
黄河三角洲柽柳灌丛对周边土壤呼吸的影响研究
2022-09-27 01:58马笑丹刘加珍陈永金马荣贞徐梦辰
地球环境学报 2022年4期
马笑丹,刘加珍*,陈永金,马荣贞,徐梦辰
1. 聊城大学 地理与环境学院,聊城 252059
2. 山东高速工程建设集团有限公司第六工程公司,济南 250101
3. 济南市环境研究院,济南 250101
土壤是陆地表层最大的碳储库(Bertora et al,2009),其与大气中的CO2交换过程被称为土壤呼吸。土壤呼吸是土壤库向大气库输出的主要途径(王丰川等,2013),其微小的变化也会对大气CO2浓度产生深远影响(Rodeghiero and Cescatti,2008),因此,土壤呼吸的研究已成为碳循环及气候变化的核心问题(Schlesinger and Andrews,2000;熊晓虎 等,2016)。植 被覆盖、根系分布、环境温、湿度和土壤特性的异质性都会导致土壤呼吸作用的空间异质性(韩广轩和周广胜,2009)。温度和湿度是影响土壤呼吸速率变化的重要物理因素(孟祥利等,2009;陈亮等,2016;王丹等,2016),温度主要通过促进或抑制酶活性、影响微生物活动和植被根的生长等来调控土壤呼吸(范洪旺等,2018),有研究表明:当温度超过15℃时,其他因素对土壤呼吸影响就会增加,超过35℃时,原生质可能开始降解、土壤呼吸速率减弱(Tang et al,2005)。干湿交替对土壤呼吸有明显的激发效应(孟伟庆等,2015),一般湿润季的土壤呼吸高于旱季(Bae et al,2013),土层上表现为5 cm土壤的含水量与土壤呼吸显著线性相关(张蔷等,2017)。
植物作为碳循环的生物库之一,除了自身是碳贮存库之外,还能通过影响其生存环境的温、湿度条件而影响土壤呼吸。在黄河三角洲,柽柳是区域木本植物的优势种,沃土、耐盐能力较强,成为滨海沿岸盐碱区及内陆低洼盐渍区主要的生态修复物种(李永涛等,2018)。柽柳的生长能够促使土壤养分不断聚集于灌丛周围而使立地周围土壤质地发生变化;从而增强柽柳对贫瘠盐碱地生态环境的适应性,促进柽柳的扩张(刘亚琦等,2017)。从固碳角度看,柽柳不仅通过植株固碳对区域碳循环起到积极作用(李晓光等,2017),而且其灌丛对周边环境的间接影响也会影响土壤CO2向大气库的输送。因此,本文以黄河三角洲柽柳灌丛周边土壤CO2通量为研究对象,分析其周边的温、湿度与土壤呼吸变化特征,解析柽柳灌丛对土壤CO2通量的影响,以期为黄河三角洲河口湿地碳循环与生态保育提供参考,对揭示滨海湿地自然植被对土壤碳库以及全球气候变化的影响具有一定的研究意义。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于东营市北部的黄河三角洲湿地,该区域受暖温带大陆性季风气候影响,四季分明、雨热同期、日照资源充足,年均气温12.1℃,年总降水量551.6 mm,蒸发量1928.2 mm,日照时数2759 h,年太阳辐射总量5278 J · m-2(丁晨曦等,2013)。土壤类型以隐域性潮土和盐土为主,土壤质地主要为砂质黏土(陈永金等,2018;靖淑慧等,2018),植被物种组成大多以盐生植物为主,群落结构简单(张绪良等,2009;刘加珍等,2015)。
1.2 研究方法
监测区选择无人为干扰且柽柳生长良好的群落(图1),选取群落中独立生长的柽柳灌丛,所选取的柽柳灌丛冠型相似且株高、冠幅相近。在柽柳灌丛周边布设监测点。本次考察重点选择南(S)、北(N)两个方向,分别布设6个监测点,每两个监测点间隔50 cm(图2)。于2015年4月、7月和10月选择晴朗的天气、采用开路式土壤碳通量测量系统(LI-8100A)进行数据监测。每个监测点提前24 h埋置基座,基座为直径20 cm、高11 cm的聚氯乙烯圆柱体,基座插入土壤6 cm,高露出地面5 cm,在每个基座内存留的地表植被自土壤表层彻底剪除,避免监测时植物的影响,减少对土壤的扰动。对监测点每2 h测定土壤碳通量,每次测定持续3 — 5 min,每个灌丛岛测定12次,共24 h。同时,在每个监测点同步测定温、湿度因子,测定指标包括近地表5 —10 cm的大气温度、大气湿度、土壤湿度(采用英国Delta-T土壤水分速测仪ML2X测定)与地面0 — 5 cm处的土壤温度。
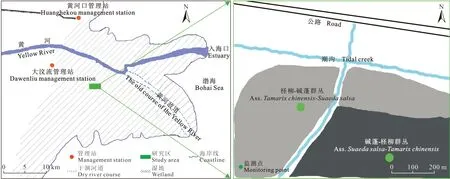
图1 黄河三角洲监测点布设Fig. 1 Sampling spot in the Yellow River Delta
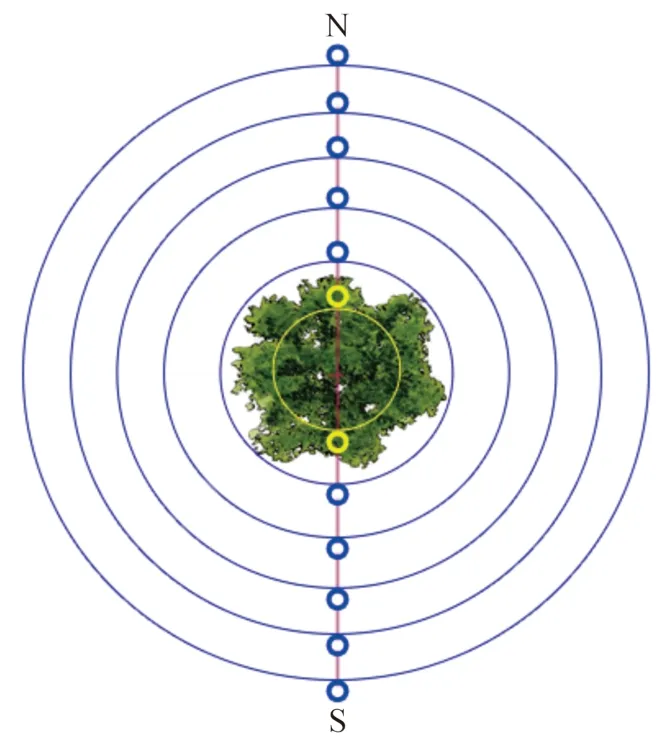
图2 柽柳周边监测点布设Fig. 2 The monitoring points around Tamarix chinensis
将分别在N、S方向随机获取的0 — 10 cm土样带回实验室,自然风干,将风干的土样挑出枯枝落叶后进行研磨,分析土壤全盐(残渣烘干法)和土壤有机质(重铬酸钾外加热法)(中国科学院南京土壤研究所,1978),实验测定在聊城大学环境生态实验室完成。成图与数据计算分别在Excel 2007与SPSS 17.0中完成。
2 结果与分析
2.1 柽柳灌丛的生境特点
监测区的植物群落类型,一种是以盐地碱蓬(Suaeda salsa(L.) Pall.)为优势种的碱蓬-柽柳群丛(Ass.Suaeda salsa-Tamarix chinensis),此群落中柽柳种群格局为随机分布型(徐梦辰,2016);另一种是以柽柳(Tamarix chinensisLour.)为优势种的柽柳-碱蓬群丛(Ass.Tamarix chinensis-Suaeda salsa),群落中柽柳种群分布格局为集聚分布(徐梦辰,2016)。从0 — 10 cm土层的土壤特征可知(表1):柽柳-碱蓬群丛中N与S方向的土壤有机质平均含量均大于碱蓬-柽柳群丛,而其土壤全盐的平均含量和变幅均小于碱蓬-柽柳群丛。在碱蓬-柽柳群丛中,全盐最大值高达5.92%,而柽柳-碱蓬群丛最大值为2.61%,且S方向的土壤全盐均值高于N方向。故柽柳-碱蓬群丛属于低盐-高养生境,而碱蓬-柽柳群丛属于高盐-低养生境。
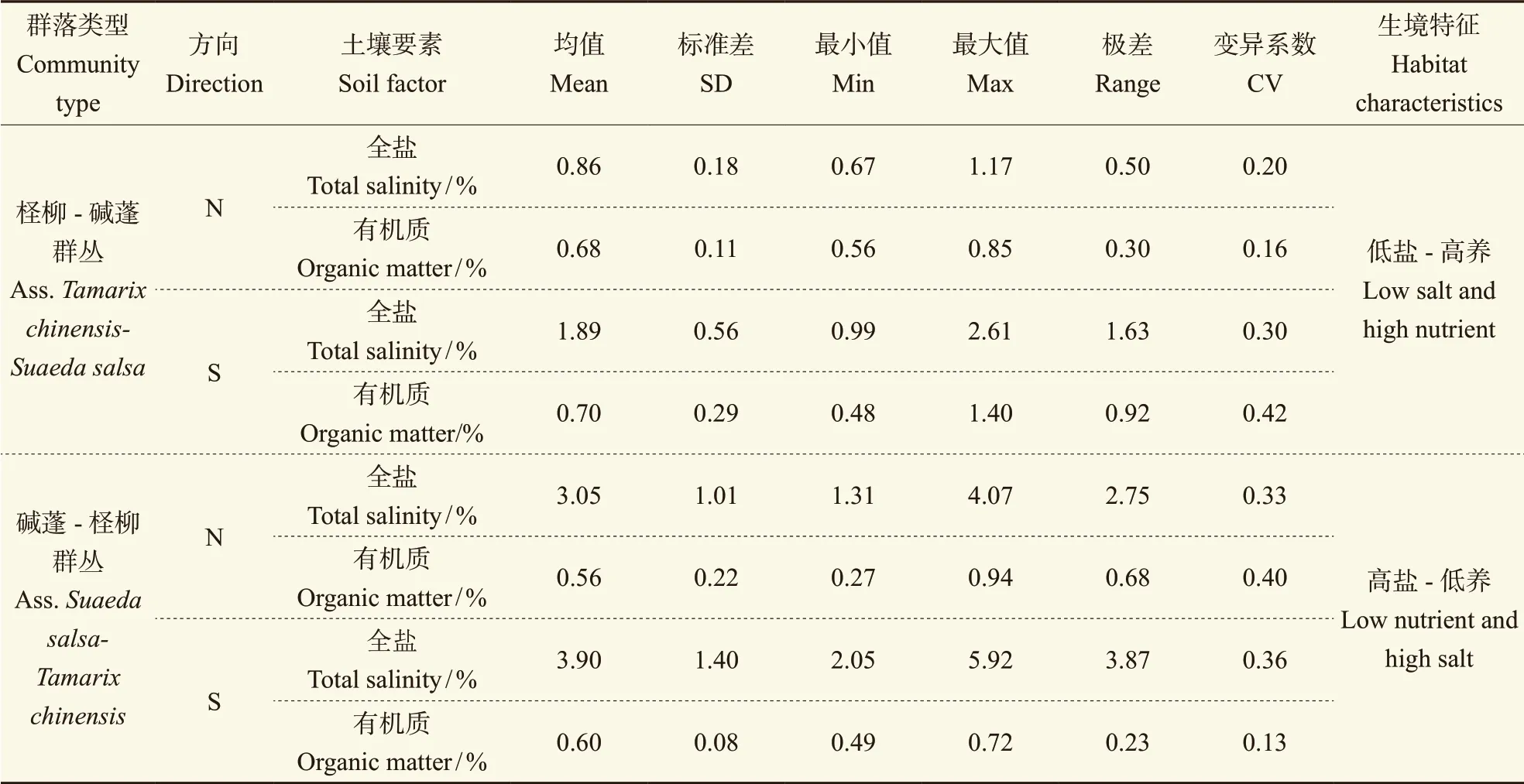
表1 柽柳灌丛0 — 10 cm土层的土壤有机质与全盐的描述统计Tab. 1 Description of soil organic matter and total salt in 0 — 10 cm soil layer around the Tamarix chinensis
2.2 柽柳灌丛周围的温湿度变化
由表2可知:春季,低盐生境与高盐生境柽柳周围的土壤温度、土壤含水率、近地表大气温度与大气湿度在南北方向上的变异系数相差不大,低盐生境土壤温度均值分别为23.99℃(N)、24.21℃(S)、近地表的大气温度分别为24.32℃(N)、24.45℃(S);高盐生境土壤温度均值分别为25.28℃(N)、25.07℃(S)、近地表的大气温度分别为25.95℃(N)、25.68℃(S)。但近地表的大气湿度极差值较大,北部的明显高于南部的,土壤含水率极差值同样也是北部高于南部。夏季,低盐生境的柽柳周围土壤均温分别为29.85℃(N)、29.42℃(S),高盐生境土壤均温分别为31.15℃(N)、31.11℃(S),南北方向上的近地表大气温度、大气湿度等均值差异不大,与春季不同的是土壤含水率极差值是南部高于北部,而高盐生境的柽柳南北方向的土壤含水率极差值、均值仍然是N方向高于S方向,且此环境下柽柳N方向的水分含量较高,变化幅度较大。秋季,在低盐生境中的柽柳周围除了土壤含水率之外,近地表的大气温湿度及土壤温度普遍高于高盐生境,这主要是由于秋季多风且高盐生境植被覆盖率低,尤其是木本灌丛少,柽柳周围的温湿度变化容易受外界干扰所致。高盐生境中土壤含水率的极差值与其他两个季节的不同,表现为柽柳S方向的变化要高于N方向的。
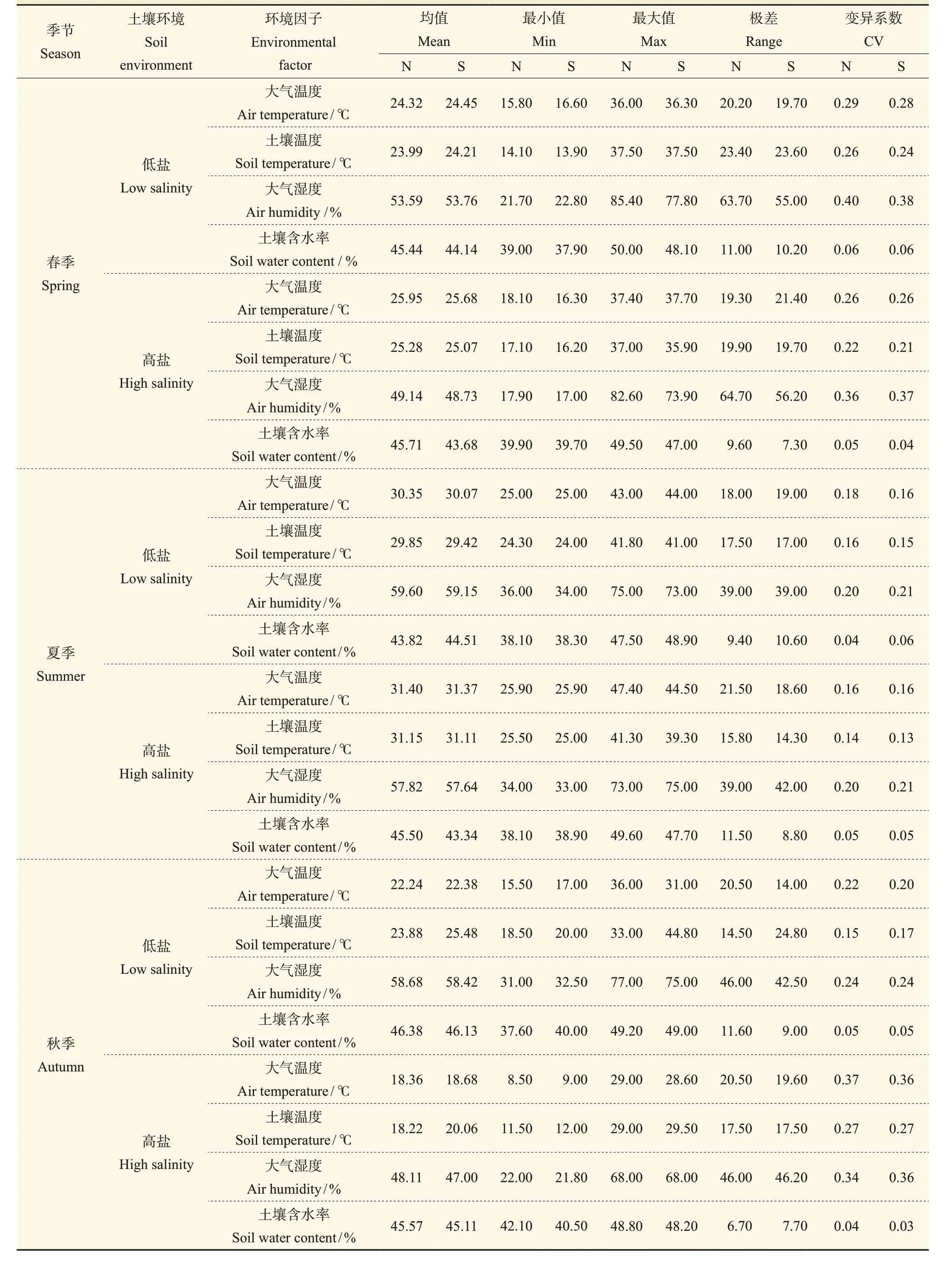
表2 两种土壤环境下柽柳周围温度与湿度的季节变化统计Tab. 2 Seasonal variation of temperature and humidity around the Tamarix chinensis in two soil environments
为了进一步对比柽柳南北方向温湿度的空间变化,对不同量纲的日均值进行总和标准化处理。以标准化值为Y轴,以北向南到柽柳的不同距离为X轴,得到高盐生境(图3)与低盐生境(图4)柽柳南北方向不同距离的温湿度变化图。春季,高盐生境(图3)近地表大气温度在距离柽柳近、远处的变化波动不大,但近地表的大气相对湿度在距离柽柳南北方向150 cm的范围内相对较高,远处的明显较低,呈单峰曲线变化。土壤温度从柽柳灌丛中心的0 — 300 cm向南北方向呈现波动的“V”字型,而土壤含水率由北向南呈逐渐减少的趋势,在柽柳S方向100 — 300 cm土壤含水率的日均值变化不大。
夏季,近地表大气温度与湿度呈相反的变化趋势,土壤温度与土壤含水率也呈现相反的变化趋势,即近柽柳处为低温高湿度的状态,远离柽柳处为高温低湿的状态。秋季,土壤温度明显呈现S方向高于N方向的分布特征;近地面大气与相对湿度在N方向为低温高湿状态,S方向为高温低湿状态。土壤含水率与春季相似,即由北向南呈逐渐减少的趋势,但变化幅度比春季要小。
春季,低盐生境(图4)柽柳周边空间波动最大的是土壤温度与土壤含水率,土壤温度呈“U”型、土壤含水率呈“V”型,即南北距离柽柳100 cm的范围内为低温低湿状态,100 cm之外则相反;夏季,柽柳周边南北方向温湿度的空间变化与春季相似。秋季,与高盐生境相同的是土壤温度明显出现S方向高于N方向的分布特征,这与秋季多北风有关。
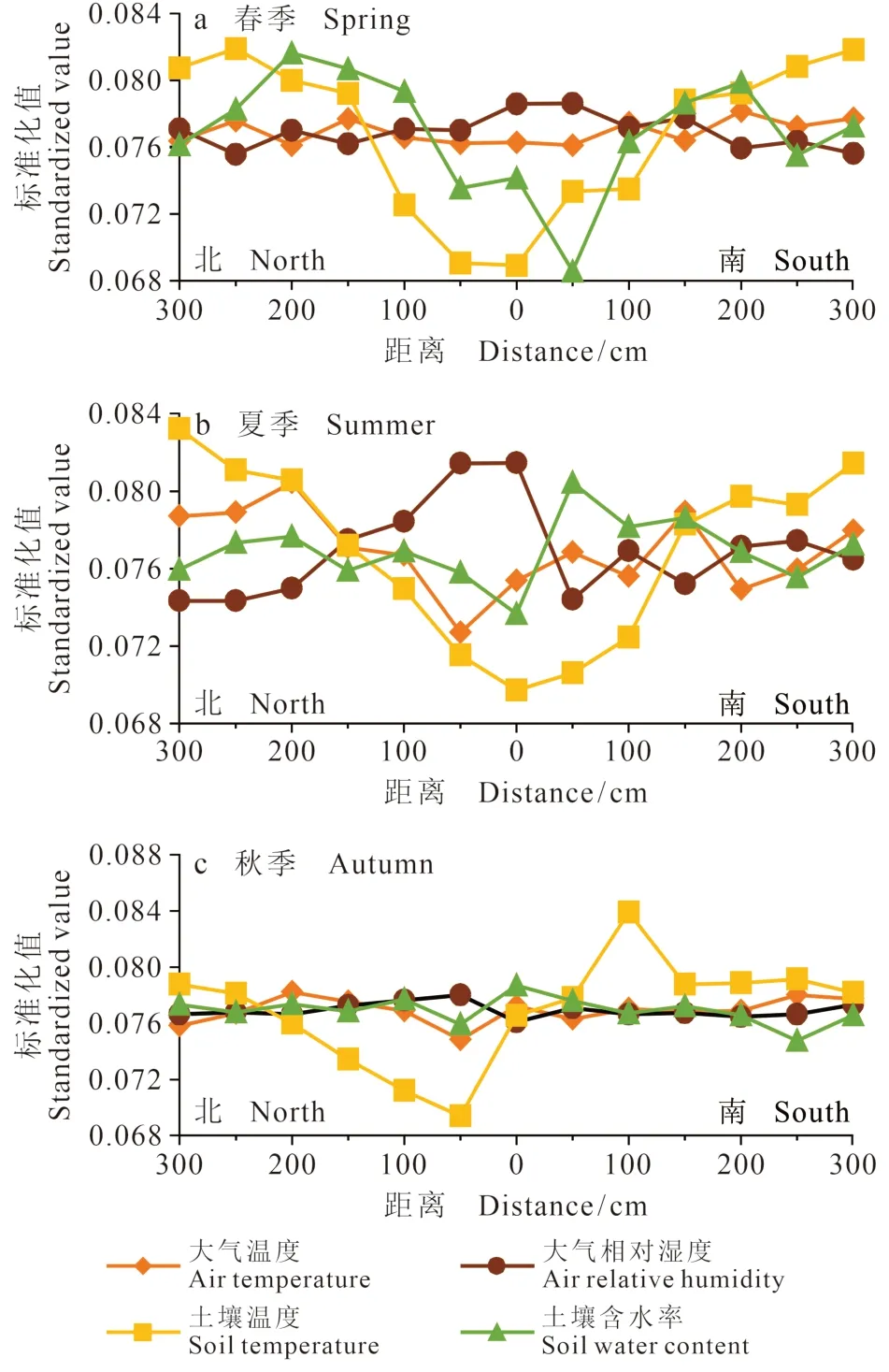
图4 低盐生境柽柳南北方向不同距离的温湿度变化Fig. 4 Temperature and humidity changes from north to south direction around Tamarix chinensis in low salt habitat
2.3 不同生境中柽柳周边的土壤呼吸季节变化
2.3.1 春季的土壤呼吸差异
由图5可知:两个群落中柽柳灌丛岛的土壤呼吸速率大致表现为距柽柳中心越近,土壤呼吸速率越大,距柽柳50 cm与300 cm处的土壤呼吸速 率 的 最 大 差 值 为1.11 μmol · m-2·s-1;土 壤 呼 吸速率的日变化基本呈先增大后减小的变化趋势,最高值出现在10∶00 — 16∶00。两种生境中土壤呼吸速率的变幅是不同的,在高盐低养生境中(图5a),柽柳灌丛岛,土壤呼吸速率的变幅为0.12 — 1.30 μmol · m-2·s-1,最 高 值 出 现 在 中 午12∶00;在低盐高养生境中(图5b),柽柳灌丛岛土壤呼吸速率变幅为0.01 — 1.62 μmol · m-2·s-1,最高值有明显的滞后性,最高值出现在16∶00左右。两种生境的柽柳灌丛下土壤呼吸速率的最大值相差0.32 μmol · m-2·s-1,反映出养分盐分对于土壤呼吸的影响。
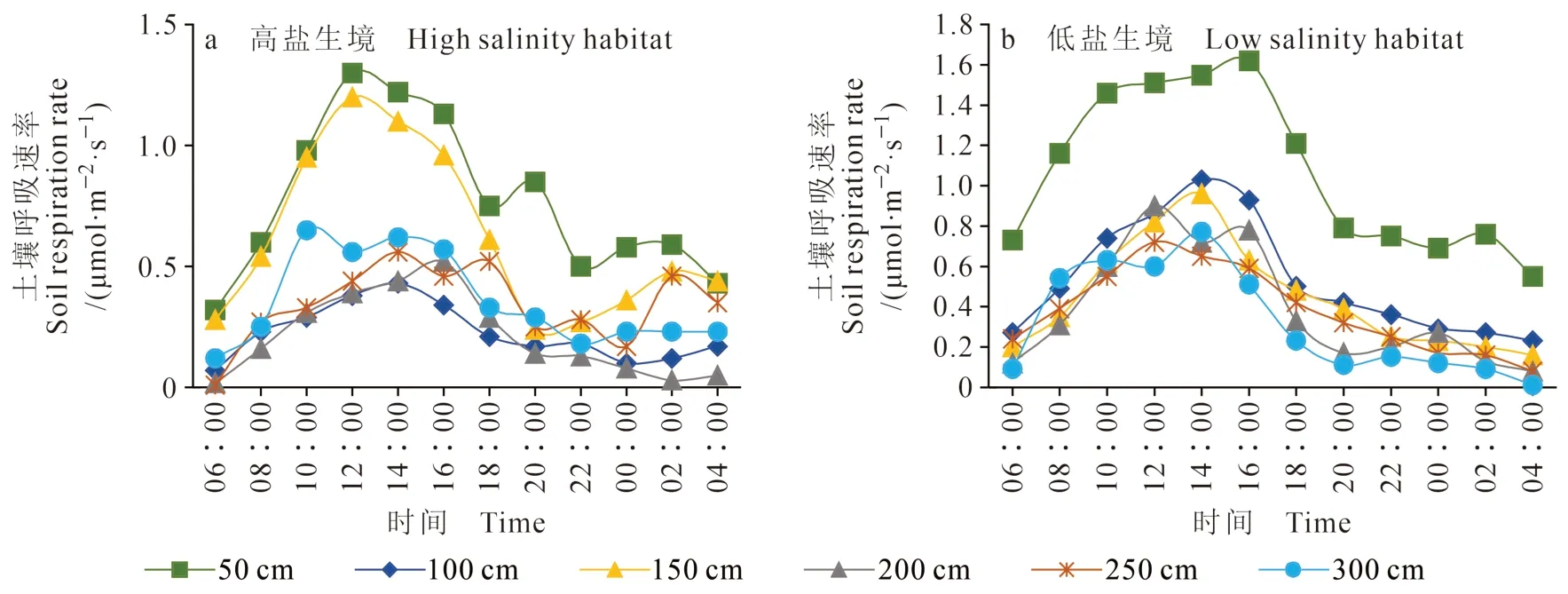
图5 春季不同生境条件下柽柳周围的土壤呼吸特征Fig. 5 Characteristics of soil respiration in two habitats in spring
2.3.2 夏季的土壤呼吸差异
由图6可知:与春季相比夏季的土壤呼吸速率整体要高,变化趋势与春季相同,灌丛周边土壤呼吸速率表现为由灌丛中心向外围逐渐减小的趋势,柽柳冠幅覆盖下的土壤呼吸速率与灌丛周边裸地的最大差值为3.01 μmol · m-2·s-1。同时,受柽柳灌丛的影响,距主径50 cm处的土壤呼吸速率明显高于其他距离处,并且此距离处的土壤呼吸速率没有非常显著的峰值(图6b)。夏季土壤呼吸速率的日变化基本呈先增大后减小的变化趋势,最高值出现在12∶00 — 16∶00。其中,高盐低养生境下(图6a)柽柳灌丛周边的土壤呼吸速率的变幅为0.38 —2.06 μmol · m-2·s-1,距柽柳50 cm处土壤呼吸速率的最高值出现在12∶00左右;低盐高养生境下(图6b)柽柳灌丛下的土壤呼吸速 率 变 幅 为0.31 — 3.42 μmol · m-2·s-1,日 变 化 最大值具有明显的滞后性,距柽柳50 cm处出现在16∶00左右。两种生境下的土壤呼吸速率最大值相差1.36 μmol · m-2·s-1,低盐高养生境中的土壤呼吸速率普遍较高。
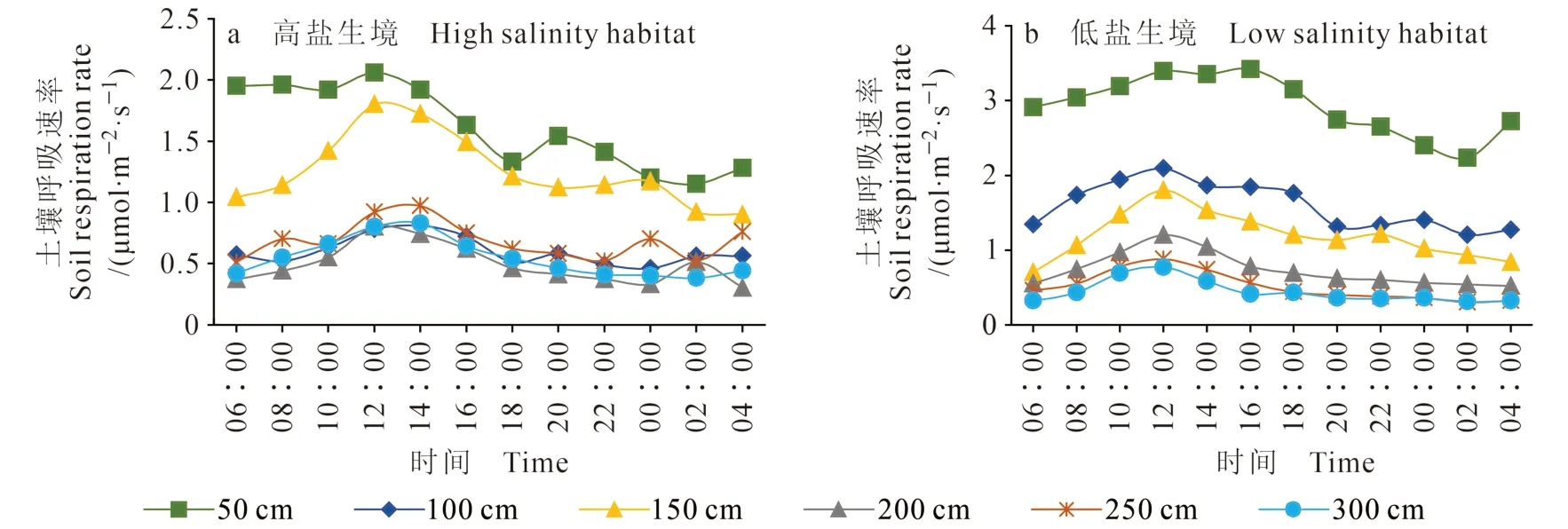
图6 不同生境夏季土壤呼吸特征Fig. 6 Characteristics of soil respiration in two habitats in summer
2.3.3 秋季的土壤呼吸差异
由图7可知:秋季的土壤呼吸速率值介于夏季、春季之间,变化趋势与两个季节基本一致,但其日变化峰值较为凸出,尤其是在高盐生境中。高盐低养生境中(图7a)土壤呼吸速率 的 变 幅 在 0.12 —1.75 μmol · m-2·s-1,低 盐 高养生境(图7b)的土壤呼吸速率变幅在0.08 —2.19 μmol · m-2·s-1,变化幅度明显低于夏季。其中,距柽柳主径50 cm处的土壤呼吸速率仍高于其他距离处,生长于高盐低养生境的柽柳灌丛周边的土壤呼吸速率都具有显著的峰值(图7a),而低盐高养生境灌丛下的土壤呼吸速率值则相反,说明该生境下柽柳灌丛对土壤呼吸的影响较为明显。
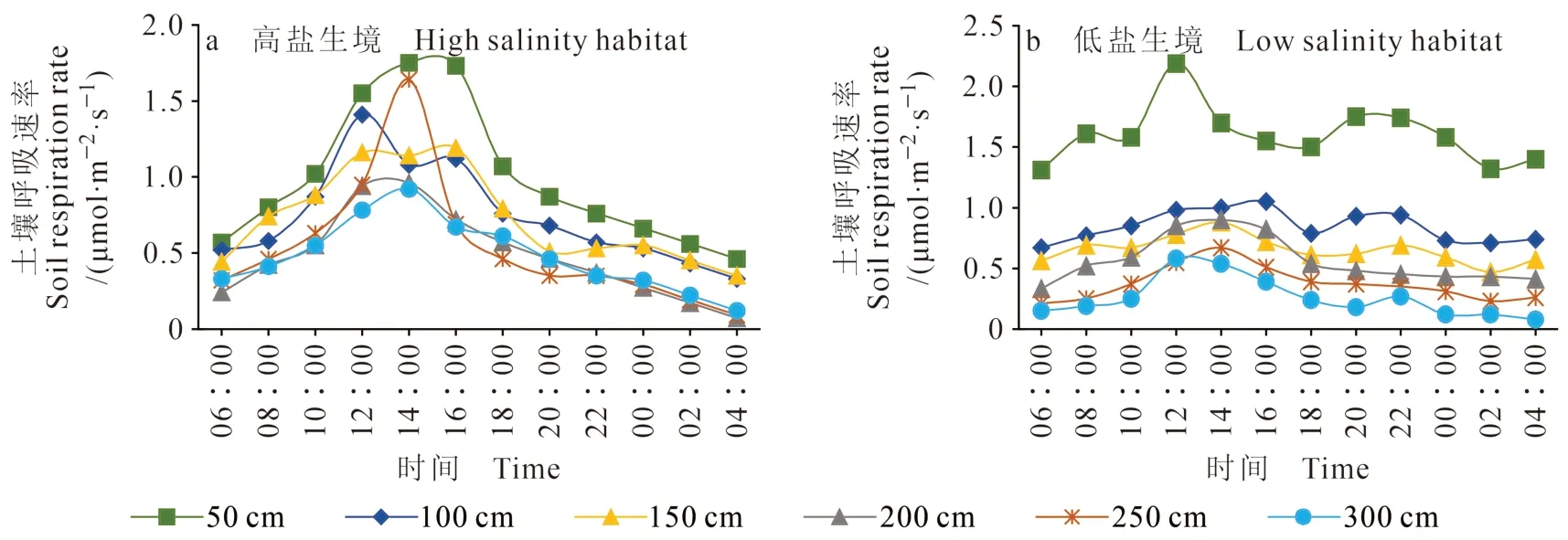
图7 不同生境秋季土壤呼吸特征Fig. 7 Characteristics of soil respiration in two habitats in autumn
土壤呼吸速率的日动态变化多呈单峰型曲线(王丹等,2016),且最高值出现在中午的12∶00 — 14∶00(王丰川等,2013)。在地面无积水时,该湿地土壤呼吸最大值出现在12∶00左右(朱敏等,2013)。本研究两种生境中的灌木土壤呼吸日变化也呈现明显的单峰曲线,与其他人研究结果相一致,但本研究中峰值出现在10∶00 — 16∶00,尤其在低盐高养环境中,距柽柳50 cm处的土壤呼吸速率没有明显的峰值,说明柽柳灌丛影响下土壤呼吸速率的变化不敏感。
2.4 土壤呼吸的昼夜差异与空间变化
2.4.1 土壤呼吸昼夜差异
土壤呼吸有明显的昼夜变化,通常白天的土壤呼吸速率要高于夜间(冯文婷等,2008;戴雅婷等,2018)。以研究区日出与日落前后光照程度为昼夜区分的依据(春秋季白昼05∶00 — 18∶00;夏季白昼04∶00 — 20∶00),将春、夏、秋三季南北方向土壤呼吸的平均值(图8)比较可知:白天土壤呼吸速率明显高于夜晚,最大差值可达0.45 μmol · m-2·s-1;南北方向同样存在明显季节差异,夏季N方向的土壤呼吸速率高于S方向,而春秋两季则恰恰相反,说明柽柳灌丛对南北方向上的温度、湿度及风速等环境条件的影响明显,从而导致土壤呼吸速率在南北方向存在明显的季节变化。在低盐生境中(图8b),柽柳灌丛周边土壤呼吸速率的昼、夜最大值均出现在夏季的N方向,分别为1.38 μmol · m-2·s-1和1.02 μmol · m-2·s-1;在高盐生境(图8a)中分别为0.98 μmol · m-2·s-1和0.72 μmol · m-2·s-1。由表3可知:两种不同生境灌丛周边白天的土壤呼吸速率均值、标准差和变幅均大于夜晚,变异系数则小于夜晚,说明夜晚不同距离上的土壤呼吸速率离散程度较大,即不同距离上的土壤呼吸速率差异较大。总之,高盐生境中柽柳周边土壤呼吸昼夜差异最小的季节是夏季,低盐生境灌丛周边昼夜差异最小的是秋季。高盐与低盐生境中,柽柳周边土壤呼吸速率昼夜差异较大的均在春季。春、夏、秋三个季节白天的土壤呼吸速率总和约占昼夜总量的60%,夜间土壤呼吸速率约占40%。由此可见,土壤CO2夜间释放量不容忽视。
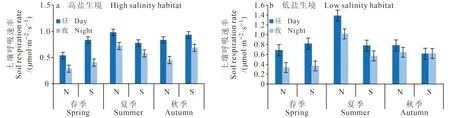
图8 不同生境下柽柳周围土壤呼吸速率昼夜差异Fig. 8 Difference of soil respiration rate between day and night around the Tamarix chinensis in different habitats
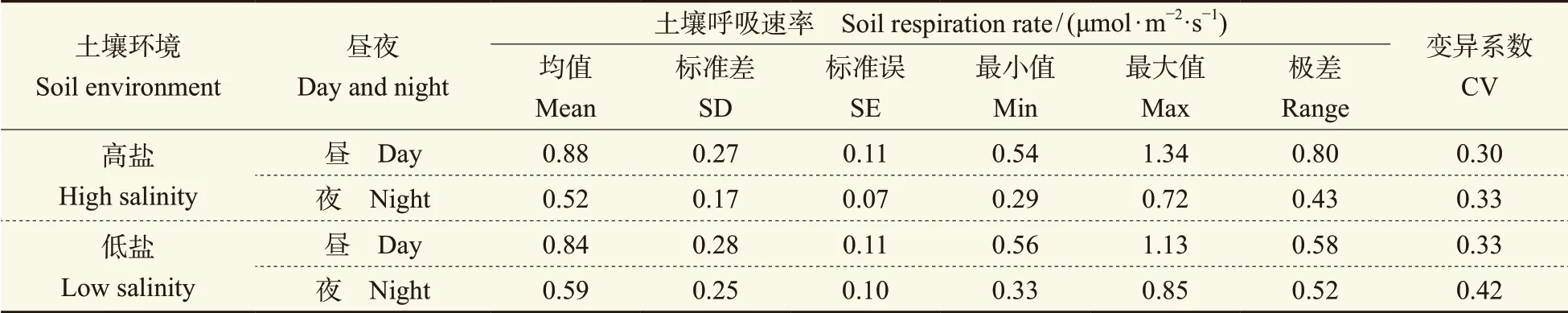
表3 两种生境下柽柳周围土壤呼吸昼夜差异Tab. 3 Difference of soil respiration rate between day and night around the Tamarix chinensis in different habitats
2.4.2 土壤呼吸的空间差异
环境的异质性变化往往会导致土壤呼吸的空间异质性。从距离柽柳灌丛50 cm、100 cm、150 cm、200 cm、250 cm、300 cm监测点的土壤呼吸速率来看(图9、图10),离柽柳较近处的土壤呼吸速率相对较高,随着距离的增加土壤呼吸速率逐渐降低。这种规律在低盐生境中尤为明显(图10),春、夏、秋三个季节都表现出距离递减规律,这是由于冠下枯落物腐烂致使其土壤养分状况好且土壤微生物活性高,促进根系活动
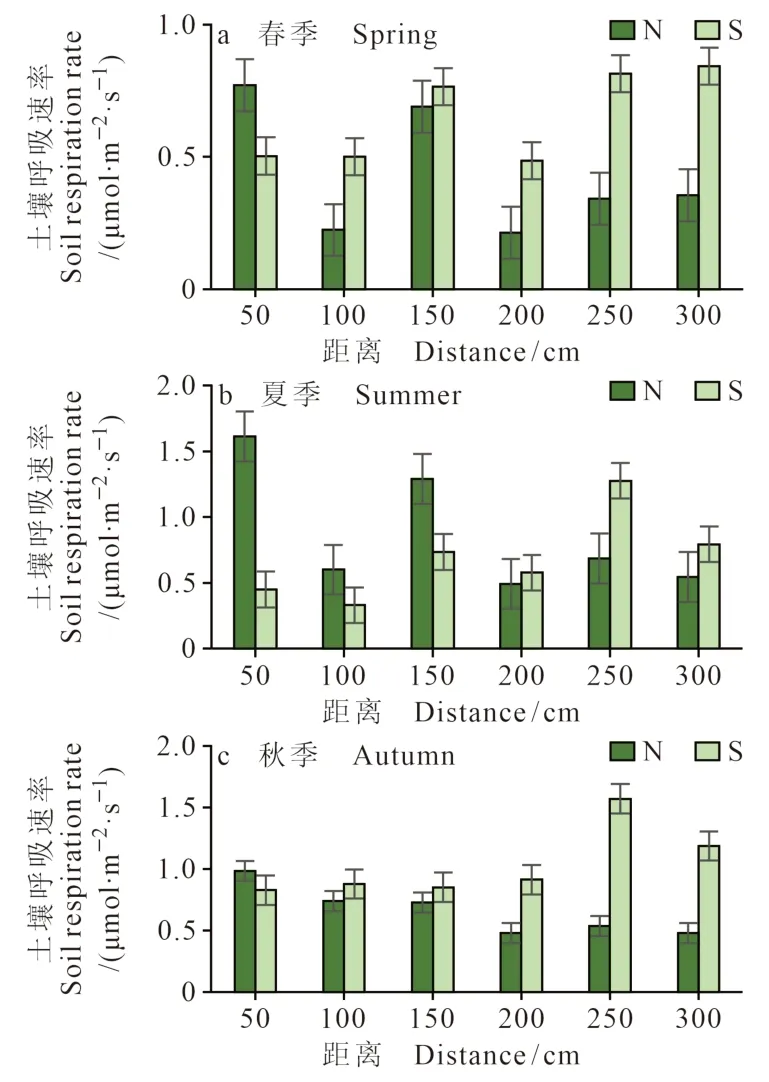
图9 高盐生境中南北方向上的土壤呼吸差异Fig. 9 Difference of soil respiration in the north — south direction in high salt habitat
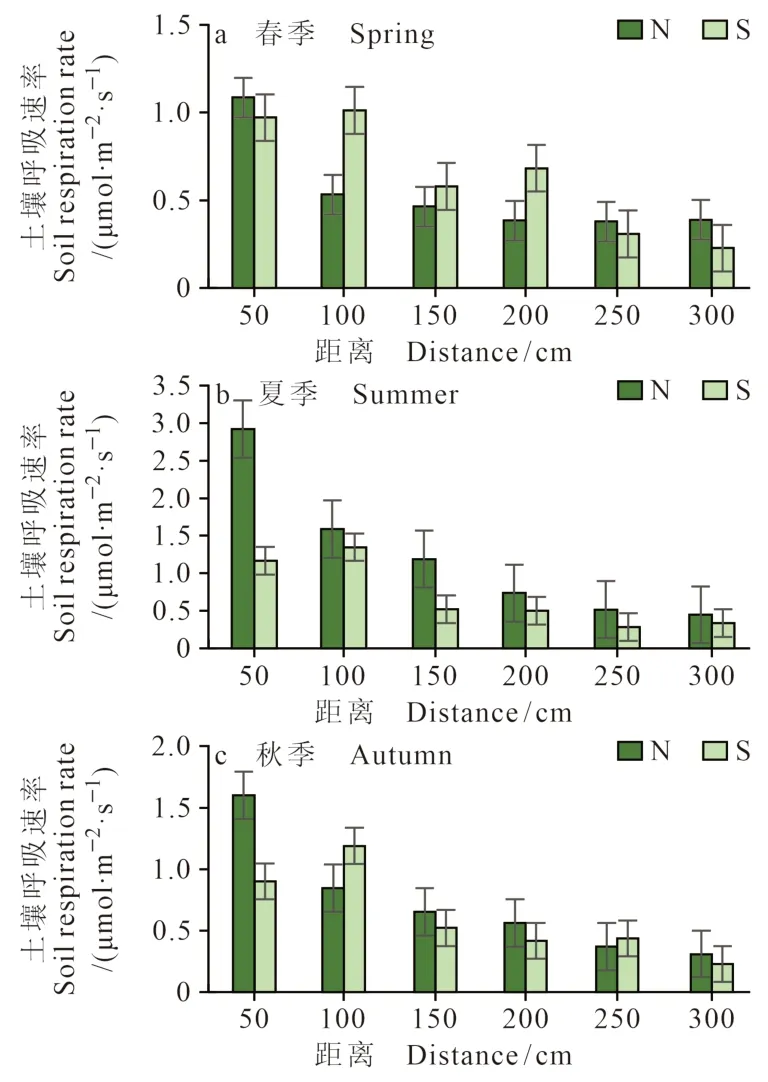
图10 低盐生境中南北方向上的土壤呼吸差异Fig. 10 Difference of soil respiration in the north — south direction in low salt habitat
在高盐生境中的灌丛周边土壤的呼吸速率,春、夏、秋三季,N方向基本表现为由灌丛岛中心向外呈逐渐递减的变化趋势,但S方向则呈现出相反的变化趋势,表现为近“岛屿”处的土壤呼吸速率低于远“岛屿”处。这种特征的影响因素存在两方面的可能:一方面是温、湿度的较大波动,有研究表明干湿波动可引起土壤呼吸的激发效应,且这类效应在各类生态系统中普遍存在(王融融等,2022)。在春秋两季S方向远离柽柳的地方,其土壤温、湿度的变异系数(图11)波动都较大,尤其是土壤温度由近及远变异系数呈增加趋势。夏季,在远离柽柳300 cm处的土壤温湿度的变异系数波动较小,而250 cm处的土壤温湿度波动较大,故夏季S方向250 cm处的土壤呼吸值高于300 cm处。另一方面,与风的影响有一定关系,黄河三角洲受海陆风的影响,风能资源丰富、风速较大,盛行风随季节的变化而变化(刘敦训等,2005),风可以将植物的枯落物等吹到低洼地聚集,从而促进微生物的活动等,从而使得土壤呼吸速率较大。和土壤呼吸作用,使近“岛”处土壤呼吸速率要明显高于远“岛”处。
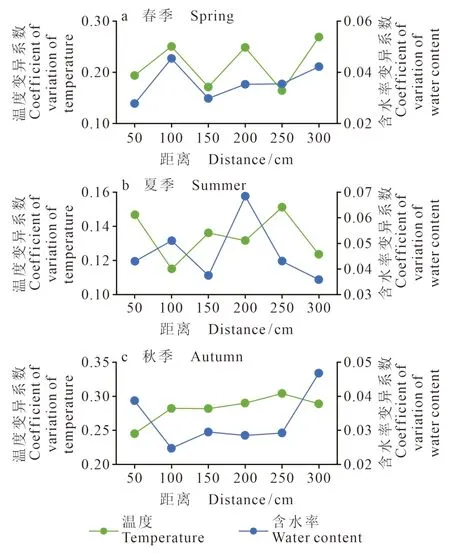
图11 高盐生境S方向上的土壤温湿度变化特征Fig. 11 Variation characteristics of soil temperature and humidity in S direction of high salinity habitat
统计分析(表4)可知:无论高盐生境还是低盐生境,春季柽柳萌发生长期,S方向的均值大于N方向,而标准差、变幅和变异系数小于N方向,夏季柽柳生长旺盛期,S方向的均值、标准差、变幅和变异系数均小于N方向,等秋季柽柳进入落叶期,S方向的均值、标准差和变幅大于N方向,变异系数则小于N方向。其中,低盐高养生境柽柳周边夏秋两季土壤呼吸的均值、标准差、变幅和变异系数均是N方向高于S方向,春季则相反;高盐低养生境影响下,柽柳周边土壤呼吸速率春夏两季S方向的均值、标准差、变幅和变异系数均小于N方向,秋季则相反。
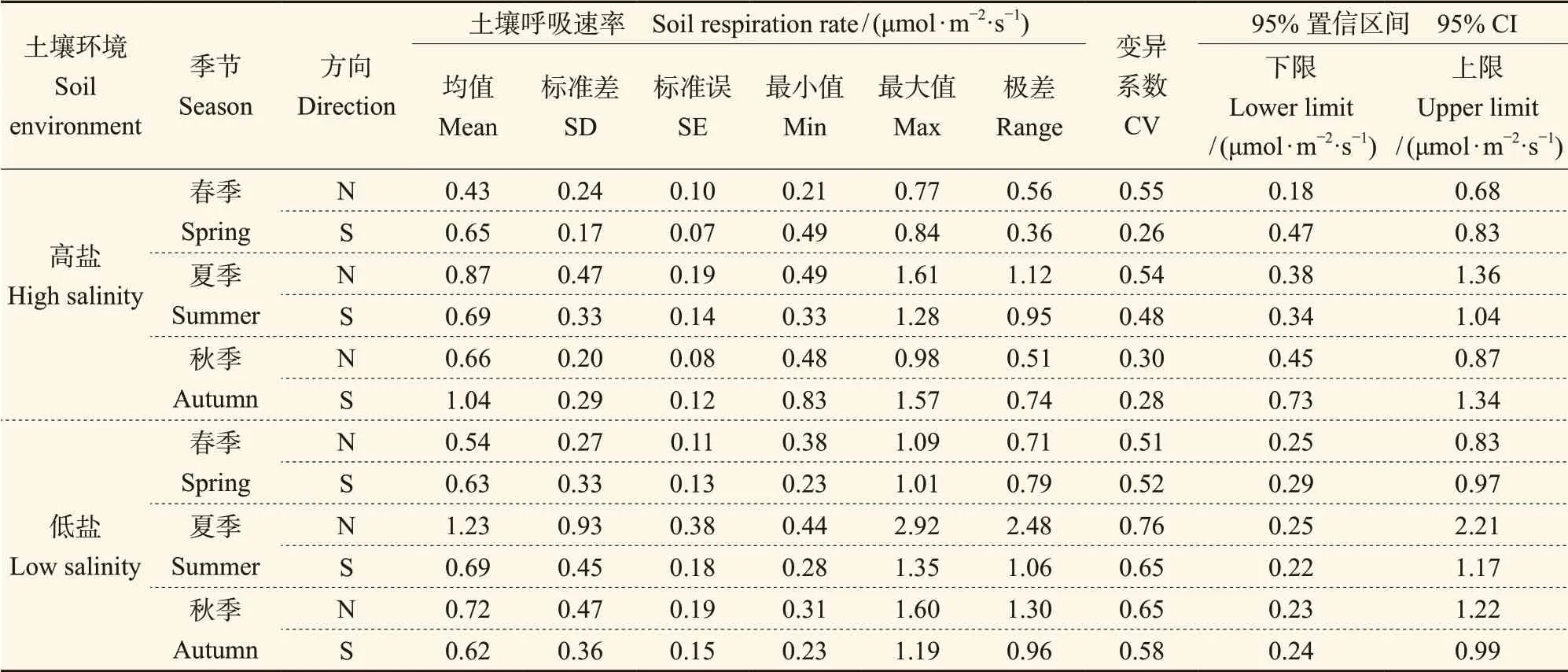
表4 不同季节柽柳南北方向上土壤呼吸的差异Tab. 4 Difference of soil respiration in the north — south direction of Tamarix chinensis in different seasons
3 结论
(1)研究区柽柳灌木生长的生境可分为高盐低养与低盐高养两种类型,不同生境中的柽柳其周边土壤呼吸的变化特征不同。低盐生境中的柽柳灌丛,其周边的土壤呼吸速率明显高于高盐生境。春、夏、秋三个季节,高盐生境下柽柳灌丛周边土壤呼吸速率的日动态呈明显的“单峰曲线”特征,峰值出现在10∶00 — 16∶00。但低盐生境中,夏秋两季柽柳灌丛影响下的土壤呼吸速率日动态的“单峰曲线”变化不明显。
(2)春、夏、秋三个季节,柽柳灌丛周边的土壤呼吸速率差异明显。柽柳灌丛周边南北不同方向上的温度、湿度差异,使得土壤呼吸速率夏季N方向的土壤呼吸速率高于S方向,而春秋两季则相反。无论低盐生境还是高盐生境,灌丛周边土壤呼吸速率昼、夜差异以春季最为明显,夜间土壤呼吸速率不容忽视,约占昼夜总和的40%。
(3)环境的异质性变化会导致土壤呼吸的空间异质性。距离柽柳灌丛50 cm、100 cm、150 cm、200 cm、250 cm、300 cm监测点的土壤呼吸速率空间变化规律为:随着距离的增加土壤呼吸速率逐渐降低,尤其是低盐生境中柽柳灌丛周边。但高盐生境S方向表现为近“岛屿”处的土壤呼吸速率低于远“岛屿”处,其标准差、变幅和变异系数均小于N方向,这与土壤温度、湿度的日较大变化有一定关系,还可能受风的影响。
猜你喜欢
农业工程学报(2022年13期)2022-10-09
中南林业科技大学学报(2022年7期)2022-09-26
中国农业气象(2022年8期)2022-08-24
农业与技术(2022年15期)2022-08-15
长江科学院院报(2022年7期)2022-08-09
东北林业大学学报(2022年1期)2022-03-28
种子(2022年2期)2022-03-21
土壤(2022年1期)2022-03-16
风景园林(2022年1期)2022-03-07
人民长江(2019年7期)2019-09-10
