一是样本的来源不同,养殖鱼类获取饵料较野生鱼类容易且能得到保障,其体形通常较野生鱼类短,野生鱼类相对较修长,李思发等[26]的研究表明生活在江河流水环境中的鲤(Cyprinuscarpio)体形较长,而饲养在池塘里的鲤体形较短;二是测量的标准不同,不同的人对同一可量性状起点和终点的界定不同,如对尾柄长有几种认识:臀鳍基部后端至尾鳍基部最后一枚椎骨末端[21]、臀鳍基部后端至尾鳍基部和臀鳍基部后端至最后一个鳞片;三是类似眼径这种测量值很小,较小的测量误差就能造成较大差异;四是不同测量者的主观因素差异造成测量结果不同。在可数性状方面,本研究结果与已有研究报道的比较见表2,其中表2中蔺玉华等[24]研究仅列出与其相近规格的大鳞鲃。本研究结果与已有研究报道大部分一致,少数可量性状如尾鳍条数、侧线鳞、侧线上鳞和侧线下鳞存在差异,分析认为造成这一差异的原因除了与样本量大小有关外,还可能与测定者的主观判断有关,如尾鳍鳍条往往排列紧密,尤其是尾鳍上叶或者下叶边缘处的鳍条更加细密,在计数时须借助显微镜或者放大镜,避免造成漏数或者多数,同样计数侧线上鳞或者侧线下鳞时,通常以背鳍基部或者腹鳍基部处的鳞片为计数起点,而这些鳞片通常外观形态未完整或者类似半圆形状的鳞片,不同的测定者取舍可能存在差异,有的测定者认为形态不完整可不做计数,而有的测定者认为应该计数,这些鳞片相对较小,尤其规格小的鱼类更不容易看清楚,容易漏数或者多数,同样需要借助显微镜或者放大镜。
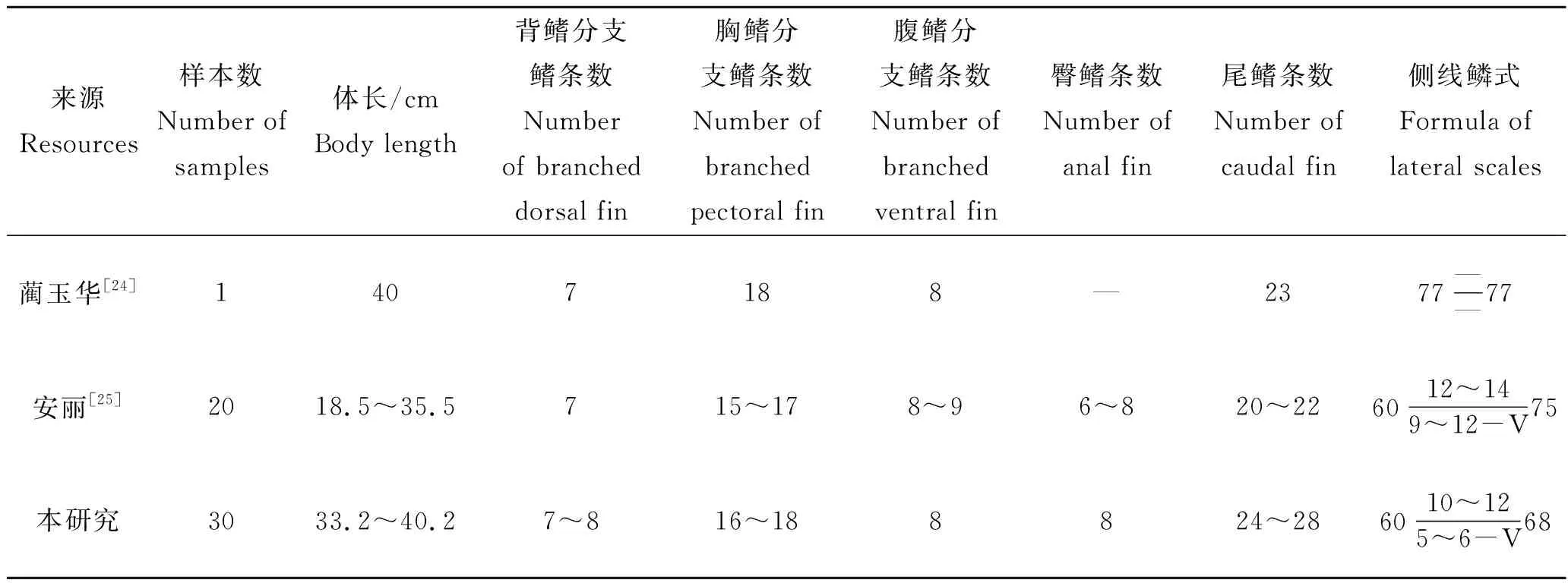
表2 大鳞鲃可数性状的比较Tab.2 Comparations of countable traits of Barbus capito in different research results
3.2 大鳞鲃同工酶表达的组织特异性
本研究中,大鳞鲃的各种组织中均能检测到LDH、MDH和EST同工酶的表达,由此可见大鳞鲃体内LDH和MDH分布比较广泛。和大多数鱼类一样,3种同工酶在大鳞鲃5种组织中的表达均具有组织特异性,这种组织特异性体现在两个方面:一是同一种同工酶在大鳞鲃不同组织中检出的酶带数目不同,以LDH同工酶为例,大鳞鲃眼睛晶状体中LDH酶带数相对其他4种组织都少,而肝脏中检出的LDH酶带数最多。另一方面,即使检出的酶带数相同,但表达的活性程度也存在差异,如MDH同工酶在肝脏中表达活性程度较其他4种组织强,但检出的MDH同工酶酶带数相同。分析认为造成这种组织特异性表达的原因主要受到两方面的影响:一是受遗传基因调控的影响,不同类型的同工酶在胚胎发育和细胞分化过程中起到的调控作用不同,如LDH同工酶在胚胎发育和细胞分化中具有显著的分化调控模式,表现出高度的发育和组织特异性[27],也就是说同工酶的组织特异性是胚胎发育和细胞分化的需要,也就决定先天发育过程中不同组织同工酶就具有组织特异性;另一方面,同工酶的表达除了受遗传基因调控外,还受到代谢调控的影响,不同组织或器官代谢过程不同,也就决定了某些同工酶仅在特定的器官或组织中表达[28]。
LDH同工酶是最早发现也是研究最深入的同工酶之一,通常认为LDH是由2个位点编码的四聚体酶,LDH-a和LDH-b基因编码了A和B两个亚基,再由这两个亚基随机组合成A4、A3B、A2B2、AB3、B45种同工酶[27]。LDH-c基因一般只在特定组织中表达,具有明显的组织特异性,对鲤形目鱼类而言一般认为LDH-c基因编码的酶只在肝脏中表达[29],可称为,一般“肝带”迁移距离距阴极较近[30]。大鳞鲃肝脏组织中LDH10~LDH13可能由LDH-c基因编码,其余组织都具有经典的5条LDH谱带,多出的LDH谱带可能是复等位基因表达的结果,这在中华倒刺鲃(Spinibarbussinensis)的研究中也观察到类似的现象[27]。
同工酶表达的组织特异性,可能是因为不同组织行使不同的生理功能,造成组织或器官间同类同工酶在含量和活性表达上的不同[31],如LDH、MDH和EST在大鳞鲃肝脏中表达活性程度都强于其他4种组织,酶谱上体现在酶带数较多或者酶带着色程度较深,这与肝脏是重要的合成、代谢和解毒器官相适应,这种现象也出现在其他鲃亚科鱼类中,如EST同工酶在黑脊倒刺鲃(Spinibarbuscaldwelli)肝脏中表达较其他组织都丰富,至少可鉴别出12条酶带[32]。葛彦龙等[20]的研究表明大鳞鲃肝脏EST、LDH、MDH和AMY淀粉酶的活性比其余2种组织高;张娟等[27]的研究表明中华倒刺鲃6种组织中肝脏的LDH强烈表达,LDH同工酶条带可达9条,肝脏MDH活性表达最强;冯为慧等[33]在研究刺鲃同工酶时也发现肝胰脏EST同工酶活性高,与EST同工酶参与肝脏的解毒功能相关。
本研究中大鳞鲃5种组织中肾脏LDH、MDH和EST同工酶表达活性都较强,反映在酶谱图上肾脏同工酶着色程度相对较深,LDH同工酶和EST同工酶不仅着色程度较深,而且具有相对较多的酶带数。肾脏是鱼类主要的排泄器官,对于类似大鳞鲃这种广盐性鱼类,在淡水中生活时主要依靠肾脏调节水分,以维持正常的渗透压,确保生理功能。大鳞鲃肾脏同工酶表达活性较强可能与其作为广盐性鱼类需要肾脏进行渗透压调节的功能密切相关。在其他广盐性鱼类的研究中也观察到这种现象[34]。
3.3 大鳞鲃同工酶的比较
关于鲃亚科鱼类同工酶已经有一些报道,大鳞鲃LDH、MDH和EST同工酶与已有研究结果的比较见表3,从表3可以看出鲃亚科不同属或不同种的鱼类同工酶存在差异,表现在即使相同组织的同一种同工酶在不同种鱼类中表达的酶带数不同,或者总存在有差异的同工酶,从这一角度可以印证同工酶可作为鉴定不同种鱼类的生化遗传指标,也就是说同工酶具有种属特异性,尤其对形态特征难以区分的不同种鱼类,可借助比较同工酶进一步进行种质鉴定。借助同工酶的种属特异性,已经运用在不少鱼类的种质鉴定中[35-37]。
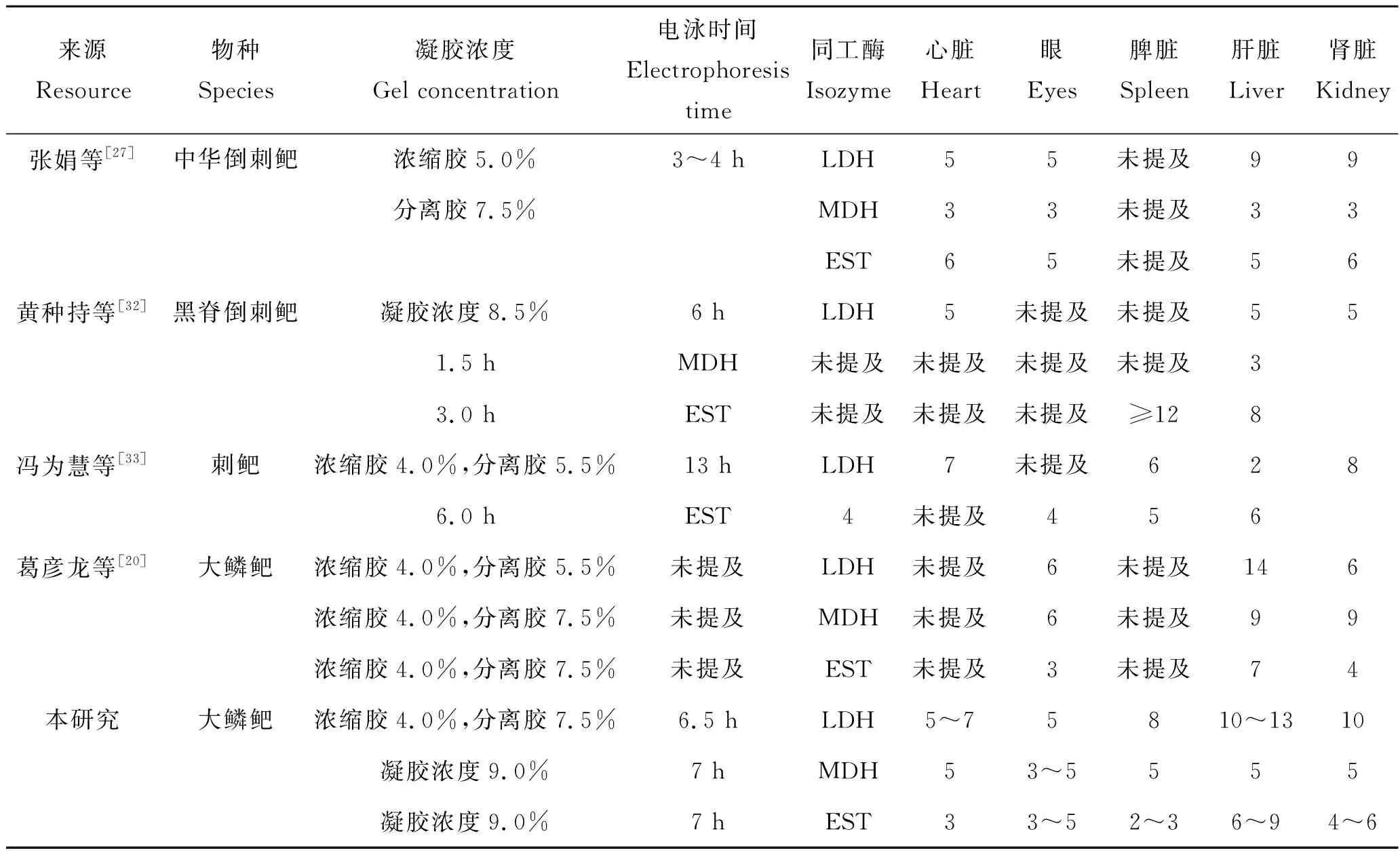
表3 大鳞鲃LDH、MDH和EST研究结果的比较Tab.3 Comparations of LDH, MDH and EST isozymes expressed in Barbus capito in different research results
关于大鳞鲃LDH、MDH和EST同工酶,本研究结果与已有研究报道[20]存在部分差异,分析认为造成这些差异的原因可能与实验方法不同有关,包括同工酶样品制备、凝胶浓度的筛选、电泳过程不同等。这种现象也见于其他鱼类的研究报道中[38]。
4 结论
本研究采用传统形态学方法观测了大鳞鲃的形态特征,通过外观描述、解剖观察内部构造特征,并结合其可数、可量性状进行了讨论分析。大鳞鲃的主要形态特征为:体呈梭形,头部较小,体修长,口亚下位,具有吻须和颌须各一对。背鳍起点位于腹鳍起点前端,尾鳍正尾形,背鳍起点至吻端距离比至尾鳍基距离短。身体被覆奇特发光的较大鳞片,鱼体背部呈银灰色,腹部银白色。鳍式为背鳍D.Ⅲ-7、胸鳍P.Ⅰ-16~18、腹鳍V.Ⅱ-8、臀鳍A.Ⅲ-5和尾鳍C. 24-28;侧线鳞数64-73、侧线上鳞数12~14、侧线下鳞数10~11;下咽齿齿式为2·3·5/5·3·2;脊椎骨数为46~49。同时,通过聚丙烯酰胺凝胶垂直板电泳对大鳞鲃心脏、眼睛晶状体、肝脏、肾脏和脾脏5种组织的LDH、MDH和EST同工酶进行了分析,大鳞鲃同工酶具有明显的组织特异性,其组织特异性与其生理功能密切相关,肝脏同工酶表达活性最强,肾脏同工酶较强的活性可能与其调节渗透压功能有关。本研究可为大鳞鲃种质鉴定、制定其种质标准提供参考依据。
猜你喜欢
海洋信息技术与应用(2022年1期)2022-06-05
中国实用医药(2021年7期)2021-04-16
宁夏医学杂志(2020年3期)2021-01-21
儿童时代·幸福宝宝(2020年9期)2020-09-08
中医眼耳鼻喉杂志(2019年2期)2019-04-13
金色少年(奇趣科普)(2016年8期)2016-09-21
中外医疗(2015年5期)2015-08-29
中国当代医药(2015年21期)2015-03-01
西南国防医药(2015年11期)2015-02-28
茶叶通讯(2014年2期)2014-02-27
