
番茄中Ty-1 抗性机制研究
2022-10-01 07:28黄毅
种子科技 2022年16期
黄 毅
(贵阳职业技术学院,贵州 贵阳 550081)
1 Ty-1 特性
番茄黄叶卷曲病毒(TYLCV)是世界上最具毁灭性的植物病毒之一。为了抵御这种病毒,一些抗性基因被用于育种中。植物基因组中存在的抗病基因通过产生R 蛋白,为植物抵抗病原体提供抗病能力。在这些基因中,Ty-1 抗性基因的应用最为广泛,许多商业杂交品种都携带Ty-1[1]。Ty-1 最早在1969 年智利南部发现,在最初的大规模重组筛选中首次提出了Ty-1 的简单图谱,如今已经鉴定并克隆了Ty-1 抗性基因。虽然Ty-1 不编码经典的NBS-LRR 基因,也不产生涉及超敏反应(HR),但Ty-1 影响病毒的复制或移动,含有该基因的植物表现出更多的耐受性。
RNA 沉默是一种基于序列同源性的基因调控机制,保留在真核生物中,不仅与调节基因表达有关,还抵御转座子等寄生遗传因素,并控制转基因和病毒[2]。RNA 沉默,也称为RNA 干扰(RNAi),通常被认为是植物和昆虫中的一种抗病毒防御机制,也被证明在哺乳动物中起到抗病毒作用。在RNAi 的机理过程中有4个关键部分:长双链RNA(dsRNA)、21-26 nt 短干扰RNA小链(siRNA)、内核糖核酸酶(Dicer)和RNA 诱导沉默复合物(RISC)(见图1)。

图1 RNAi 沉默机制
在植物中,RNA 病毒通过转录后基因沉默(PTGS)作为靶标,而像双子病毒的DNA 病毒则通过转录后基因沉默(TGS)作为靶标[3]。PTGS 和TGS 的抗性策略不同(见图2),TGS 通过细胞核中的DNA 甲基化与转座子的调节有关,而PTGS 通过细胞质中的长双链RNA调节病毒感染。在植物中,病毒和转基因都能通过PTG诱导基因沉默,这一过程被普遍认为是植物抵御某些病毒病原体感染的组成部分。
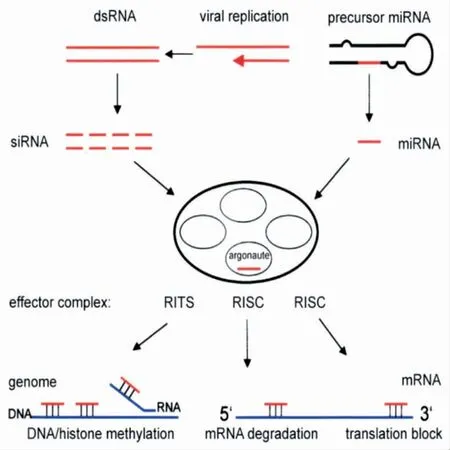
图2 基因沉默和转录后基因沉默模型
RNA 诱导DNA 甲基化(RdDM)已被植物用作对抗双子病毒的表观遗传防御[4](见图3)。在RdDM的过程中,涉及两种关键酶——RDR2 和DCL3。RDR2 是一种功能酶,可调节内源性植物siRNA 和病毒次级siRNA的生物合成,并在RdDM途径中靶向dsRNA 的转化。dsRNA 被消化成大小不同的siRNA,主要是24nt(由DCL3 产生)、21nt(由DCL4 产生)和22nt(由DCL2 产生)。不同大小的siRNA 具有不同的功能,例如,24-nt siRNA(与DCL3 相关)可能指导病毒DNA 甲基化和转录沉默。DCL3 是一种RNA 酶Ⅲ样酶,属于由4 个原型成员组成的家族,参与RDR 最后一步,以帮助24-nt siRNA 的发生。重要的是,DCL3 和24-nt siRNA 显然与染色质甲基化有关。
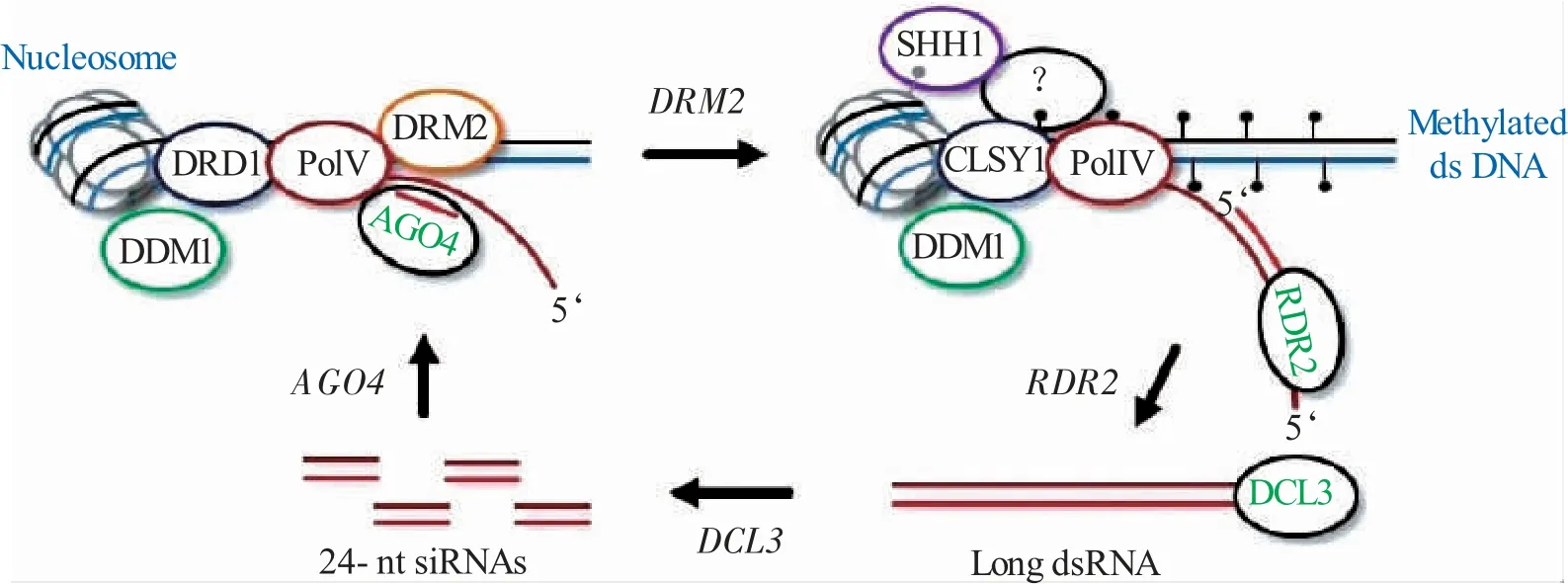
图3 RNA 诱导DNA 甲基化模型(Pooggin 2013)
2 研究目标
分析Ty-1 是否对其他双子病毒有抗性,Ty-1 番茄植株将受到BCTV 侵染,将其与易感MM系的含量进行比较分析。根据最新研究,Ty-1 不是TYLCV 的特异性,也可以对抗其他双子病毒甚至其他DNA 病毒,例如,Ty-1 似乎也能控制不同(双组分)双病毒物种的耐受性反应,如ToSRV。但是几乎没有关于Ty-1 对其他病毒的耐药性研究[5]。
3 试验方法
3.1 植物
番茄试样MM、Ty-1、Ty-2 的种子在土壤中,24 ℃和60%相对湿度的温室条件下,光照16 h、夜间8 h 养护3 周生长。烟草置于相同的温室条件下。在接种大约3~4 周后,将番茄试样的顶部叶片采摘并在-20 ℃下储存至使用。
3.2 TYLCV,BCTV,ToSRV 接种
用番茄黄叶卷曲病毒、甜菜卷曲顶病毒或番茄皱纹病毒对番茄品系进行接种。将含有该病毒克隆的根癌农杆菌(A.tumefaciens)转化并在3 mL LB3 培养基中与6 μL 卡那霉素和3 μL 利福平抗生素一起培养。液体培养物在28 ℃温度下以180 rpm 摇动过夜。第二天,将600 μL 过夜培养基转移到3mL 诱导培养基中,相同条件下培养。悬浮在3 mL MS-MES 培养基中,样品以3 500 rpm 离心20 min 后充分混合。测量OD600为0.5 时稀释。在温室条件下,用无针注射器加压5 mL接种,对3 周龄番茄植株进行渗透。
3.3 DNA 提取和PCR 分析
为了确认敏感番茄试样中是否存在BCTV 的DNA,设计BCTV 引物进行PCR 正向引物(5`-TGTAVT VVGATVGAGCGTGTT-3`)和反向引物(5`-TCATACAA CGAACACTTCATGA-3`),产物长度为700 bp。
从6 周幼苗中分离DNA,用液氮中研磨1 g 植物样后,与分离DNA 缓冲液混合。用Eppendorf 旋转约2 min 后,以12 000 rpm 的转速旋转5 min,将透明液体移到新管中。洗涤后用离心机以12 000 rpm 的速度去除多余液体。向每根试管中加入30 μL 的G.lig EB,等待3 min。试管在12 000 rpm 下旋转2 min 后准备进行PCR。使用1ul DNA 提取物作为模板,检查DNA 片段扩增。
4 结果
4.1 BCTV 克隆成功感染Nicotiana benthamiana
BCTV 对番茄品种和烟草影响不同。对于番茄而言,其会导致植株发育迟缓、变黄、叶脉膨胀变紫、向上卷曲叶片。其对烟草特有的症状是向下卷叶和发育迟缓。
在测试Ty-1 是否也具有对不同于TYLCV 的双子病毒的抗性之前,将BCTV 的一种PGV 3850 农杆菌克隆接种在本氏烟上,测试其传染性。PGV 3850 接种后13 d,本氏烟下叶出现黄变、坏死等症状。为了确认是否存在BCTV,采用特异性PCR 引物检测病毒的存在。结果显示预期大小为700 bp 的条带,说明设计的引物对扩增BCTV有用,进一步说明本生烟已被BCTV感染。
4.2 Ty-1 系列表现出对BCTV 的抗性
为测试Ty-1 是否也能抵抗其他双子病毒,用BCTV 克隆对Ty-1 番茄植株进行接种,以测定感染后的病毒含量,并与敏感MM进行比较。采用烟草响尾蛇病毒(TRV)的病毒诱导基因沉默(VIGS)方法构建了两个VIGS 结构,分别用于Solyc06g051180 的TRV2-180和Solyc06g051190 的TRV2-190。
由于Solyc06g051180 和Solyc06g051190 之间同源性较低,两个VIG 载体TRV2-180 和TRV2-190 是特异性的,并且都被假定为单个RDR。含有Ty-1 的植株通过TRV2-180 或TRV2-190VIG 进行侵染沉默,出现TYLCV 疾病症状并观察抗性受损。在用BCTV 感染之前,首先用TRV2-180 和TRV2-190 对Ty-1 植株进行接种,使VIGS 沉默Ty-1 并损害其抗性。作为对照,在Ty-1 植株上接种TYLCV。Ty-1 和MM在温室中播种,1 周后移栽到不同的花盆中,用TRV2-180 或TRV2-190进行接种。约3 周后,感染叶片表现为发育迟缓、变黄、叶脉肿胀、变紫以及向上卷曲的叶片(见图4)。在对Ty-1 进行VIG 沉默的MM和Ty-1 植株上,症状清晰可见,典型症状是感染TYLCV 的叶片变黄和卷曲。对于受到BCTV 攻击的Ty-1 和MM番茄植株,这些症状表明,Ty-1 的VIGS 沉默(TRV2-180 和TRV2-190)相对于未被VIGS 沉默的Ty-1 系表现出比BCTV 更明显的症状,如严重发育迟缓、向上卷曲叶片、发育缓慢。相比之下,在BCTV 侵染的Ty-1 株系上几乎看不到症状,而少数Ty-1 植株表现出与MM相似的症状。

图4 Ty-1 和MM 分别被TYLCV 或BCTV 侵染后的症状
4.3 扩增BCTV 片段并进行PCR 检测
为了检测感染的Ty-1 番茄叶片中BCTV 含量,采用qPCR 方法对RV2-180/190 VIGS 沉默的Ty-1 植株、Ty-1 植株和易感MM 番茄中的BCTV 滴度测定。分析前,设计了一套特定的引物,从这些感染的植物样本中PCR 扩增BCTV 片段,并获得约700 bp 的产物。作为对照,加入TYCLV 感染样本,并使用设计成功的引物进行测试,得到了300 bp 的产物。在被侵染的植物中用PCR 检测BCTV 的DNA(见图5),结果显示,MM 株系和TRV2-180/190 VIGS 沉默的Ty-1 番茄植株样本分别呈现预期的DNA 产物,其中TYLCV 为300 bp,BCTV 为700 bp。由此可以证明MM 系和TRV2-180/190 VIGS 沉默的Ty-1 番茄植株系已感染BCTV。使用BCTV 引物对所有试验植物的分离样本进行PCR,以扩增BCTV,并证明所有MM 系和TRV2-180 VIGS 沉默的Ty-1 系均成功感染BCTV。但结果显示,其中一个Ty-1 样本也感染了BCTV。

图5 在被侵染的植物中用PCR 检测BCTV 的DNA
4.4 用qPCR 测定BCTV 有效量
BCTV 的侵染成功和PCR 引物的确定后,分别对Ty-1 植株、感病MM番茄和TRV2-180/190 VIGS 沉默的Ty-1 番茄叶片样本进行qPCR 检测,以确定BCTV滴度。内部对照,在Butterbach 等人的TYLCV 试验的Ty-1 的qPCR 分析中使用了GFP 基因的Ty-1 进行的qPCR 分析相同。
根据qPCR 结果,在样本中观察到不同的BCTV病毒滴度(见图6)。正如预期,从易感MM采集的大多数植物样本中普遍观察到较高的滴度,而在Ty-1 中观察到较低的滴度,但样本除外。由于Ty-1 的VIGS沉默在早期损害了对TYLCV 的抗性,因此经类似处理(TRV2-180 或TRV2-190)并随后用BCTV 激发的Ty-1 植株显示出比Ty-1 更高的滴度以及与MM相似的滴度水平,并进一步表明Ty-1 的VIGS 沉默损害了对BCTV 的抗性,结果表明Ty-1 对BCTV 具有抗性。虽然Ty-1 株系的平均滴度低于对照组,但一个Ty-1株系样本显示出比其他Ty-1 株系更高的含量(见图7)。
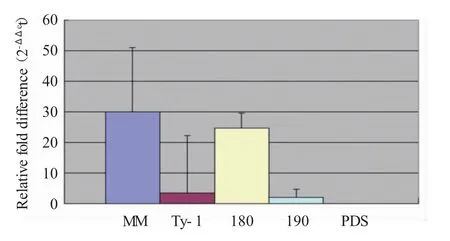
图6 qPCR 分析BCTV 分别侵染MM、Ty-1 lines、TRV2-180 和190 VIGS-silenced Ty-1

图7 2-ΔΔCt 方法分析qPCR 结果
5 讨论
试验植物表现出了预期症状,BCTV 的PCR 结果与阳性对照(TYLCV)相同,从而证明MM、TRV2-180或TRV2-190 VIGS 沉默的Ty-1 系感染了BCTV。通过qPCR 方法在Ty-1 系中产生的较低滴度,在大多数易感MM和TRV2-180 VIGS 沉默的Ty-1 系中产生的滴度较高,综上结果Ty-1 确实对BCTV 产生了抗性。
另外,观察到一株被BCTV 攻击的Ty-1 番茄植株表现出易感MM 番茄的症状,这株Ty-1 番茄植株被BCTV 感染,产生了更高的滴度。结果表明该Ty-1 番茄植株不对抗BCTV,Ty-1 通过增强TGS 而产生抗性。可能Ty-1 番茄植株不能产生足够的siRNA 信号,会导致TGS 抑制和RNAi 沉默失败。目前尚不清楚这种新型Ty-1 植物产生的原因和机制,因此在不久的将来对其进行研究将是一个挑战。
猜你喜欢
当代水产(2022年7期)2022-09-20
当代水产(2022年1期)2022-04-26
西南农业学报(2022年3期)2022-04-25
昆明医科大学学报(2022年2期)2022-03-29
特别健康·下半月(2020年12期)2020-12-14
中国瓜菜(2019年8期)2019-09-19
植物保护(2019年2期)2019-07-23
江苏农业科学(2016年11期)2017-03-21
江苏农业科学(2017年1期)2017-02-27
江苏农业科学(2016年4期)2016-06-14
