
不同樱属植物叶形态与解剖结构的比较
2022-09-24 06:50柏文富张文昕聂东伶李建挥严佳文吴思政
中南林业科技大学学报 2022年8期
柏文富,张文昕,禹 霖,聂东伶,李建挥,熊 颖,严佳文,吴思政
(1.湖南省植物园,湖南 长沙 410116;2.中建文化旅游发展有限公司,北京 100037)
樱属Cerasus植物隶属蔷薇科Rosaceae 李亚科Prunusoidea,全球约150 种,分布于欧亚大陆至北美洲的北半球温带[1],大多数樱花既有桃之艳丽,又有梅之芳香,是著名的春季木本观花植物和秋色叶树种[2-4]。虽然分子标记在一定程度上能准确快速地反映物种的遗传多样性,但形态标记在植物分类学上仍发挥着无法替代的作用[5-6]。叶片是植物输送水分和进行物质交换的主要器官,是植物进行同化作用和光合作用的场所,其形态特征和解剖结构特征是其在长期特定环境条件下适应并演化的结果,不会随环境因子的暂时改变而发生较大的变化[7]。因此,叶片能反映环境因子的影响程度及植物对环境条件的适应能力[8-10]。叶片表皮细胞、气孔性状和表皮附属物是植物叶片解剖研究中的主要内容[11]。叶肉是叶片进行光合作用的重要场所,在叶片结构中变异最大,叶片的气孔密度、气孔长宽比等性状的变异则较小,属于相对稳定的系统演替性状[12]。植物的种(品种)不同,叶脉维管系统的形状与隆起程度也有所不同,这也使叶片形态特征和解剖结构特征成为系统分类学中植物属种间划分与判别、系统演化以及对环境适应能力的重要依据之一[13]。
目前,对植物的叶形态及解剖结构研究主要是根据叶片解剖特征的数量性状探讨其抗性的关系,如对5 个楸树无性系[14]、金花茶组植物[15]、238 份板栗品种[16]、3 个芭蕉芋品种[9]、5 个沙棘品种[17]和6 个卫矛属植物[18]的抗旱研究,对6个油茶品种[19]、7种番荔枝果树的抗寒研究[20],对70 份武夷名丛茶的抗虫、抗旱、抗寒研究[10]等。通过叶片解剖特征进行对比分类分析的文献较少,仅见对中国肉桂与越南清化肉桂叶解剖结构对比分析[21]。樱属植物的分类研究主要是根据樱花的花器官、叶、果形态特征和生活型进行分类。例如,根据樱属植物的生活型、叶、花形态特征对中国李属樱亚属进行数量分类,将41 个种划分成11 个类群[22];根据我国樱属植物的资源分布生物学表型性状,将樱属51 个种和12 个变种划分为2个亚属,4 个组,设11 个系[23]。对樱属植物不同种和品种间的叶片解剖结构进行分类特征对比分析的相关基础研究未见报道。本研究以樱属7 个种和9 个品种共16 份樱属植物的成熟叶为试验材料,通过叶片形态学、植物显微技术观测其叶片形态特征与解剖结构,分析樱属不同植物叶片形态和解剖结构差异,为樱属植物分类研究提供科学依据。
1 研究地概况
研究地位于湖南省植物园樱花国家林木种质资源库种质资源保存区,中心位置为(113°01'27″E,28°06'22″N),海拔78.5~105.7 m,属于典型中亚热带季风湿润气候,四季分明,年均气温17.2℃,极端高温40.6℃,极端低温-11.3℃,年平均降雨量1 412.3 mm,平均相对湿度80%,日照时数年平均1 726 h,无霜期281 d。土壤为四纪红壤,pH 值4.5~5.5,坡度15°左右,土层深厚,肥力较好,适宜樱属植物生长发育。
2 材料与方法
2.1 试验材料
本试验材料来源见表1,采集于湖南省植物园樱花国家林木种质资源库种质资源保存区的尾叶樱桃等16 份樱属植物,试材取自健康植株上生长健壮、无病虫害、向阳枝条中部的成熟叶片。

表1 樱属植物来源Table 1 Cerasus materials
2.2 试验方法
2.2.1 叶形态特征的观测
利用LI-3000C 叶面积仪测定每一份樱属植物资源的30 片成熟叶片的叶长、叶宽和叶面积。
2.2.2 组织透明法
将每份材料的30 片成熟叶片分为5 组,每组6 片,用单面刀片于叶片中间沿主脉两侧切取0.5 cm×0.5 cm 大小的方块,立即投入标准固定液(70%酒精∶冰醋酸∶甲醛=18 ∶1 ∶1,V/V),抽气30 min,48 h 后置于约为材料体积20 倍的Jeffrey 离析液(10%铬酸∶10%硝酸=1 ∶1,V/V)中,放入40℃恒温箱约24 h 后倒去离析液。在放有清水的培养皿中利用镊子、毛刷和解剖针等撕取叶表皮,置于1%的番红染液中染色1 min。去除番红染液,清水多次反复清洗,制作临时切片,用Motic 数码显微镜(Motic B5 Professional Series)及图像分析系统(Motic Images Advanced 3.0)观察并拍照,每种试材各随机选择10 张图片,用Digimizer 软件测量相关指标。
2.2.3 石蜡切片法
用单面刀片于叶片基部三分之一处沿主脉横切,切取0.5 cm×1.0 cm 大小的方块,立即投入标准固定液,抽气30 min,固定48 h。按常规石蜡制片技术(10 μm),梯度乙醇脱水,番红固绿对染,二甲苯透明,中性树胶封片,制成永久性切片于Motic V 1 数字切片扫描系统中进行显微观察并拍照,在图像测量软件Digimizer 下测定叶片解剖结构参数,重复15 次后取其平均值。
2.3 数据分析
采用Microsoft Excel 2013 和SPSS 17.0 软件对数据进行单因素方差分析,多重比较采用Duncan氏新复极差检验(P<0.05)。

3 结果与分析
3.1 不同樱属植物试材的叶片形态特征的比较
樱属植物的叶有叶柄、叶片和托叶,叶缘有锯齿,叶柄、托叶和锯齿前端常有腺体。叶柄腺体着生位置、腺体形状和数量、叶片气孔器性状和大小、表皮毛被情况、角质层纹饰等具有丰富的变异,多样性高,对樱属植物的分类具有重要的参考价值[24-25]。不同樱属植物的叶形态如图1所示。叶片边锯齿形态如图2所示,叶的形态特征参数见表2。

图1 不同樱属植物叶的形态特征Fig.1 Morphological characteristics of different Cerasus leaves

图2 不同樱属植物试材叶边锯齿Fig.2 Characteristics of leaf edge serration of different Cerasus leaves
樱属植物的叶片多为长椭圆形或倒卵状长椭圆形,先端尾尖,均有叶柄。由图1可知,樱属植物叶片的变异较大。
由图1~2 和表2可知,16 个樱属植物试材叶片边缘均有锯齿,且各有特点,各种间差异较大,相关的品种间相近或差异较小。除钟花樱、‘寒绯樱’和‘阳光樱’齿端腺体较少外,其他叶片齿端有明显腺体,‘寒绯樱’是日本选育的钟花樱桃,‘阳光樱’是‘天城吉野’Cerasus yedoensis cv.Amagi-yoshino 与‘寒绯樱’杂交育成;大岛樱、‘松月樱’‘花笠樱’‘郁金樱’‘天之川樱’的锯齿均为芒状腺齿,大岛樱是山樱花的一个类群,其他为山樱花与其他樱花的杂交品种,尾叶樱桃、迎春樱桃、高盆樱桃、华中樱桃、‘杭州早樱’、大岛樱、‘修善寺寒樱’‘花笠樱’‘郁金樱’和‘天之川樱’的锯齿为混合锯齿。钟花樱桃和‘寒绯樱’为斜向上尖锐单锯齿,‘染井吉野’‘阳光樱’和‘松月樱’为重锯齿,尾叶樱桃、迎春樱桃、大叶早樱、‘杭州早樱’‘染井吉野’叶片两面均有毛被,‘阳光樱’仅背面有少量柔毛,其他樱花叶片均无毛。16 种樱属植物试材的叶片形态特征参数的方差方析和多重比较见表3。由表3可知,华中樱桃、尾叶樱桃、钟花樱桃、高盆樱桃、大叶早樱、迎春樱桃和大岛樱、‘染井吉野’彼此之间的叶柄、叶长、叶宽、长宽比和叶面积均有较显著性差异,选育或杂交的品种与原种之间差异不大。‘郁金樱’叶柄最长,平均3.04 cm,迎春樱桃叶柄最短,仅0.63 cm;‘花笠樱’平均叶长最长,为14.38 cm,迎春樱桃平均叶长最短仅7.57 cm;华中樱桃平均叶宽最宽为6.80 cm;‘杭州早樱’平均叶宽最狭窄仅3.77 cm;‘染井吉野’叶片长宽比最小为1.87;大叶早樱叶片长宽比最大为2.48,叶片细长;平均叶面积以华中樱桃最大,为60.14 cm2;以‘杭州早樱’最小,为19.10 cm2。‘杭州早樱’是从迎春樱桃中选育出来的品种,两者的叶长、叶宽、叶片长宽比和叶面积均无显著性差异。‘松月’‘天之川’是由山樱花与其他樱花杂交育成,它们的叶柄、叶长、叶片长宽比和叶面积均无显著性差异。钟花樱桃、‘寒绯樱’‘阳光’亲缘关系近,它们之间的以上性状也差异不大。

表2 不同樱属植物的叶形态特征Table 2 Leaf morphology of different Cerasus

表3 不同樱属植物的叶片形态特征参数分析†Table 3 Analysis of morphological characteristics of different Cerasus leaves
3.2 不同樱属植物的叶片表皮微形态特征的比较
16 个樱属植物的不同种的叶片表皮微形态特征差异较大,来源于较近亲缘关系的品种间差异较小,叶片表皮微形态特征可以作为樱属植物种间鉴定和分类的重要依据。
3.2.1 叶片表皮细胞特征
不同樱属植物叶片的上下表皮离析样本细胞特征如图3~4 所示。通过观察叶片表皮离析样本,发现16 个樱属植物叶片上下表皮除尾叶樱桃密被长表皮毛,迎春樱桃、‘杭州早樱’、大叶早樱和‘染井吉野’有短表皮毛外,其他试材叶片均无表皮毛。对叶片表皮离析样本分析(表4),发现仅华中樱桃与钟花樱桃的上表皮细胞呈无规则形,其余14 份樱花的叶片上表皮细胞均呈多边形;下表皮细胞形状仅尾叶樱桃呈多边形,其余15 份樱花叶片下表皮细胞均呈不规则形。不同樱属植物上表皮的垂周壁式样有弓形和平直2 种。16 份樱花叶片中仅迎春樱桃、大叶早樱和‘杭州早樱’的上表皮垂周壁式样是弓形(其中,‘杭州早樱’是从迎春樱桃中选育出来的),其余13 份樱花叶片的上表皮垂周壁式样均为平直;下表皮的垂周壁式样有弓形和波状2 种。16 份樱花叶片中仅钟花樱桃、大叶早樱、‘花笠樱’和‘郁金樱’的下表皮垂周壁式样是波状,其余12 份樱花叶片的下表皮垂周壁式样均为弓形。
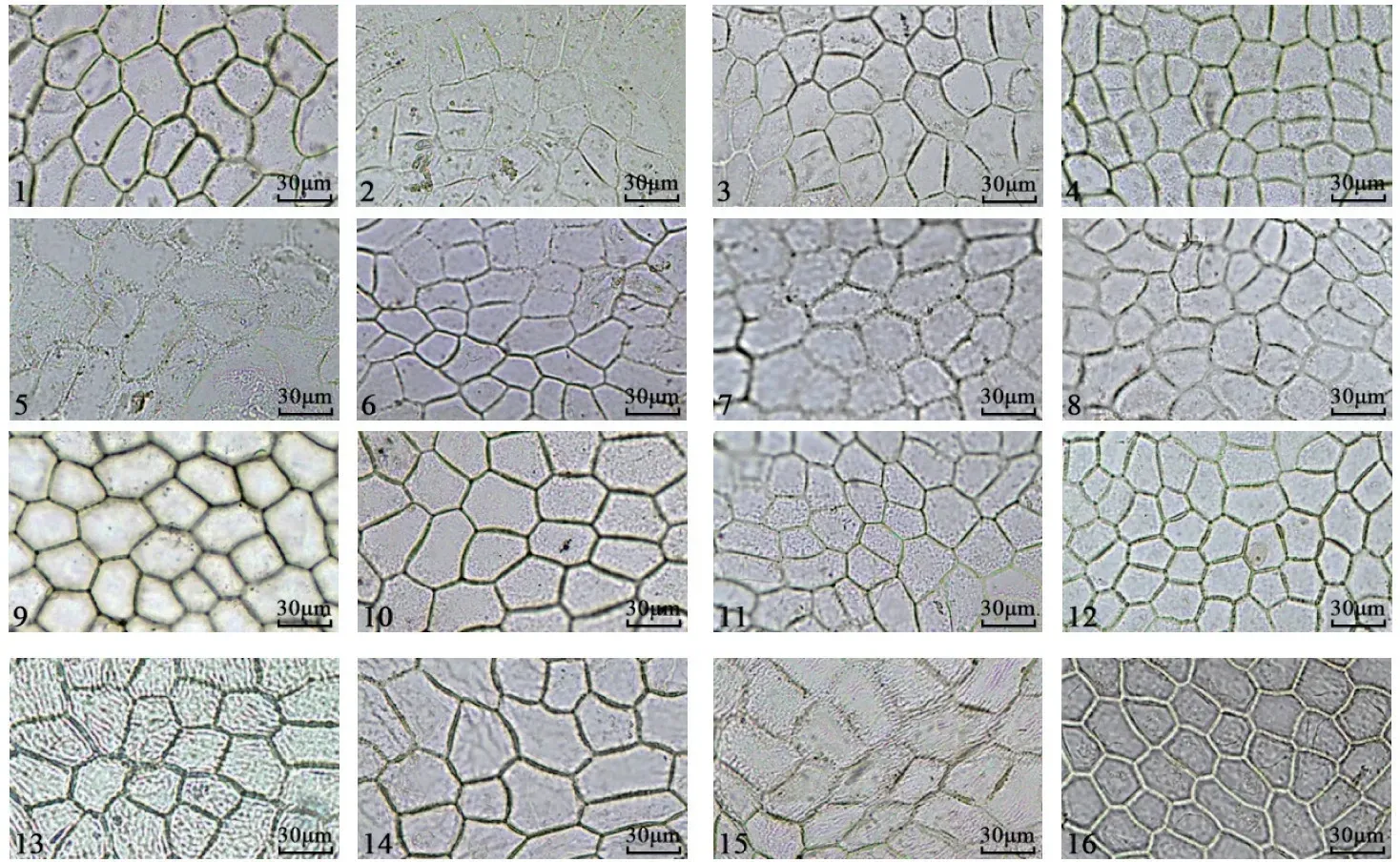
图3 不同樱属植物叶片上表皮细胞特征Fig.3 Characteristics of upper epidermal structures of different genera of Cerasus leaves
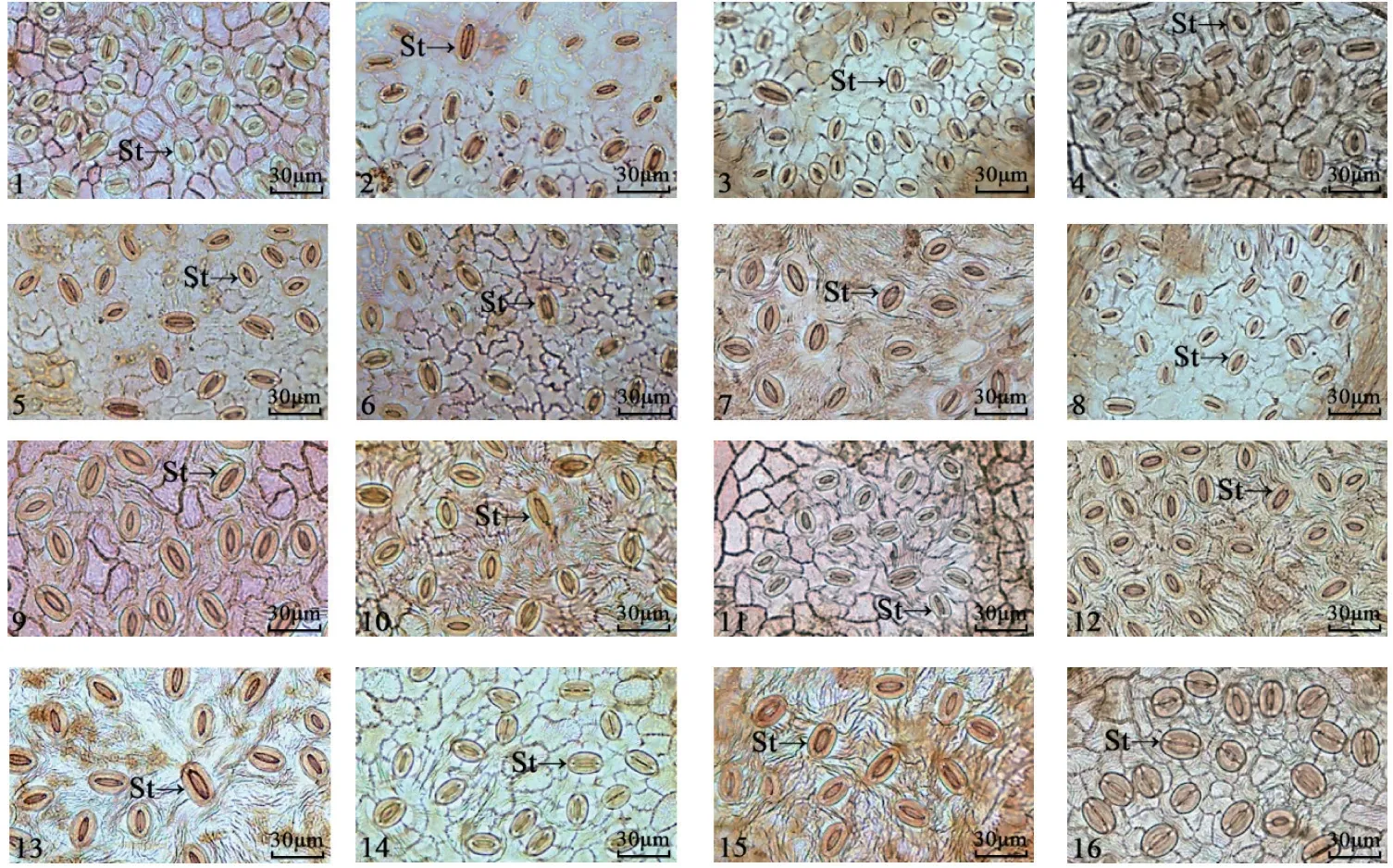
图4 不同樱属植物叶片下表皮细胞及气孔特征Fig.4 Characteristics of lower epidermal structures and stomatal characteristics of different Cerasus leaves
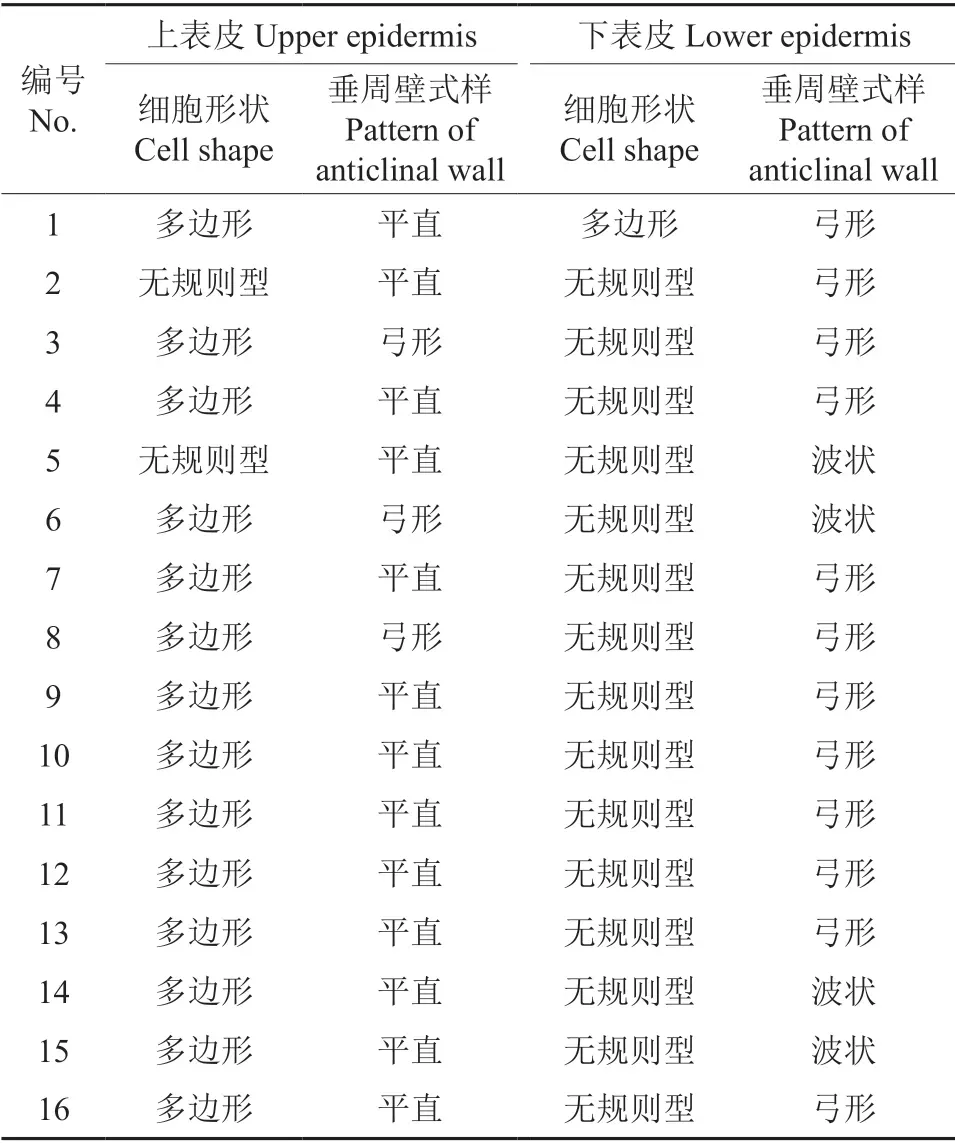
表4 不同樱属植物叶表皮细胞特征Table 4 Characteristics of leaf epidermal cells of different Cerasus leaves
3.2.2 气孔特征
在光学显微镜下16 份樱花叶片的表皮细胞外均有角质层结构,上表皮中未发现气孔存在,在下表皮中均观察到气孔的存在。气孔器由2 个对称排列的肾形保卫细胞组成,形状大多为椭圆形,呈不规则排列。不同樱花叶片之间的气孔大小差异显著(P<0.05,表5)。气孔长度以‘修善寺寒樱’与‘松月樱’的较长,分别为27.00、26.64 µm,显著高于其他樱花叶片的气孔长度;‘寒绯樱’‘杭州早樱’、尾叶樱桃和迎春樱桃的气孔的长度则较 短,分别为18.22、17.58 17.48、17.25 µm,显著低于其他樱花叶片的气孔长度;‘修善寺寒樱’与‘松月樱’的气孔长度分别约为‘寒绯樱’‘杭州早樱’,尾叶樱桃和迎春樱桃的1.5 倍。气孔宽度以‘天之川樱’的最宽,达19.28 µm,显著高于其他樱花叶片气孔宽度;迎春樱桃的气孔最窄,仅为11.62 µm,显著低于其他樱花叶片的气孔宽度;‘天之川樱’的气孔宽度约为迎春樱桃的1.7 倍。‘松月樱’‘修善寺寒樱’‘天之川樱’和‘郁金樱’的单个气孔面积显著大于其他樱花叶片的单个气孔面积,但这4 个试材间的差异未达显著性水平。气孔密度的种间差异明显(P<0.05)。迎春樱桃和‘杭州早樱’的气孔密度最大,分别为667.92 和652.43 个/mm2,显著高于其他樱花叶片的气孔密度;‘郁金樱’的气孔密度最小,为250.64 个/mm2;迎春樱桃和‘杭州早樱’的气孔密度约为‘郁金樱’的气孔密度的2.6 倍。16 个樱属植物叶片中,尾叶樱桃、钟花樱桃、大叶早樱、‘修善寺寒樱’和‘天之川樱’气孔器保卫细胞两极有“T”形加厚。
3.3 不同樱属叶片解剖结构特征的比较
樱属植物的叶片解剖结构特征可以作为种间鉴定和分类的重要依据。16 个樱属植物的叶片均为背腹型,叶横切面形态结构和叶解剖结构基本相同,均为异面叶,明显分化为表皮、叶肉和叶脉3 个部分。叶片组织结构从下往上,依次为下表皮、海绵组织、栅栏组织和上表皮(图5)。从表5可知,‘花笠樱’的叶片最厚(193.56 µm),高盆樱桃、大岛樱、‘松月樱’和‘郁金樱’次之(171.07~182.20 µm),‘染井吉野樱’较厚(154.93 µm),尾叶樱桃、华中樱桃、钟花樱桃、大叶早樱、‘修善寺寒樱’‘阳光樱’和‘天之川樱’较薄(133.00~144.46 µm),迎春樱桃和‘杭州早樱’最薄(99.30~101.57 µm)。
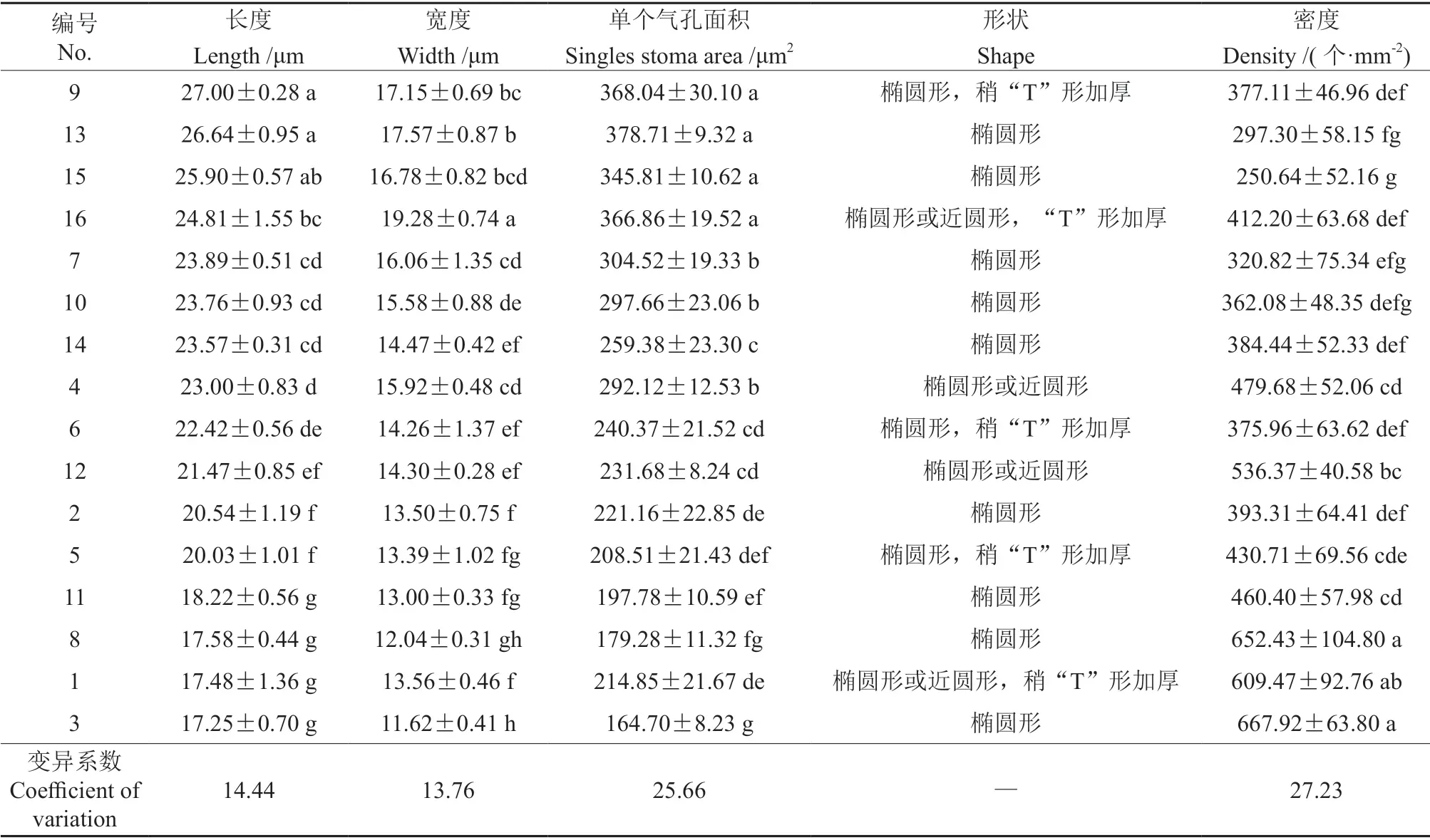
表5 不同樱属植物叶片的气孔特征†Table 5 Characteristics of stomata of different Cerasus leaves
3.3.1 叶表皮结构特征
从图5可知,16 个樱属植物试材叶片的表皮除‘郁金樱’的上表皮由两层细胞构成外,其他都由单层近矩圆细胞构成的很薄的角质层。高盆樱桃、大岛樱和‘松月樱’叶上表皮由大型细胞组成,表皮厚度分别为32.23、34.71 和29.40 µm;而迎春樱桃和‘杭州早樱’上表皮细胞很小,表皮厚度仅为12.42 和11.40 µm。下表皮以‘花笠樱’的最厚,为16.21 µm;尾叶樱桃的下表皮最薄,为7.44 µm。
3.3.2 叶肉结构特征
由图5可知,16 个樱属植物的叶肉由海绵组织和栅栏组织构成,栅栏组织位于海绵组织上方,由1~2 层排列紧密的呈柱状的薄壁细胞构成,海绵组织细胞排列无规则略疏松,有明显细胞间隙。从表6中可知,16 个樱属植物的叶片中,高盆樱盆、‘修善寺寒樱’‘染井吉野樱’、大岛樱、‘花笠樱’的栅栏层为2 层,其中高盆樱桃和‘花笠樱’的栅栏组织最厚,分别为65.37 和69.40 µm,显著高于其他樱属植物叶片的栅栏组织厚度;尾叶樱桃虽然只有1 层栅栏层,但其栅栏组织比有2 层栅栏层的‘寒绯樱’和‘修善寺寒樱’厚;华中樱桃、迎春樱桃、钟花樱桃、‘杭州早樱’叶片的栅栏组织最薄,为21.70~27.41 µm。海绵组织厚度在不同樱属植物叶片间的差异明显(P<0.05):‘郁金樱’叶片的海绵组织最厚,达99.85 µm,显著高于其他樱属植物叶片的海绵组织;尾叶樱桃、华中樱桃、迎春樱桃、钟花樱桃、‘寒绯樱’‘杭州早樱’的叶片海绵组织厚度相对较低,仅为50.73~54.81 µm。16 个樱属植物的叶肉组织细胞排列紧密程度有差异:尾叶樱桃和高盆樱桃的叶肉组织细胞排列紧密(图5-1 与图5-4),而华中樱桃、钟花樱桃、大叶早樱、‘松月樱’和‘郁金樱’的叶肉细胞排列则较稀疏(图5-2、图5-5、图5-6、图5-13 和图5-15)。16 个樱属植物中高盆樱桃的栅海比最高,达1.01,显著高于其他樱属植物栅海比;迎春樱桃、钟花樱桃、大叶早樱、‘杭州早樱’‘松月樱’‘郁金樱’和‘天之川樱’的栅海比则较小,其范围在0.37~0.48 之间。

图5 不同樱属植物叶片横切面的叶肉结构Fig.5 Mesophyll structures of different Cerasus leaves in transverse section
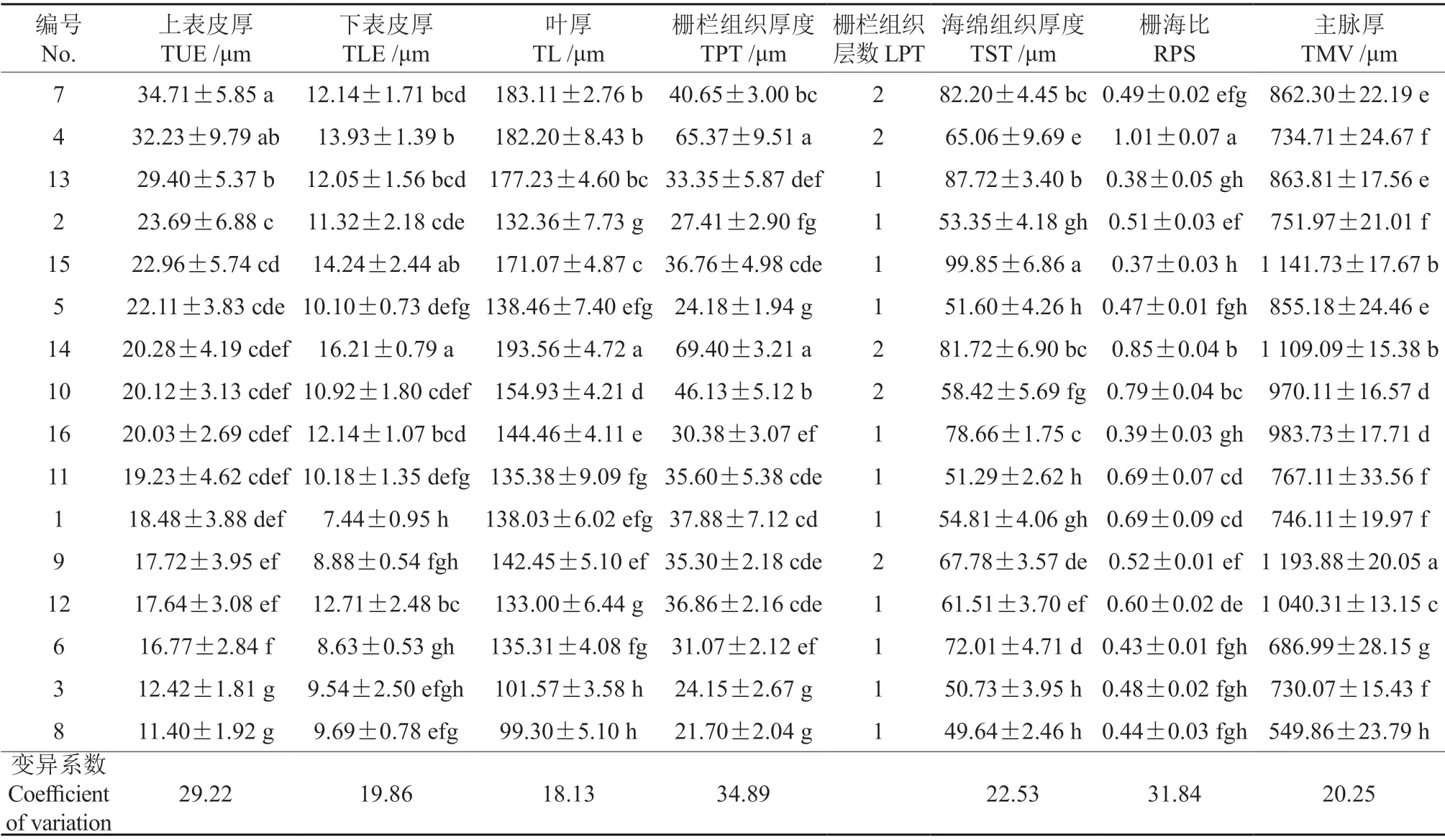
表6 不同樱属植物叶片的主要解剖结构特征†Table 6 The main anatomical structure characteristics of different Cerasus leaves
3.3.3 主脉特征结构
由表5~6可知,16 个樱属植物试材的主脉结构均为开放的半圆形,主脉厚度存在显著性差异。其中,‘修善寺寒樱’主脉最粗,厚为1 193.88 µm;其次是‘郁金樱’和‘花笠樱’,厚度分别为1 141.73 和1 109.09 µm;主脉最小,厚度最薄的是杭州早樱,厚度仅为549.86 µm,不到‘修善寺寒樱’厚度的50%。主脉由厚到薄排序为‘修善寺寒樱’>‘郁金樱’>‘花笠樱’>‘阳光樱’>‘天之川樱’>‘染井野樱’>‘松月樱’>大岛樱>钟花樱桃>‘寒绯樱’>华中樱桃>尾叶樱桃>高盆樱桃>迎春樱桃>大叶早樱>‘杭州早樱’。

图6 不同樱属植物叶片横切面的主脉结构Fig.6 The structure of midrib in transverse sections of different Cerasus leaves
4 结论与讨论
叶片形态特征是植物遗传多样性的表现形式之一,同时也是种间或种以下亲缘关系之间鉴定和分类依据之一。16 个樱属植物试材的叶片除尾叶樱桃叶片上下两面均密被长表皮毛,迎春樱桃、‘杭州早樱’、大叶早樱有短表皮毛外,其他樱花叶片均无表皮毛。形态特征参数中,华中樱桃、尾叶樱桃、钟花樱桃、高盆樱桃、大叶早樱、迎春樱桃、大岛樱和‘染井吉野’彼此之间的叶柄、叶长、叶宽,叶片长宽比和叶面积均有较显著性差异,其他选育或杂交而来的品种与原种之间差异不大。
植物在进化的过程中,叶与周围的环境关联密切[26]。叶片形态和结构因环境和功能的差异而有所不同,叶片解剖结构是植物遗传多样性的重要表现形式[27],不同樱属植物的叶片表皮微形态特征参数和叶解剖结构的参数存在显著差异(P<0.05),可以作为种间鉴定和分类的重要依据。在不同樱属植物叶的各项解剖结构参数中,各项指标的排序都有一定的变化,7 个种间差异显著,其他选育或杂交而来的品种与原种间差异小。华中樱桃与钟花樱桃的上表皮细胞呈无规则形,其他樱花上表皮细胞均呈多边;尾叶樱桃的叶片下表皮细胞呈多边形,其他樱花叶片下表皮细胞均呈不规则形;迎春樱桃、大叶早樱和‘杭州早樱’的叶片上表皮垂周壁式样呈弓形,其余试材均为平直;钟花樱桃、大叶早樱、‘花笠樱’和‘郁金樱’的叶片下表皮垂周壁式样呈波状,其余试材均为弓形;‘松月樱’的单个气孔面积最大,而气孔密度则相对较小;与之相反,迎春樱桃的气孔密度在16 个樱花叶片中是最大的,而其单个气孔长度面积则最小;气孔的形状均为椭圆形,其中尾叶樱桃、钟花樱桃、大叶早樱、‘修善寺寒樱’和‘天之川樱’的气孔器保卫细胞两极有“T”形加厚;尾叶樱桃、‘阳光樱’和‘郁金樱’的栅栏组织虽为1 层,但其厚度大于有两层栅栏组织的‘寒绯樱’和‘修善寺寒樱’;‘寒绯樱’和‘修善寺寒樱’的栅栏组织柱形细胞的长轴较短,但层数较多,这表明植物为保证光合作用与蒸腾作用的正常运行,可能会通过增加细胞层数来弥补其细胞体积较小的特点。叶肉组织均分化为栅栏组织和海绵组织,主脉均为开放的半圆形结构,厚度存在显著性差异(P<0.05)。‘花笠樱’的下表皮厚度、叶片厚度和栅栏组织厚度最厚,‘高盆樱桃’的栅栏组织厚度与海绵组织厚度之比最大。
樱属植物各种之间容易在风媒、虫媒条件下自然杂交授粉,产生丰富的变异和形态多样性,出现大量的中间过渡类型。许多樱属植物代表种与植物志等文献记录的表形特征存在较大差异,同一种质资源在不同居群中也存在表型差异。这些差异给樱属植物在实践中利用形态特征分类带来较大的困扰。2013年发表的新种磐安樱桃[28]通过叶绿体DNA atpB-rbcL 片段测序[29],说明相比浙闽樱桃与尾叶樱桃有更近的亲缘关系。本研究仅对樱属植物的7 个种和9 个品种的不同樱属植物的叶形态和解剖结构特征进行对比分析,以此为依据来对整个樱属的叶形态和解剖结构特征对比分析则存在一定的局限性,以后应扩大对樱属植物叶形态和解剖结构对比分析的种类,同时依据它们的茎、花、果实等形态特征,结合分子标记等技术手段,对樱属植物进行精准分类。
猜你喜欢
航空材料学报(2022年4期)2022-08-04
今日农业(2022年3期)2022-06-05
电影文学(2022年4期)2022-05-20
贵州农业科学(2022年4期)2022-05-06
中国农学通报(2022年6期)2022-04-15
保健与生活(2022年8期)2022-04-08
时代邮刊·下半月(2022年1期)2022-03-22
干旱地区农业研究(2022年1期)2022-01-28
祝您健康·文摘版(2019年4期)2019-06-11
中学生天地(A版)(2019年3期)2019-04-19
