
外源γ-氨基丁酸对鲜切南瓜品质和γ-氨基丁酸代谢的影响
2022-09-27 15:09梁静宜王鸿飞
食品工业科技 2022年19期
梁静宜,郭 凡,赵 科,王鸿飞,许 凤
(宁波大学食品与药学学院,浙江宁波 315800)
-氨基丁酸作为一种自由态的四碳非蛋白氨基酸,广泛分布在自然界的原核生物和真核生物中,是哺乳动物中枢系统抑制性神经递质,参与脑循环,具有降低神经元细胞活性、防止动脉硬化、改善肝功能和降低血压等作用。在植物体内合成GABA 有两条代谢途径,主要由谷氨酸在GAD 的催化下生成GABA,由GABA-T 催化GABA 进入三羧酸循环中,这条代谢途径被称为GABA 支路。GABA 在多胺降解途径中通过二胺氧化酶(Diamine oxidase,DAO)和多胺氧化酶(Polyamine oxidase,PAO)两种关键酶催化多胺(腐胺(Putrescine,Put)、精胺(Spermine,Spm)、亚精胺(Spermidine,Spd))后,再由4-氨基丁醛脱氢酶(Aminoaldehyde decarboxylase,AMADH)将其氧化生成GABA 留在植物体内。
研究发现,GABA 参与植物C/N 营养平衡、植物微生物相互作用、抵御虫害、响应生物和非生物胁迫以及作为细胞内信号分子调控传导等活动。当生物体在处于钠盐、低氧、机械损伤等胁迫条件下时,会产生应激反应,富集大量GABA 来提高自身的抗逆性,从而更好地适应环境。机械损伤下猕猴桃和胡萝卜体内会迅速合成大量的GABA,响应非生物胁迫。近年大量关于GABA 外源应用的研究指出,GABA 用于水果或蔬菜的采后处理,可以有效缓解生物或非生物胁迫的影响。
南瓜富含多种氨基酸和维生素,具有预防糖尿病、溶解结石以及催化分解致癌物质等作用,且价格低廉,已然成为食品界的新宠。近年来,由于快餐和方便食品行业的迅速发展,鲜切产品吸引了大量消费者的关注。鲜切南瓜因其营养高,味道好和便利性更是受到消费者的广泛青睐。然而,南瓜受到由加工处理(如去皮、修剪和切割)导致的组织分解引起的损伤,难免会发生表皮微生物侵染、营养损失、质地软化和颜色变化等情况,降低鲜切产品的潜在经济价值。因此,需要研究一种方法来抑制鲜切南瓜品质变化,同时保证产品健康安全。以往对GABA 的研究主要集中在代谢层面上,然而,GABA 对鲜切南瓜品质的影响与否尚不清楚。本文通过外源GABA 和GABA 合成抑制剂(3-巯基丙酸,3-MP)处理研究其对鲜切南瓜中GABA 代谢和品质的影响。主要测定不同处理下鲜切南瓜中谷氨酸、GABA 和多胺含量,GAD、GABA-T、PAO、DAO、AMADH 活性以及相关基因表达水平和品质指标,以解决鲜切南瓜贮藏期间品质变化的相关问题,为鲜切南瓜GABA 富集工艺转化提供重要的理论和实践依据。
1 材料与方法
1.1 材料与仪器
本实验选用新鲜南瓜(贝贝南瓜)为供试材料,购买于京东商场,挑选大小均匀,表面无病虫害和机械损伤;GABA 标准品(98%)、甲醇(色谱纯)Sigma-Aldrich 公司;3-巯基丙酸(3-Mercaptopropionic acid,3-MP)、谷氨酸脱氢酶(1 U/mL)分析级,麦克林;氯化镧、-烟酰胺腺嘌呤二核苷酸(-NAD+)分析级,阿拉丁;Tris-HCl 溶液 分析级,上海生工;腐胺、精胺、亚精胺标准品 分析纯(T≥98%),上海源叶生物科技有限公司;植物总RNA 提取试剂盒 成都福际生物技术有限公司;RNA isolater Total RNA Extraction Reagent HiScript® Ⅱ QRT SuperMix for qPCR(+Gdna Wiper)试剂盒、ChamQTMSYBR®qPCR Master Mix 试剂盒 南京诺唯赞生物科技有限公司;其他试剂均为国药分析纯。
AFX-2001-U 超纯水机 上海纯浦实业有限公司;SYNAPT G2 超高效液相色谱仪 美国Water 公司;UV-3200 型紫外分光光度计 上海普美达;Mastercycler nexus PCR 仪 Eppendorf 中国有限公司;A40426 实时荧光PCR 仪 赛默飞世尔科技(中国)有限公司;CR-400 型色差仪 柯尼卡美能达办公系统(中国)有限公司;PAL-1 型数显糖度计 日本爱宕ATAGO 公司;JXDC-200 型氮吹仪 拓赫机电科技(上海)有限公司。
1.2 实验方法
1.2.1 原料处理 南瓜在0.02%(v/v)的次氯酸钠溶液中浸泡2 min 进行消毒,将其削皮去瓤后,切成长约5 cm,宽0.2 cm,高0.5 cm 左右的丝状,并分别浸泡在80 mmol/L GABA 和80 mmol/L 3-MP 溶液中作为处理组(经过预实验得到最佳处理浓度),以浸泡在蒸馏水中的南瓜丝作为对照组。浸泡10 min 后取出用滤纸擦去表面多余的液体并放置在塑料透明鲜切盒内,并置于4 ℃,相对湿度90%培养箱中贮藏,分别于0、3、6、9 h 取样,用液氮快速冷冻,并置于-80 ℃冰箱中备用。在本实验中,每个处理至少使用12 个南瓜。
1.2.2 品质指标测定 菌落总数测定参考食品安全国家标准食品微生物学检验菌落总数测定(GB 4789.2-2016),白度使用色差仪测定,随机选取若干点测定样品的颜色参数,得到白色系中的L值,白度值(White index,WI)的计算公式为:WI=100-[(100-)++];可溶性固形物含量的测定采用郭丹等的方法,取10 g 的样品与20 mL 的蒸馏水使用匀浆机混匀,取200 μL 的溶液放置在糖度计上测定,记录数值;-胡萝卜素含量的测定参考食品安全国家标准食品中胡萝卜素的测定(GB/T 5009.83-2003)。
1.2.3 谷氨酸与GABA 含量的测定 谷氨酸含量的测定是在Al-Quraan 等的方法上稍作修改,将冷冻样品用冰浴在3 mL 氯化镧(0.05 mol/L)中进行提取。在0.5 mL 提取物中加入0.2 mL 的0.1 mmol/L Tris-HCl,0.1 mL 的7.5 mmol/L-NAD和1 U/mL谷氨酸脱氢酶后充分震荡混合,然后将混合液30 ℃水浴孵育60 min,读取波长为340 nm 时的吸光度。结果用mmol·kg来表示。计算公式如下:

GABA 含量的测定参照Hu 等的方法,在氯化镧溶液中加入0.5 g 样品进行萃取,在室温下震荡15 min后12000×g 离心5 min。取2 mL 上清液加入1 mol/L氢氧化钾溶液,振荡5 min 后12000×g 离心5 min。取0.4 mL 上清液依次加入800 μL 1 mmol/L 硼酸盐缓冲液(pH9.0)、1 mL 体积分数为6%的苯酚和0.5 mL 5%的次氯酸钠,混合均匀后沸水浴10 min,拿出后立即冰浴5 min。加入1 mL 体积分数为60%的乙醇,混匀后采用紫外分光光度法,在645 nm 处测定吸光度。结果用mmol·10 gFW 来表示。计算公式如下:

1.2.4 谷氨酸脱羧酶与GABA 转氨酶活性的测定GAD 活性的测定参考Bartyzel 等的方法并稍加修改,称取1.0 g 南瓜样品,在冰浴条件下通过含有-巯基乙醇、PLP、EDTA 和甘油的磷酸钾缓冲液研磨获得匀浆。最大转速离心10 min 后,取0.5 mL 粗酶液加入含有10 g/L 谷氨酸缓冲液,40 ℃下水浴2 h后通过沸水浴5 min 终止反应。用等量蒸馏水对照,在645 nm 下比色,测吸光度。每分钟生成1 μmol的GABA 为GAD 活性为一个酶活力单位。
GABA-T 活性的测定参照Deewatthanawong等的方法并加以修改,取0.5 g 南瓜果肉,加入Tris-HCl 提取缓冲液研磨,离心获得上清液即为酶液。取0.5 mL 酶液与缓冲液混合均匀后,放置30 ℃水浴中孵育1 h,加入磺基水杨酸后,再在25 ℃下孵育20 min 终止反应。在340 nm 处读取吸光值,每30 s 记录1 次,记录5 min。以1 min 内OD降低0.01 为一个酶活力单位(U),活性表示为U·g。
1.2.5 多胺的测定 参照宋春波等所述的方法,将南瓜样品在冰浴中加入3 mL 5%预冷的高氯酸进行均质研磨,12000×g 离心收集上清液。取2 mL 的上清液与2 mL 的NaOH(2 mol/L)和10 μL 的苯甲酰氯混合在37 ℃下孵育1 h,然后加入饱和NaCl溶液和乙醚,充分振荡混合均匀。通过2000×g 低速离心后,将1 mL 有机溶剂相转移至新管中,并用氮吹仪吹干。待乙醚蒸发后,加入500 μL 甲醇,充分震荡得到待测定的多胺样品。通过配备反相Sun Fire C色谱柱(150 mm×3.9 mm,4 μm)的高效液相色谱(HPLC)分析苯甲酰化后的多胺,色谱条件:柱温30 ℃;流动相为甲醇和水(比例为64:36,v/v);检测波长为230 nm;流速0.7 mL/min,进样量10 μL。
将Put、Spd 和Spm 3 种多胺分别配成1 mmol/L的标准贮备液,然后将这3 种标准贮备液分别苯甲酰化。将衍生化的母液稀释形成不同浓度的多胺标准液(1、5、12.5、25、50 mmol/L)进样,根据标品进样量制作标准曲线,然后以峰面积计算多胺的含量。以鲜样中的mg/kg 来表示。
1.2.6 胺氧化酶活性测定 参照Gao 等的方法测定DAO 和PAO 活性。取1 g 南瓜样品与磷酸钾缓冲液(pH6.5)冰浴研磨后,将样品在4 ℃下12000×g离心,收集上清液。取0.5 mL 上清液加入磷酸钠缓冲液(含有0.2 mL 显色液,250 U/mL 辣根过氧化物酶溶液)。通过分别添加20 mmol/L 腐胺溶液以及精胺和亚精胺混合溶液,启动反应来进行DAO 和PAO 测定,在555 nm 处测定吸光值。每30 s 记录1 次,记录5 min。以1 min 内OD降低0.01 为一个酶活力单位(U),活性表示为U·g。
AMADH 活性测定参照Šebela 等的方法稍作修改,取1 g 南瓜样品,加入100 mmol/L 磷酸钾缓冲液(2 mmol/L 二硫苏糖醇,乙二胺四乙酸,10%蔗糖),12000×g 高速离心30 min,收集1 mL 酶液加入1 mL Tris-HCl 反应液和4-氨基丁醛溶液用以启动反应,37 ℃条件下孵育10 min 后,在340 nm 处测定吸光值。每1 min 记录1 次,记录10 min。以1 min内OD降低0.01 为一个酶活力单位(U),活性表示为U·g。

荧光定量使用Vazyme ChamQ TM Universal SYBR qPCR Master Mix 试剂盒,采用2法来计算相对表达量。内参基因采用(XM_023077371.1),5'-AAACCTTACCAGCCCTTGAC-3'(正 向)和5'-CGCTCGTTATAGGACTTGACC-3'(反向)(XM_023082806.1)为5'-CCGTCGAAGGCCGATATCAA-3'(正向)和5'-AGCTCATCTGCTCTCACTTCAC-3'(反向),实验重复3 次。
1.3 数据处理
每次实验进行三次生物学重复,使用SPSS18.0(SPSS,Chicago,IL,USA)统计软件对试验数据进行方差分析,采用Duncan’s Range Test(<0.05)分析数据之间的差异性,数值以平均值±标准误差表示。使用不同的字母表示同一贮藏时间处理方式之间存在显著性差异,并用Origin 10.0 进行绘图。
2 结果与分析
2.1 外源GABA 和3-MP 对鲜切南瓜品质的影响
在鲜切果蔬加工工艺中,切割过程容易导致呼吸强度增加,鲜切表面更容易受到微生物感染,保质期缩短,因此菌落总数是用来评价果蔬安全性和货架期的重要指标。由图1A 可知,在贮藏过程中,对照组和处理组中菌落总数无显著差异(>0.05)。在贮藏9 h 后,南瓜中的菌落总数均未超过10CFU·g,说明南瓜符合可食用和商业标准。白度作为衡量鲜切南瓜贮藏期间新鲜程度的重要指标之一。如图1B 所示,鲜切和外源处理组WI 值均无明显变化。另外,研究发现南瓜果实中含有丰富的-胡萝卜素,使南瓜富含营养物质并呈现丰富鲜艳的颜色。胡萝卜素的含量差异也会导致果肉发生颜色变化。由图1D 可知,三组南瓜样品中的-胡萝卜素含量不存在显著性差异(>0.05)。根据WI 和-胡萝卜素的数据趋势可知,外源GABA 不会影响果肉的色泽,从而影响消费者的购买欲。可溶性固形物是影响果实风味和营养的主要成分。对照组和GABA 处理组的可溶性固形物含量在贮藏期间没有显著变化(>0.05),而抑制剂组的含量显著低于对照组(<0.05)。该结果表明GABA 处理能较好地维持南瓜果实中可溶性固形物含量。综上所述,外源GABA 条件下,鲜切南瓜在贮藏期内的品质并未受到影响,且符合国际标准。
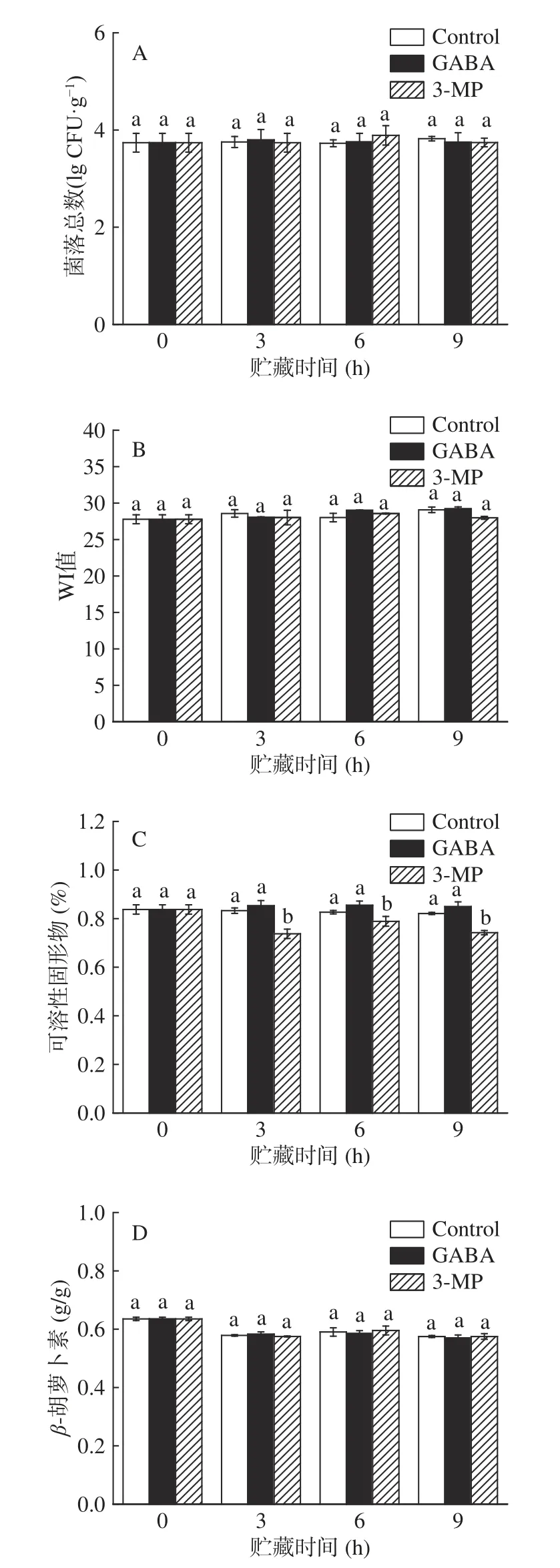
图1 外源GABA 和3-MP 处理对鲜切南瓜中菌落总数、白度、可溶性固形物和β-胡萝卜素含量的影响Fig.1 Effect of exogenous GABA and 3-MP treatments on the total number of colonies,whiteness,soluble solids and βcarotene content in fresh-cut pumpkins
2.2 外源GABA、3-MP 对谷氨酸和GABA 含量的影响
谷氨酸作为合成GABA 的前体物质,植物体内GABA 的富集受到谷氨酸浓度的影响。由图2A可知,对照组和外源3-MP 组的谷氨酸含量在贮藏期间不断下降,而外源GABA 组中的谷氨酸含量则呈现先下降后上升的趋势。与对照组相比,GABA 处理组谷氨酸含量显著低于对照组和抑制剂处理组(<0.05),3-MP 处理后的南瓜丝在整个贮藏期间都保持着较高的谷氨酸含量。这表明,外源GABA 可以促进更多的谷氨酸向GABA 合成和转化。为了进一步研究外源GABA 对内源性GABA 的影响,本文测定了鲜切南瓜中GABA 的含量,由图2B 所示,随着贮藏时间的延长,GABA 处理鲜切南瓜中GABA含量呈现先上升后下降的趋势,而对照组中GABA含量整体无明显变化,经GABA 处理组在6 h 时的GABA 含量达到最大值,约7 mmol·10 gFW,且显著高于对照组中GABA 含量57%(<0.05)。而外源3-MP 处理组的GABA 含量显著低于对照组(<0.05),这说明3-MP 具有抑制内源GABA 合成的作用。与本文结果相似的是,外源GABA 降低了采后西葫芦果实和苹果中谷氨酸的含量,提高了GABA含量。
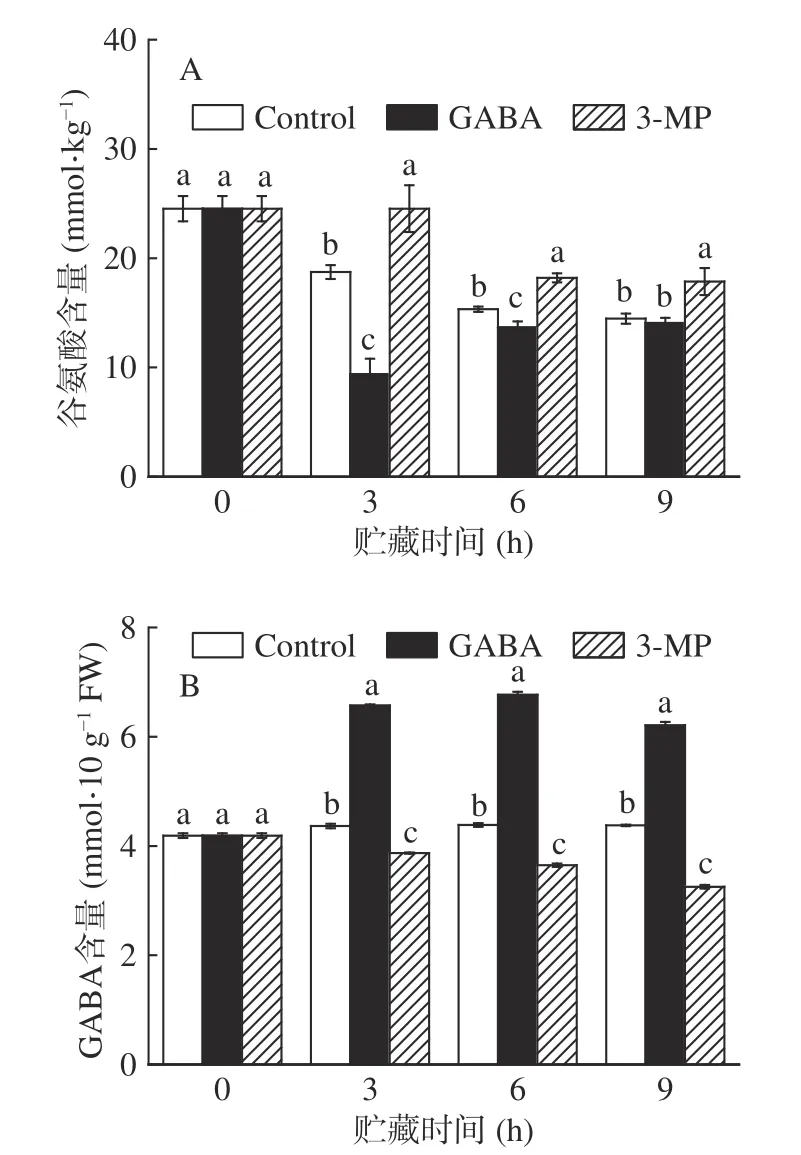
图2 外源GABA 和3-MP 处理对鲜切南瓜中谷氨酸和GABA 含量的影响Fig.2 Effect of exogenous GABA and 3-MP treatments on glutamate and GABA contents in fresh-cut pumpkins
2.3 外源GABA、3-MP 对GAD 和GABA-T 活性的影响
GABA 支路中,GAD 是GABA 支路中GABA合成的限速酶。如图3A 所示,对照组中的GAD呈现缓慢上升的趋势,外源GABA 组在贮藏期间呈现先上升后下降的趋势,与对照组相比,维持较高的GAD 活性。这表明GABA 处理可以提高南瓜中GAD 的活性,促进体内谷氨酸转化成GABA。而抑制剂处理后,南瓜中GAD 活性显著(<0.05)低于对照组和GABA 处理组(图3A)。另外,GABA 的积累不是GAD 的单一作用,GABA-T 能催化GABA和琥珀酸半醛的可逆转化,因此GABA-T 活性的变化对GABA 的富集也同样至关重要。如图3B 所示,GABA 处理组和对照组中GABA-T 活性在贮藏期间呈缓慢上升的趋势,与对照组相比,GABA 处理组GABA-T 活性无显著变化(>0.05),而3-MP 处理显著抑制了GABA-T 活性(<0.05),维持在较低水平。这种影响趋势与GABA 处理对内源GABA含量变化趋势基本一致。在研究杨树、人参和烟草遭受胁迫时,都观察到了类似的结果。因此,综合两个酶指标来看,GABA 处理能促进GAD 和GABA-T 酶活性,激活GABA 支路,从而一定程度上促进鲜切南瓜富集大量GABA 以应对胁迫。因此,本文认为外源GABA 处理诱导的内源性GABA 积累是由GABA-T 和GAD 活性增加所引起的。

图3 外源GABA 和3-MP 处理对鲜切南瓜GAD 和GABA-T 活性的影响Fig.3 Effect of exogenous GABA and 3-MP treatments on GAD and GABA-T activities in fresh-cut pumpkins
2.4 外源GABA、3-MP 对多胺含量的影响
植物响应环境胁迫与多胺的分解代谢有关,在胁迫条件下,Put、Spd 和Spm 会氧化生成GABA。如图4 所示,外源GABA 处理组中腐胺含量在前3 h急速下降,在后续6 h 无明显变化,GABA 处理组中腐胺含量显著低于对照组(<0.05),对照组和3-MP 处理组中腐胺含量均呈先上升后下降的趋势,6 h前两组无明显差异(>0.05),在6 和9 h 出现显著性差异(<0.05)(图4A)。如图4B 所示,在整个贮藏过程中,对照组和两个处理组精胺含量呈先上升后下降趋势,除前3 h 无显著差异外(>0.05),在后6 h中对照组中精胺含量显著高于外源GABA 组(<0.05)。抑制剂处理提高了精胺含量,在贮藏期间保持了高于对照组的水平。GABA 处理组中的亚精胺含量在整个贮藏过程中逐渐降低,对照组呈现先下降再上升,然后下降的变化趋势,而抑制剂处理组中的含量呈现先下降后上升的趋势,且与对照组相比,外源GABA 组中亚精胺含量显著降低(<0.05)。与本文结果相似的是,外源GABA 可以通过激活多胺降解途径,改变多胺生物合成能力,调节了游离Spd 和Spm 与游离Put 的比例,一定程度的促进甜瓜幼苗内GABA 含量增加。因此,本文认为外源应用GABA 可能是通过刺激多胺代谢,降低内源多胺水平,促进GABA 在南瓜体内富集,从而维持鲜切南瓜采后品质。
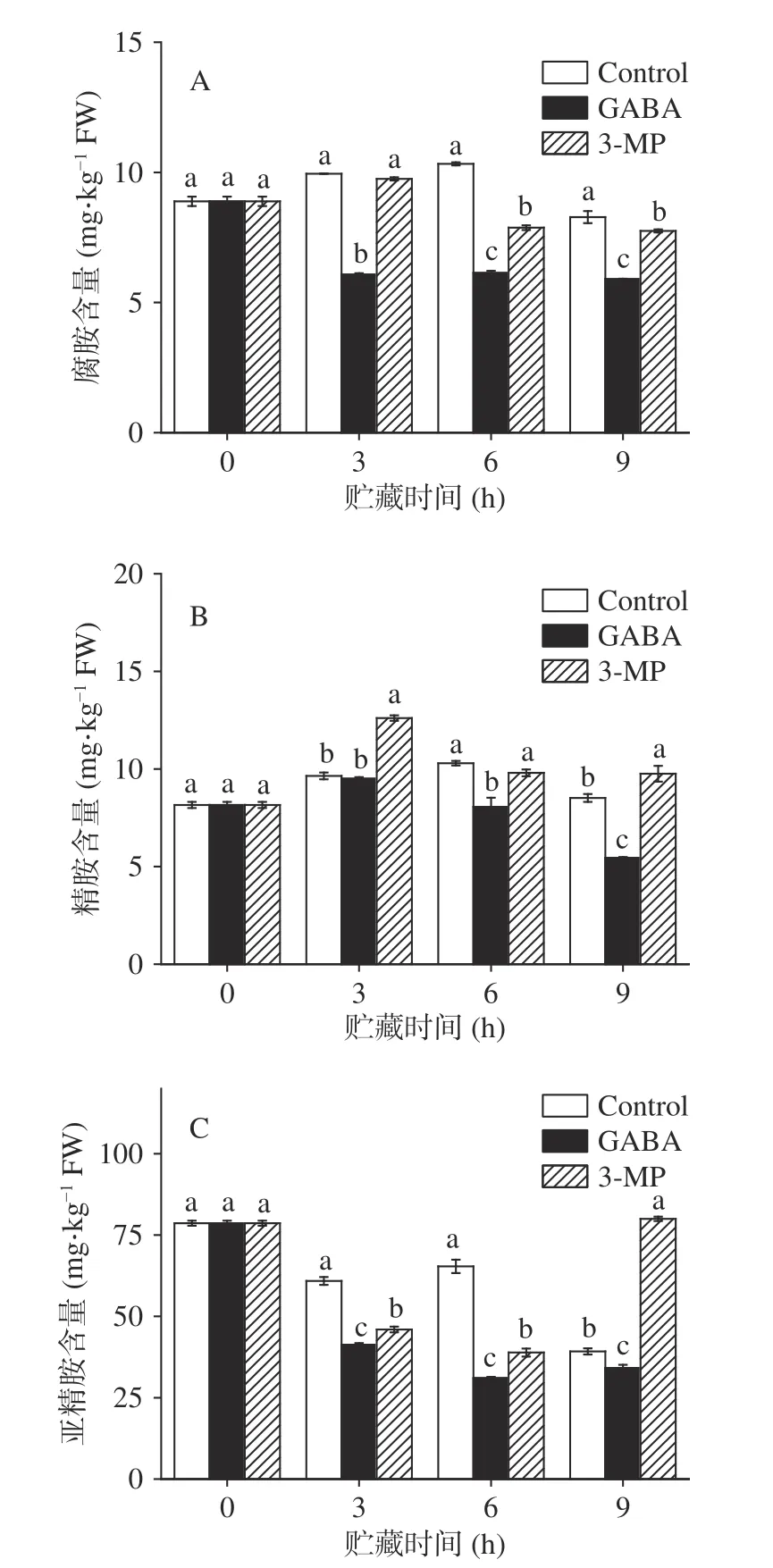
图4 外源GABA 和3-MP 处理对鲜切南瓜中多胺含量的影响Fig.4 Effect of exogenous GABA and 3-MP treatments on the content of polyamines in fresh-cut pumpkins
2.5 外源GABA、3-MP 对DAO、PAO 和AMADH活性的影响
当植物受到胁迫时,腐胺、精胺和亚精胺可直接或间接被DAO、PAO 和AMADH 等氧化酶氧化生成GABA。胺氧化酶协同作用参与植物中多胺降解,可以维持植物中GABA 的内源性水平。如图5A 所示,在贮藏期间,对照组中DAO 活性呈缓慢下降趋势,而两处理组DAO 活性先下降后升高。GABA 处理组的DAO 活性除第9 h 外,其余与对照组无显著差异(>0.05)。而3-MP 处理组的DAO 活性始终显著低于对照组(<0.05)。对照组和外源GABA 组中的PAO 活性无显著差异(>0.05)。和其他组相比,抑制剂组呈现较低的酶活性(图5B)。对照组和GABA 处理组的AMADH 活性在贮藏期间呈现先上升后下降的趋势。与对照组相比,GABA处理显著提高了AMADH 活性(<0.05),且AMADH活性始终高于对照组,而3-MP 处理显著抑制了AMADH 活性(<0.05)。
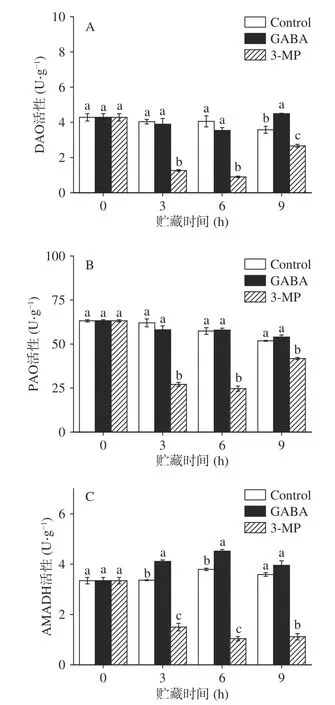
图5 外源GABA 和3-MP 处理对鲜切南瓜中DAO、PAO 和AMADH 活性的影响Fig.5 Effects of exogenous GABA and 3-MP treatments on the activities of DAO,PAO and AMADH in fresh-cut pumpkins
虽然GABA 处理对DAO 和PAO 的酶活性影响不大,但根据多胺含量和AMADH 的变化趋势,推测外源GABA 处理虽然可以激活多胺降解途径,增加GABA 含量,但主要还是通过GABA 支路富集内源GABA。与本文结果相似的是,在大豆发芽期间,虽然DAO 和AMADH 的活性无显著变化,通过促进多胺降解途径,仍提高了其GABA 含量。近期有研究指出,通过增强多胺降解途径催化酶活性,促进多胺的代谢能够迅速积累GABA 并参与防御反应系统,但多胺降解途径合成GABA 的能力远低于GABA 支路。综合以上结论,本文认为GABA的富集有多胺代谢途径的参与,只是对GABA 积累的贡献率较低。
2.6 外源GABA、3-MP 对CmGAD 基因表达的影响
如图6 所示,对照组和处理组南瓜中表达量均呈现先上升后下降的趋势,在6 h 到达峰值。在贮藏期间,GABA 处理组相对表达量显著高于对照组(<0.05),而3-MP 处理组其表达量显著低于对照组(<0.05)。基因表达量与GAD 在贮藏期间活性变化趋势相似。同样也与上调西兰花芽菜中、以及杨树中基因的表达量可以促进内源GABA 含量的结果一致。因此,本文认为外源GABA 可以通过调控表达量,促进内源GABA 的作用。

图6 外源GABA 和3-MP 处理对鲜切南瓜中CmGAD 基因表达的影响Fig.6 Effect of exogenous GABA and 3-MP treatments on CmGAD gene expression in fresh-cut pumpkins
3 结论
结果表明,外源GABA 条件下,促进GABA 支路中谷氨酸含量显著上升,提高了GAD、GABAT 的活性和的基因表达量,促使足够多的谷氨酸向GABA 转化,增强GABA 支路的合成效率。多胺降解途径中,多胺(Put、Spd、Spm)的转化率显著提高(<0.05),虽然DAO 和PAO 的活性与对照组没有显示出较为明显的差异,但AMADH 的活性在贮藏期间始终高于对照组,加速了多胺转化富集成GABA,说明多胺途径同样得到了激活。而3-MP 抑制剂处理则出现了完全相反的效果,GABA 的含量远不如对照组。综上所述,GABA 支路和多胺代谢在外源GABA 诱导鲜切南瓜内源GABA 具有协同作用。此外,GABA 处理是一种安全、健康、环保的果蔬处理方式,可以维持鲜切南瓜中可溶性固形物和颜色的变化,抑制菌落总数上升,防止鲜切南瓜受到微生物的侵染,减少-胡萝卜素的流失,有助于保持南瓜的品质。本实验并未从多胺分子层面进行深入研究,这也可能是导致某些酶活性能并未表现出显著提高的原因之一,若能进一步研究,将为GABA处理采后果蔬投入市场应用提供更加可靠的理论依据。
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
中国土壤与肥料(2021年5期)2021-12-02
中国饲料(2021年17期)2021-11-02
农业资源与环境学报(2021年5期)2021-10-06
阅读与作文(小学高年级版)(2019年5期)2019-05-27
分析化学(2019年3期)2019-03-30
复旦学报(医学版)(2016年6期)2016-12-20
医学研究杂志(2015年12期)2015-06-10
食品工业科技(2014年21期)2014-03-11
