
大气CO2浓度升高对高、低响应水稻品种根系生长的影响——FACE研究①
2022-09-27 01:36:06王东明朱春梧
土壤 2022年4期
陶 冶,蔡 创,韦 薇,杨 雄,王东明,沈 敏,宋 练,朱春梧*
大气CO2浓度升高对高、低响应水稻品种根系生长的影响——FACE研究①
陶 冶1,2,蔡 创1,韦 薇1,2,杨 雄1,2,王东明1,2,沈 敏1,2,宋 练1,朱春梧1*
(1 土壤与农业可持续发展国家重点实验室(中国科学院南京土壤研究所),南京 210008;2 中国科学院大学,北京 100049)
大气CO2浓度([CO2])升高是未来气候变化的主要特征之一,研究表明高[CO2]会提高水稻产量,且水稻两个亚型(籼稻和粳稻)的产量增幅存在很大的差异。然而,目前还不明确这种基因型的响应差异是否与其根系的生长响应差异有关。本试验依托自由大气[CO2]升高研究平台(free air CO2enrichment, FACE),以低产量响应品种粳稻 (L.) 武运粳23和高产量响应品种籼稻 (L.) 扬稻6号为试验材料,设置高[CO2] (590 μmol/mol) 和对照(390 μmol/mol)处理。使用微根窗法连续动态检测两种水稻的根系生长状况,进一步计算根系周转率指标。结果表明:高[CO2]显著促进了两个品种的水稻根系生长,其根系长度、表面积和体积相对于对照平均增加了120%、106% 和98%;两个品种间,高[CO2]处理使武运粳23的根系周转率显著下降,降幅达66%;扬稻6号的根系周转率则上升了约18%;高[CO2]处理下的武运粳23的根系增量主要分布在浅层(32% 升至56%),而扬稻6号的根系在深层的分布从12% 增加至20%。结合已有产量响应及氮素吸收结果,我们认为相较于根长等根系形态指标,水稻根系的周转率与深层根系分布比例可能是不同基因型水稻品种的产量对高[CO2]响应存在明显差异的主要原因。
CO2浓度升高;水稻;根系生长;高低应答品种;微根窗
受人类活动的影响,大气CO2浓度([CO2]) 大概率将持续增加,到21世纪末最高可达1 135 μmol/mol (SSP5-8.5)[1]。水稻作为全球超过一半人口的主粮[2],其生产对高[CO2]的适应性关乎未来粮食安全[3-4]。许多研究表明高[CO2]会促进水稻产量,且产量增幅在品种间有较大的差异[5-9],许多研究从光合响应、产量形成等方面探明水稻产量响应差异的机理[5-6, 10-11],却很少有研究从根系生长角度进行解释。水稻根系是支撑地上部、吸收水分和养分的重要器官[12],根系的生长发育与地上部生长、产量与品质的形成密切相关。因此,监测根系生长对高[CO2]的响应在研究水稻对全球变化的适应性上具有重要意义。
由于根系生长在地下的特性,长期以来,水稻根系研究多采用挖掘法、根钻法、容器法等传统方法[13],这些传统方法的缺点是取样具有破坏性,不仅工作量大,且无法对特定根系进行连续动态的观测。微根窗 (minirhizotrons) 法是一种原位的、非破坏性和可定点持续观测植物根系的方法,此方法不仅可以无损地对特定时期根数、根长和根表面积等形态指标的相对量进行测定,其主要优势在于可以连续、定点地观测特定根系或根系片段在整个生长季的动态变化。不仅如此,通过分析根管视野内老根的衰老和新根的产生,微根窗法还可获得根系的更替状态,研究中通常使用根系周转率进行表示。根系周转率不仅可以表达根系新老更替的速率,还可以反映植物本身的动态资源分配。因为植物发新根消耗的能量要远高于保持相同量的老根所需的能量[14],较高的根系周转率表示较多的新根产生,可以维持较高的根系活力[15]。因此,许多研究也发现根系周转率相对于静态的根系形态指标对全球变化因子(如CO2、温度升高)具有更强的敏感性,可以很好地表征植物根系对环境变化的响应[15-17]。
已有的研究表明高[CO2]会使水稻根系生物量显著增加,对不定根数、不定根总长度、根系体积等形态指标也有促进作用[18-19],但对水稻生长季内根系周转率的影响鲜有报道,也未从根系周转、深层分布等角度研究高低响应品种的差异。因此,本研究依托自由大气[CO2]升高研究平台(free air CO2enrichment, FACE),以低产量响应品种粳稻 (L.) 武运粳23和高产量响应品种籼稻 (L.) 扬稻6号为试验材料,设置高[CO2] (590 μmol/mol)和对照 (390 μmol/mol)处理。使用微根窗法连续动态检测两种水稻的根系生长发育状况,进一步计算根系生长动态指标,以期阐明高[CO2]对水稻根系形态指标、根系周转率的影响以及其基因型差异,为全球变化条件下水稻根系调控、品种选育及制定水稻生产对大气[CO2]升高的应对策略提供参考依据。
1 材料与方法
1.1 试验平台信息
FACE平台位于江苏省苏州市常熟市古里镇康博村(31°30′N, 120°33′E),于2010年建设完成。该处的海拔高度为6 m,属于典型的亚热带季风气候,其年降水量1 100 ~ 1 200 mm,年均温度16 ℃,年均日照时长>2 100 h,无霜期>200 d。该区土壤类型为乌栅土,耕作方式为水稻–冬小麦轮作。耕层土壤基本理化性质为:有机碳16 g/kg,全氮1.9 g/kg,有效磷12.3mg/kg,速效钾94.3mg/kg。试验共设有两个[CO2]处理,分别为390 (Ambient CO2,记为Amb.)和590 μmol/mol(Elevated CO2,记为E[CO2]),每个处理设3个重复,共计6个试验圈,其中各试验圈均为正八边形,直径8 m,面积约50 m2。FACE圈内CO2放气管道距水稻冠层50 cm左右,系统根据风速、风向来调整FACE圈CO2气体的喷施。FACE平台的设计原理、运行模式和性能等详见Cai等[5]。
1.2 试验设置与栽培管理
试验于2020年水稻季进行,供试品种为低产量响应品种武运粳23 (WYG23)和高产量响应品种扬稻6号(YD6)[8]。5月27日播种,6月23日移栽,武运粳23和扬稻6号的收获日期分别为10月15日和10月18日。CO2喷施处理始于6月24日,止于10月10日。氮、磷、钾肥的施用量分别为18.1、6.9和6.9 g/m2,其中磷、钾肥作为基肥一次性全部施入,氮肥基肥、分蘖肥和穗肥的量分别是6.9、6.0和5.2 g/m2。基肥采用氮磷钾比例为15︰15︰15的复合肥于移栽前一天施入,于7月4日和8月9日分别追施分蘖肥和穗肥。武运粳23每穴2棵苗,扬稻6号每穴1棵苗,种植密度均为24穴/m2。生育期内(5月27日—10月18日)最高、最低及平均温度分别为29.1、22.0 和25.1℃,降水量为571.4 mm。
1.3 微根窗法根系测量
根管安装:于水稻移栽后封行前(7月4日)埋入根管。考虑水稻根系分布较浅特性,同时为了保证留有足够长度使根管上口高于稻田的水层,于水稻两行间以与地面呈45°角埋入1 m长根管,控制深度,使得埋入土壤中部分和露出部分分别为70 cm和30 cm,露出部分包裹遮光材料,根管顶部使用塑胶盖密封。由于稻田保有水层,于根管上悬挂重物保证根管不因浮力发生移动。每个品种在一个试验圈内埋设2根根管,共24根。
根系扫描:根系生长在一天内的变化几乎可以忽略,为了避免清晨露水影响仪器,试验于下午时段(14:00—19:00)进行根系扫描。首先将原位根系扫描仪(CI-600, CID Inc., Camas, WA, USA) 放入自带白色根管中进行校正,随后连接便携式电脑于田间放入根管进行扫描,每根根管扫描得到3张不同深度的根系图像。抽穗前扫描时间为8月11日,抽穗期扫描时间为9月4日,灌浆期扫描时间为9月21日、10月1日、10月10日、10月17日(其中10月17日武运粳23已收获,未扫描)。
图像分析:使用RootSnap (CID Bio-Science, Inc., Camas, WA, USA) 软件对扫描得到的图像进行整理,描绘根系和分析。根系周转率选用根长指标进行计算,本文计算了8月11日—9月4日(阶段1)和9月4日—9月21日(阶段2)之间的根系周转率。前(时期1)后(时期2)两个时期内的根系周转率使用以下公式进行计算:

1.4 数据处理
使用Microsoft Excel 2016对数据进行整理与初步分析,使用Statistix 9对数据进行统计分析,利用Sigma Plot 14软件进行作图,使用LSD (least significant difference)法在0.05水平进行显著性水平检验。
2 结果与分析
2.1 [CO2]升高对水稻根系形态的影响
2.1.1 总根长 总体来说,武运粳23和扬稻6号的总根长均在齐穗前达到最大值(图1A)。就处理而言,与对照相比,高[CO2]下的两个水稻品种的根长均显著增加,具体表现为:FACE处理使武运粳23的总根长平均增加了170%,使扬稻6号的总根长平均增加了70%。然而,武运粳23和扬稻6号的总根长在高[CO2]下增加的比值随着生育期的推进而明显降低,表明高[CO2]加速了两个品种根系的衰老。而且,高[CO2]加速武运粳23根系衰老的速度要明显快于扬稻6号。
2.1.2 根表面积 [CO2]升高对两种水稻品种根表面积的影响如图1B所示,根表面积指标在两个品种及处理间的变化趋势与根长表现相似。就处理而言,高[CO2]处理使得两个水稻品种的根表面积相比于对照均显著增加,具体表现为:高[CO2]处理使武运粳23的根表面积平均增加153%,扬稻6号平均增加了59%。同样,高[CO2]下武运粳23和扬稻6号的根表面积增幅随着生育期的推进而明显降低。
2.1.3 根体积 [CO2]升高处理下,两个品种在测量期间的根体积见图1C。总体来说,高[CO2]显著增加了两个水稻品种的根体积,具体表现为:高[CO2]下武运粳23的根体积平均增加了138%,扬稻6号的根体积平均增加58%。
2.2 [CO2]升高对水稻根系周转率及根系分层分布的影响
2.2.1 根系周转率 武运粳23和扬稻6号的根系周转率对高[CO2]的响应如图2A所示。在阶段1中,对照环境下,两个品种的根系周转率无显著差异;而高[CO2]处理对两个品种周转率的影响则不同,FACE处理使武运粳23的根系周转率显著下降,降幅达66%;而对扬稻6号的根系周转率则有促进作用(+18 %),但未达到显著水平。在阶段2中,两品种整体的根系周转率均有明显的下降,同时,FACE处理降低武运粳23而提高扬稻6号根系周转率的趋势保持不变,均未达到显著水平,且作用的幅度小于阶段1。
2.2.2 根系分层分布 [CO2]升高对两个水稻品种根系在地下分布的影响如图2B所示。在对照环境下,武运粳23和扬稻6号的根系分布无显著差异,均是中层分布最多(54% 和50%),浅层次之(32% 和38%),深层最少(14% 和12%)。在高[CO2]处理下,两个品种的根系分布比例均发生了变化,武运粳23的根系分布在FACE处理下改变为中层和深层分布比例降低,而浅层根系分布例提高了66.7%;扬稻6号的根系在浅层的分布比例变化不大(38% ~ 42%),中层根系分布整体降低了12%,深层的根系分布则从12% 提升至20%。
3 讨论
与地上部生物量、产量等指标对高[CO2]响应一致,已有的试验表明大气[CO2]升高同样会使水稻各生育期根系生物量、根长和根体积等形态指标显著高于对照[18-27]。刘红江等[26]和杨洪建等[27]分别以杂交籼稻籼优63和粳稻武香粳 14为试验材料,发现FACE处理使其抽穗期根系干重分别增加了50.2% 和38.3%。以根长指标为例,在本研究中,高[CO2]处理使武运粳23和扬稻6号的根长在齐穗期分别增加了170% 和70%,这与前人的结果基本一致。高[CO2]环境下,供试品种根长等形态指标的提升与产量增加密切相关。
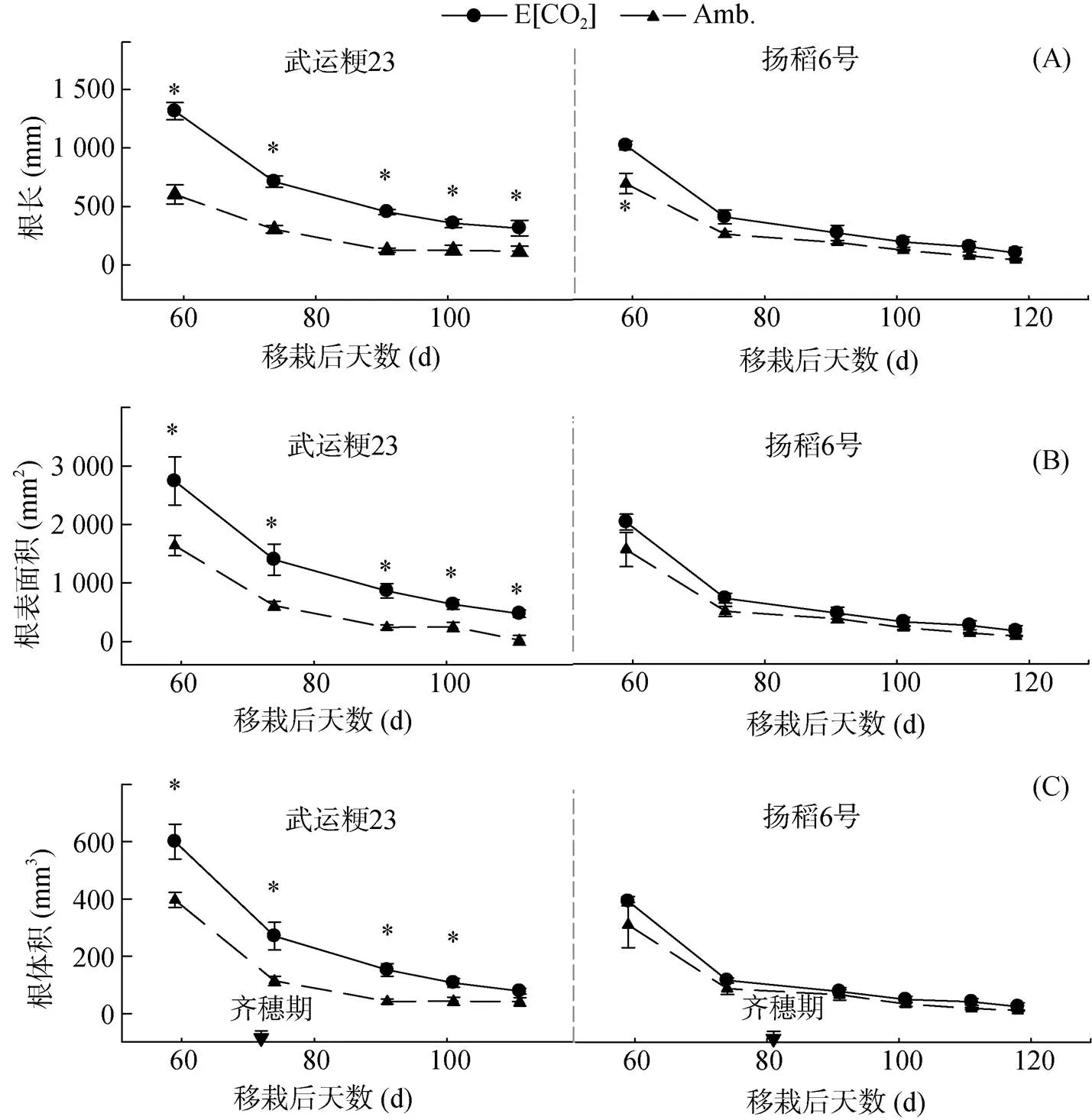
(图中*表示两处理间差异达P<0.05显著水平;Amb.:390 μmol/mol;E[CO2]:590 μmol/mol;下图同)
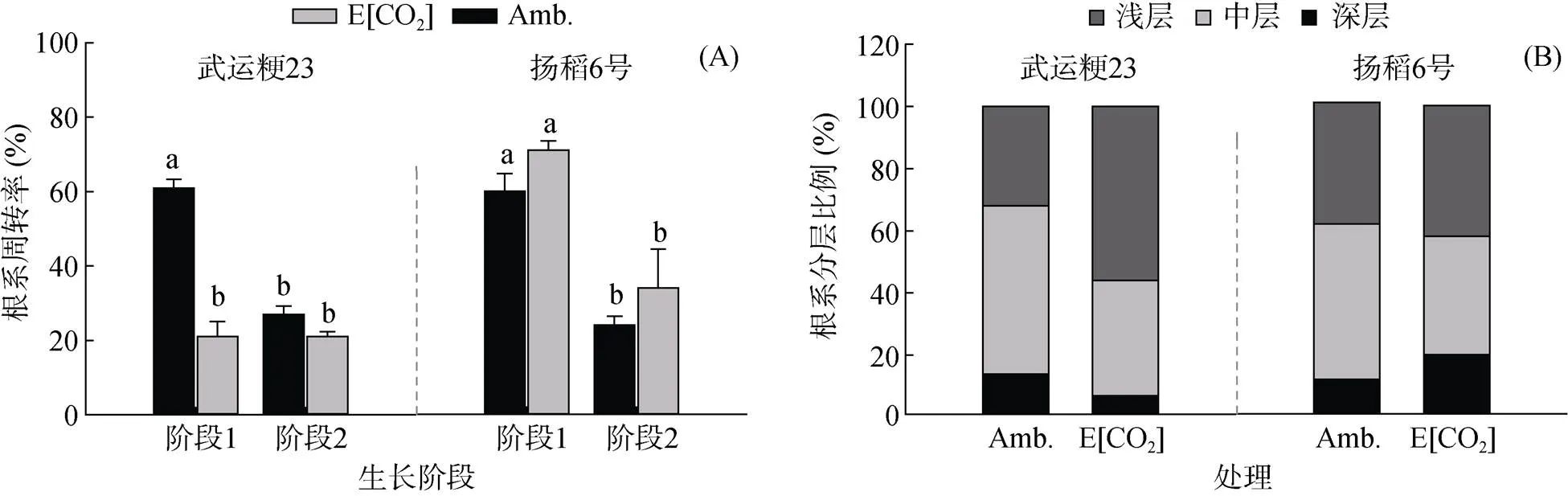
(阶段1和阶段2分别为8月11日—9月4日和9月4日—9月21日,图中小写字母不同表示同一水稻品种不同处理间差异达P<0.05 (LSD)显著水平,下图同。)
受限于水稻一年生特性和传统研究方法的限制,破坏性的根系取样很难获得根系周转率相关的数据,已有的有限的使用微根窗法有关水稻的研究也并未展示相关数据[20-25]。本研究对比前3次的测量结果,计算出武运粳23和扬稻6号在该段时间内的根系周转率。这3次测量时间覆盖了两个品种整个开花期。与根长等形态指标不同,本试验选用的两个品种在根系周转率的变化趋势上表现出了品种间的差异。在对照情况下两个品种的周转率无显著差异的情况下,阶段1中高[CO2]处理使武运粳23的根系周转率显著下降,降幅达66%;扬稻6号的根系周转率则上升了约18%。阶段2中两品种根系周转率对高[CO2]处理的响应保持了与前一阶段相同的趋势,但由于生长后期根系衰老,整体数值下降明显。以往的研究已表明,较高的根系周转率意味着根系可以保持较高的根系活力,有利于根系吸收营养物质[15-16]。同样的,有关根系分布与产量关系的研究表明,浅层根的质量与产量之间无显著的相关关系,而深层根质量与产量呈显著正相关[28]。在本研究中,在高[CO2]环境下,武运粳23的根系增量主要分布在浅层土壤中,而扬稻6号深层土壤中的根系分布明显增加。在以往的高[CO2]研究中,籼稻品种产量增幅的表现要普遍高于粳稻[8],本试验的结果也表明扬稻6号的地上部氮吸收量及增幅均高于武运粳23(图3A)。综合本研究所测指标,在FACE条件下,扬稻6号在根长等形态指标上,即使以单茎为单位相对于武运粳23也未有优势(图3B)。而在根系周转率和根系的深层分布上优于武运粳23。因此,我们认为相较于根长等根系形态指标,水稻根系的周转率与深层根系分布比例与水稻对高[CO2]产量响应差异有着更为密切的关系。另一方面,根系周转过程中的碳输入是陆地生态系统向土壤有机碳输入的主要贡献者[15]。本研究表明,在高[CO2]环境下,高产量响应品种根系周转率显著高于低响应品种,在应对全球变化中,高响应品种不仅在产量上具有优势[8],在对土壤有机碳输入方面同样具有较大的潜力。
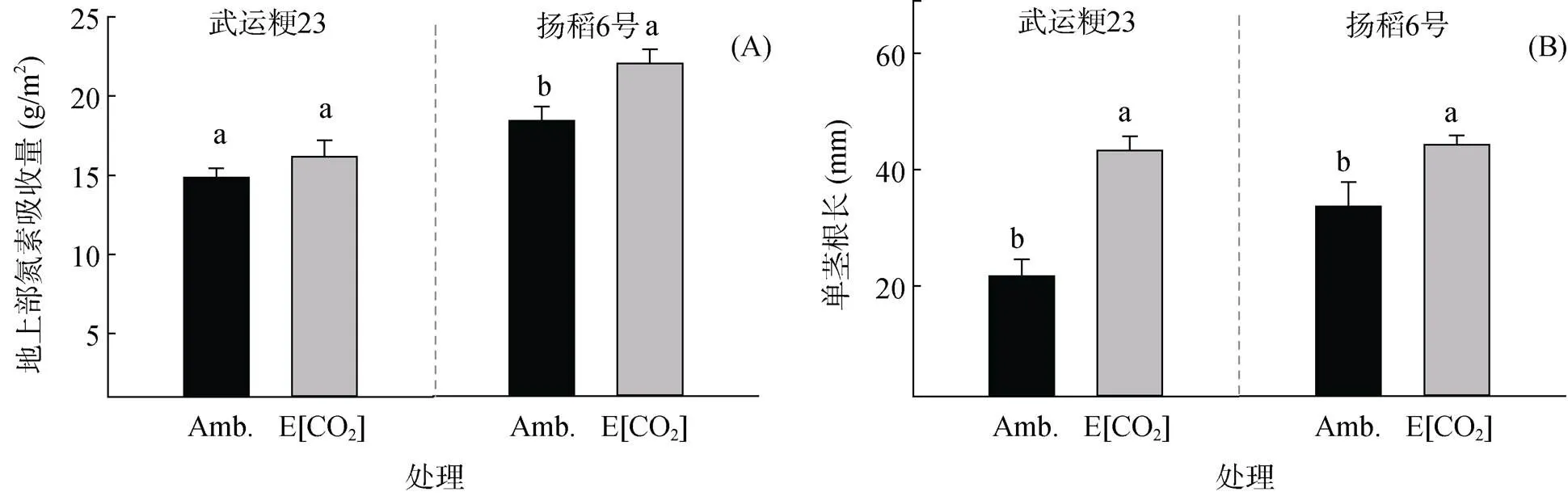
图3 大气[CO2]升高对武运粳23和扬稻6号地上部氮素吸收量(A) 和单茎根长(B)的影响
微根窗法在安装稳定后可以非常方便地动态监测根系的生长,获得根的颜色、分支特征、根系衰亡、分解等诸多信息。诚然,微根窗法本身也具有一定的局限性。首先,此方法的观测是以单个根管为单位,无法测得通用的以土地面积为单位的根系形态指标,且无法直接获取到根系样品进行后续生化分析;其次,水稻是须根系作物[12],根系主要分布在浅层土壤中,而根管的直径从浅层至深层则是一致的,这造成了不同深度根系被根管观察到的概率是不同的,其中较浅层的根系更易被根管监测到,浅层根系的生长变化也更易被根管捕获。上述微根窗法的特性决定此方法能够很好地适用于比较处理间的差异,而对品种间的差异,还需考虑品种间根系生长及分布特性并配合根钻法等直接取样的方法来做进一步的验证。武运粳23在FACE下根量较大根系周转率却较低以及不同于前人研究结果[24, 26-27]的高[CO2]对两个品种间的增幅差异也可能与微根窗法的特性有关。此外,武运粳23在FACE下根系周转率较低的另一个可能原因是根系过早的衰老,武运粳23的齐穗期及成熟期均早于扬稻6号,衰老过程中较低的新生根量也一定程度上导致了武运粳23较低的根系周转率。尽管微根窗法具有一定的局限性,但依然为研究水稻根系生长提供了一种原位、非破坏性并且可连续监测的方法,为研究大气[CO2]升高或其他环境因子的变化对水稻根系生长影响提供便利。
4 结论
高[CO2]显著促进了供试品种水稻根系的生长,使根系长度、表面积和体积相对于对照平均增加了120%、106% 和98%,这与其产量和地上部氮素吸收量在高[CO2]下明显增加有关。就品种间差异而言,在FACE条件下,扬稻6号在根长等形态指标上相对于武运粳23未有优势、而在根系周转率和根系的深层分布上优于武运粳23。结合扬稻6号在高[CO2]下的产量和地上部氮素吸收量的增幅均高于武运粳23,我们认为相较于根长等根系形态指标,水稻根系的周转率与深层根系分布比例的差异可能是水稻产量及氮素吸收对高[CO2]产量响应差异的主要原因。
[1] Meinshausen M, Nicholls Z R J, Lewis J, et al. The shared socio-economic pathway (SSP) greenhouse gas concentrations and their extensions to 2500[J]. Geoscientific Model Development, 2020, 13(8): 3571–3605.
[2] Fukagawa N K, Ziska L H. Rice: Importance for global nutrition[J]. Journal of nutritional science and vitaminology, 2019, 65(Supplement): S2-S3.
[3] Rosenzweig C, Elliott J, Deryng D, et al. Assessing agricultural risks of climate change in the 21st century in a global gridded crop model intercomparison[J]. PNAS, 2014, 111(9): 3268–3273.
[4] Rosenzweig C, Parry M L. Potential impact of climate change on world food supply[J]. Nature, 1994, 367(6459): 133–138.
[5] Cai C, Yin X Y, He S Q, et al. Responses of wheat and rice to factorial combinations of ambient and elevated CO2and temperature in FACE experiments[J]. Global Change Biology, 2016, 22(2): 856–874.
[6] Hasegawa T, Sakai H, Tokida T, et al. Rice cultivar responses to elevated CO2at two free-air CO2enrichment (FACE) sites in Japan[J]. Functional Plant Biology: FPB, 2013, 40(2): 148–159.
[7] Shimono H, Okada M, Yamakawa Y, et al. Rice yield enhancement by elevated CO2is reduced in cool weather[J]. Global Change Biology, 2008, 14(2): 276–284.
[8] Zhu C W, Xu X, Wang D, et al. An indica rice genotype showed a similar yield enhancement to that of hybrid rice under free air carbon dioxide enrichment[J]. Scientific Reports, 2015, 5: 12719.
[9] Zhu C W, Zhu J G, Cao J, et al. Biochemical and molecular characteristics of leaf photosynthesis and relative seed yield of two contrasting rice cultivars in response to elevated CO2[J]. Journal of Experimental Botany, 2014, 65(20): 6049–6056.
[10] Hasegawa T, Sakai H, Tokida T, et al. A high-yielding rice cultivar “takanari” shows no N constraints on CO2fertilization[J]. Frontiers in Plant Science, 2019, 10: 361.
[11] Yang L X, Huang J Y, Yang H J, et al. The impact of free-air CO2enrichment (FACE) and N supply on yield formation of rice crops with large panicle[J]. Field Crops Research, 2006, 98(2/3): 141–150.
[12] Yoshida S. Fundamentals of rice crop science[C]. Manila: International Rice Research Institute, 1981.
[13] 张福锁, 申建波, 冯固. 根际生态学—过程与调控[M]. 北京: 中国农业大学出版社, 2009.
[14] Canadell J G, Pitelka L F, Ingram J S I. The effects of elevated[CO2]on plant-soil carbon below-ground: A summary and synthesis[J]. Plant and Soil, 1995, 187(2): 391–400.
[15] Bader M, Hiltbrunner E, Körner C. Fine root responses of mature deciduous forest trees to free air carbon dioxide enrichment (FACE)[J]. Functional Ecology, 2009, 23(5): 913–921.
[16] Norby R J, Ledford J, Reilly C D, et al. Fine-root production dominates response of a deciduous forest to atmospheric CO2enrichment[J]. PNAS, 2004, 101(26): 9689–9693.
[17] 倪惠菁, 苏文会, 范少辉, 等. 养分输入方式对森林生态系统土壤养分循环的影响研究进展[J]. 生态学杂志, 2019, 38(3): 863–872.
[18] 沙霖楠. CO2浓度和温度升高对水稻根系生长的影响[D]. 南京: 南京林业大学, 2015.
[19] 许高平. 水稻根系发育对大气CO2浓度升高响应的基因型差异-FACE研究[D]. 扬州: 扬州大学, 2011.
[20] Lou Y S, Inubushi K, Mizuno T, et al. CH4emission with differences in atmospheric CO2enrichment and rice cultivars in a Japanese paddy soil[J]. Global Change Biology, 2008, 14(11): 2678–2687.
[21] Sun C M, Liu T, Guo D D, et al. Numerical simulation of root growth dynamics of CO2-enriched hybrid rice cultivar shanyou 63 under fully open-air field conditions[J]. Journal of Integrative Agriculture, 2013, 12(5): 781–787.
[22] 陈改苹, 朱建国, 谢祖彬, 等. 开放式空气 CO2浓度升高对水稻根系形态的影响[J]. 生态环境, 2005, 14(4): 503–507.
[23] 牛耀芳, 宗晓波, 都韶婷, 等. 大气CO2浓度升高对植物根系形态的影响及其调控机理[J]. 植物营养与肥料学报, 2011, 17(1): 240–246.
[24] 陶文辉, 王丹. CO2浓度升高对籼、粳稻光合生理和根系生长的影响[J]. 南京农业大学学报, 2021, 44(1): 27–35.
[25] 武慧斌, 宋正国, 沈跃, 等. 水稻根系生长发育对CO2浓度升高的响应及其品种间的差异[J]. 生态环境学报, 2014, 23(3): 439–443.
[26] 刘红江, 杨连新, 黄建晔, 等. FACE对三系杂交籼稻汕优63根系活性影响的研究[J]. 农业环境科学学报, 2009, 28(1): 15–20.
[27] 杨洪建, 杨连新, 刘红江, 等. FACE对武香粳14根系生长动态的影响[J]. 作物学报, 2005, 31(12): 1628–1633.
[28] 凌启鸿, 凌励. 水稻不同层次根系的功能及对产量形成作用的研究[J]. 中国农业科学, 1984, 17(5): 3–11.
Effects of Elevated Atmospheric CO2Concentration on Root Growth of “Strong” and “Weak” Response Rice Varieties:A FACE Study
TAO Ye1,2, CAI Chuang1, WEI Wei1,2, YANG Xiong1,2, WANG Dongming1,2, SHEN Min1,2, SONG Lian1, ZHU Chunwu1*
(1 State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China; 2 University of Chinese Academy of Sciences, Beijing 100049, China)
As one of the main scenarios of future climate change, studies have shown that high atmospheric concentration CO2([CO2]) will promote rice yield and the yield increase varies greatly between two rice subspecies (and). However, there is still a knowledge gap on whether the yield response difference between subspecies is related to root growth. Here we conducted a FACE (free air CO2enrichment, FACE) experiment using “weak” (L., Wuyungeng 23) and “strong” (L., Yangdao 6) yield responsive varieties, elevated [CO2] (590 μmol/mol) and control (390 μmol/mol) treatments were set. Minirhizotrons were used to detect the root growth status of the two varieties continuously and dynamically, and the root turnover rate was further calculated. The results showed that elevated [CO2] significantly enhanced the growth of rice roots of the two varieties, and the root length, surface area and volume across two varieties were increased by 120%, 106% and 98% on average compared to the control, respectively. The root turnover rate of Wuyungeng 23 was significantly reduced by 66% under elevated [CO2], the root turnover rate of Yangdao 6 increased by around 18%. For Wuyungeng 23, most of the root increased by elevated [CO2] was mainly distributed in the shallow layer (32% to 56%), however, the root distribution of Yangdao 6 in the deep layer was increased from 12% to 20% by elevated [CO2]. Combination with the yield response and aboveground nitrogen uptake, we concluded that compared to root morphological indicators such as root length, the turnover rate of rice root system and the distribution ratio of deep root system may be the main reason for the obvious difference in the response ofandrice to elevated [CO2].
Elevated CO2; Rice; Root growth; Strong and weak response varieties; Minirhizontrons
S511
A
10.13758/j.cnki.tr.2022.04.014
陶冶, 蔡创, 韦薇, 等. 大气CO2浓度升高对高、低响应水稻品种根系生长的影响—FACE研究. 土壤, 2022, 54(4): 763–768.
国家自然科学基金青年科学基金项目(32001191,31800359)和江苏省自然科学基金项目(BE2018402)资助。
(cwzhu@issas.ac.cn)
陶冶(1992—),男,安徽淮南人,博士研究生,主要研究气候变化对作物产量的影响。E-mail: taoyetoy@gmail.com
猜你喜欢
小小说月刊·下半月(2021年8期)2021-09-05 05:49:31
环渤海经济瞭望(2021年10期)2021-03-13 13:21:10
动漫界·幼教365(大班)(2018年3期)2018-05-14 11:50:46
农民致富之友(2018年22期)2018-01-29 12:46:18
上海农业科技(2017年3期)2017-07-18 11:47:49
山东农业科学(2017年4期)2017-05-18 17:06:13
上海农业科技(2017年1期)2017-03-08 07:39:48
现代经济信息(2016年15期)2016-07-07 13:57:15
财税月刊(2016年4期)2016-07-04 22:58:24
山东农业科学(2014年2期)2014-03-24 12:05:43
