
硒、硅对镉胁迫下小麦生长、生理特性及镉分布的影响
2022-09-23 12:11张明辉时曼丽
江苏农业科学 2022年17期
张明辉, 时曼丽
(南阳市农业科学院,河南南阳 473000)
镉(Cd)是一种剧毒性化学元素,存在于水体和土壤环境介质中。人类活动,包括金属采矿和冶炼、废水灌溉、磷肥施用等是环境Cd污染的主要来源,人类行为产生的Cd含量比与地质自然Cd含量高出3~10倍。Cd是流动性较快的重金属元素之一,并且在水-土壤-植物系统中具有较高的活性;农业生产中向土壤施入Cd是一种净汇过程,这意味着一旦Cd进入到环境介质中,很容易在植物中积累,严重影响植物生长和作物产量,然后通过食物链进入动物体,对人体肝脏、生殖器、心血管、内分泌以及肝脏系统造成损害,从而对环境和人类健康造成不利影响。因此,在农业生产中限制Cd从土壤转移到植物对于食品安全至关重要。
目前,已探索了一系列农艺措施、生物技术及土壤钝化技术来管理和修复Cd污染土壤并控制其在作物中的积累。研究发现,为植物提供有益元素是减少小麦Cd积累和毒性的最具成本效益、环境友好的可持续策略之一。硅(Si)和硒(Se)是广受关注的重要有益元素,大量研究表明,Si、Se对生长发育、产量收获和抵抗各种生物/非生物胁迫皆具有较佳的作用效果。研究表明,Si可以有效减轻不同作物的重金属毒性,如玉米、小麦和水稻。在Cd胁迫下,Si可以调节负责Cd从外部溶液转运到水稻植物根部细胞的相关基因转录水平,从而抑制Cd吸收。在细胞中,Si与半纤维素结合具有净负电荷可抑制Cd吸收,使其在细胞壁上形成共沉积。Cd受到HMA基因家族表达的调节,促进液泡中的Cd分离,以减轻Cd在植物中迁移和运输过程中的毒性。Se是动物必需的营养元素,也是植物的有益营养元素,Se主要通过防止氧化应激,调节光利用率,修复细胞损伤和调节基因表达来降低Cd毒性。
Si和Se的比较研究在研究人员中也越来越受到关注。 Wu等的研究表明,在卷心菜植株中同时喷洒5 μmol/L Si和Se可显著降低卷心菜的根系和地上部中的Cd含量。然而,关于这些必需微量元素的相互作用及其对缓解Cd毒性影响的认识仍然有限。小麦是全球30%以上人口的主要食物,小麦中的高Cd积累导致过量的Cd摄入从而对人类健康造成巨大威胁;因此,减少小麦从土壤中吸收Cd是一个至关重要的粮食安全环节。基于此,本研究探索了Se和Si相互作用对小麦发育代谢、抗氧化系统、Cd累积分布及相关基因表达的影响。
1 材料和方法
1.1 供试材料
试验于2021年2—5月在南阳市农业科学院进行。供试小麦品种为郑麦1354,种子来自河南省农业科学院经济作物研究所。种子采用0.5%次氯酸钠进行表面灭菌15 min,然后用去离子水冲洗数次并浸泡12 h,然后放置于铺垫润湿滤纸的培养皿中,28 ℃培养箱暗处理催芽24 h。
供试镉为氯化镉(CdCl),硒为五水亚硒酸钠(NaSeO·5HO),硅为九水偏硅酸钠(NaSiO·9HO),均购自国药集团。
供试土壤取自河南省南阳市农业科学附近原始土,土壤类型为中性沙壤土。土壤经风干后混匀过 3 mm 网筛,土壤pH值为7.11,土壤有机质含量10.06 g/kg,全氮含量0.85 g/kg,碱解氮含量 42.80 mg/kg,有效磷含量12.23 mg/kg,速效钾含量75.29 mg/kg,全镉含量0.04 mg/kg,根据GB 15618—2018《农用地土壤污染土壤环境质量风险控制标准》,当土壤6.5 试验设置5个处理:CK,不施用镉、硒、硅;Cd处理:仅施用镉,不施用硒、硅;Cd+Si处理:施用镉、硅,不施用硒;Cd+Se处理:施用镉、硒,不施用硅;Cd+Si+Se处理:施用镉、硒、硅。各处理重复4次。其中Cd用量为15 mg/kg,Si、Se单一元素用量皆为4 mg/kg。 盆栽装置为黑色方形塑料盒(20 cm×15 cm×15 cm),每盆装土2.5 kg。Cd、Se、Si皆采用水溶方式提前施入。每盆施入小麦种子20粒,出苗后减至10株。为保证幼苗的正常生长及养分平衡,每周向盒中加入50 mL小麦营养液。共培育35 d,种植期间不定时加入无菌水,其他管理措施同作物培育方法。 1.3.1 小麦植株生物量、Cd浓度及Cd亚细胞分布测定 培养结束后将小麦植株地上部、根系分开,采用直尺测量根长及株高参数,将植株105 ℃杀青30 min,65 ℃烘干至恒质量以计算生物量。Cd浓度采用HNO消解,采用电感耦合等离子体光谱仪(ICP-OES)测定样品中的Cd浓度。 细胞壁(Fcw)、细胞质(Fs)和细胞器(Fo)分级用于确定Cd的亚细胞分布,相关方法参考Xin等的研究并略有修改,即将冷冻小麦植物样品称量到冷砂浆中,并用预冷的250 mmol/L蔗糖和1 mmol/L二硫代甲状腺素醇(DTT)混合缓冲溶液(50 mmol/L Tris-HCl,pH值7.5)提取样品。然后将匀浆以7 000 r/min离心30 s,沉淀物即为Fcw,在上清液以15 000 r/min进一步离心45 min后,上清液溶液和沉积物分别鉴定为Fs和Fo。 1.3.2 光合特征参数及叶绿素含量测定 光合色素包含叶绿素a、叶绿素b均采用丙酮-乙醇混合浸提,采用紫外分光光度计(UV-755B,青岛聚创华业分析仪器有限公司)在665、649 nm处测定吸光度,具体步骤参照李合生等的研究。采用LI-6400便携式光合测定系统(LI-6400;LI-COR,America)测定叶片的净光合速率()、胞间CO浓度()、蒸腾速率()和气孔导度()等指标,叶室温度设置为(25±1) ℃,CO浓度为440 μmol/mol,光量子密度为800 μmol/(m·s)。 1.3.3 丙二醛(MDA)、抗坏血酸(AsA)、谷胱甘肽(GSH)和植物螯合肽(PC)含量的测定 丙二醛含量的测定采用硫代巴比妥酸比色法测定,抗坏血酸含量的测定采用2,4-二硝基苯肼比色法。 采用三氯乙酸(DTPA)和5%磺基水杨酸顺时针快速研磨获得粗提物,GSH含量的测定参照韩丹的方法,即抽取0.25 mL上述提取物加入 2.6 mL 150 mmol/L NaHPO(pH值7.7)和 0.18 mL 5,5-二硫代双(2-硝基苯甲酸)(DTNB)试剂。采用30 ℃水浴振荡5min,采用紫外分光光度计在412 nm处测定吸光度,并根据标准曲线计算样品的GSH含量。PC含量测定:将1 mL粗提物与2.2 mL的0.2 mol/L的磷酸盐缓冲液(pH 值7.0)和0.3 mL的DTNB试剂混合,随后在30 ℃水浴振荡10 min,然后测量412 nm处的吸光度。 1.3.4 Cd转运蛋白基因表达水平测定 采用TRIzol试剂盒(Invitrogen)对小麦根系进行总RNA提取,使用DNaseI-Verso cDNA合成试剂盒将RNA反转构建cDNA。使用ReverTra Ace qPCR RT Kit从总RNA合成第一链cDNA。实时PCR采用SYBR Premix ExⅡ 与 7500快速实时PCR序列检测系统上进行。以为看家基因,相关Cd转运蛋白基因的引物序列见表1。热曲线程序:90 ℃30 s,95 ℃15 s,60 ℃30 s和72 ℃30 s,共40个循环。绘制熔解曲线进行分析和琼脂糖凝胶电泳以保证扩增子。具体反应体系、反应程序步骤见Zhou等的研究,采用2-ΔΔ断层扫描方法计算目标基因的相对转录丰度。 表1 Cd转运蛋白相关基因的qRT-PCR 引物序列信息 采用Microsoft excel 2013进行数据整理,采用IBM SPSS 19.0软件进行试验数据统计分析(=0.05),采用Origin 2021进行图形绘制。 由表2可知,株高和根长中,各处理均表现为Cd处理 表2 硒、硅对镉胁迫下小麦生长参数及叶绿素含量的影响 由图1-A可知,净光合速率指标中,以CK最高,Cd+Si+Se处理次之,二者无显著差异,均显著大于其他3个处理,且Cd、Cd+Si、Cd+Se这3个处理间差异不显著。由图1-B可知,胞间CO浓度指标中,以Cd处理最低,CK、Cd+Se处理、Cd+Si处理、Cd+Si+Se处理较Cd处理分别显著提高173.63%、104.74%、136.80%、153.82%。气孔导度指标中,CK处理最高,Cd处理、Cd+Se处理、Cd+Si处理、Cd+Si+Se处理较CK分别显著降低75.34%、64.38%、67.12%、57.53%,同时这4个处理处理间差异不显著(图1-C)。蒸腾速率指标中,各处理表现为Cd处理 由图2-A可知,镉胁迫下,各处理根系的Cd浓度为地上部浓度的7.03~45.08倍,表明镉胁迫下小麦根系是主要的Cd累积器官。地上部中,以Cd处理浓度最高,为239.61 μg/g,其他处理较其显著降低34.04%~69.69%;Cd+Si+Se处理的镉浓度最低,其他处理较其显著增加112.57%~229.95%。各处理根系镉浓度规律与地上部相反,Cd+Si+Se处理镉浓度最高,Cd、Cd+Si、Cd+Se处理较Cd+Si+Se处理显著降低48.55%、26.33%、53.94%,且Cd+Si处理的浓度显著高于Cd、Cd+Se处理。镉转移系数为地上部浓度与根系浓度之比,因此,在转移系数中各处理呈Cd+Si+Se处理 小麦地上部和根系的细胞被分成细胞壁(Fcw)、细胞质(Fo)和细胞器(Fs)3个亚细胞部分。对亚细胞镉浓度的分析(图3-A)表明,各处理地上部的亚细胞镉总浓度均低于地下部,尤其是Cd+Si+Se处理。地上部中,亚细胞镉总浓度以CK处理最高,Cd、Cd+Si、Cd+Si+Se处理较CK处理降低67.37%~88.73%;根系中,各处理亚细胞镉总浓度表现为Cd+Se处理 由图4-A可知,各处理地上部的丙二醛含量明显高于根系,与CK相比,Cd胁迫处理显著增加了MDA含量,其地上部、根系增幅分别为38.20%、28.43%,在此基础上添加Si、Se、Si+Se显著降低了小麦植株地上部和根系的MDA含量,地上部的降幅为64.44%、19.69%、56.85%;根系的降幅为20.74%、18.73%、38.13%。由图4-B可知,AsA含量指标中,与相应部位的CK相比,Cd处理地上部、根系的AsA含量分别显著降低34.66%、40.08%,而施用硒、硅单一和组合处理的地上部显著提高30.01%~45.68%,根系提高8.14%~61.59%。而在GSH含量指标中,各处理的GSH含量与AsA含量的规律趋于一致,即与CK相比,镉胁迫降低了地上部和根系的GSH含量,施用硒、硅单一及组合处理则增加了地上部和根系的GSH含量(图4-C)。由图4-D可知,PC含量指标中,与CK相比,Cd、Cd+Si、Cd+Se、Cd+Si+Se处理的地上部和根系分别显著降低18.18%~48.27%和65.27%~81.52%;无论地上部还是根系,均以Cd处理的PC含量最低,与Cd处理相比,Cd+Si、Cd+Se、Cd+Si+Se处理的地上部和根系分别显著提高33.89%、30.54%、58.16%和84.92%、81.16%、87.94%。 测定小麦中参与镉转运的基因(、、和)的相对表达量,以了解硒和硅缓解小麦耐镉的分子机制。由图5-A可知,与CK相比,Cd处理降低了水稻根系中、和的相对表达量,增加了的表达水平;与Cd处理相比,添加Si、Si+Se使的表达水平分别显著上调118.18%、122.73%。与Cd处理相比,单施Si、Se使的相对表达量分别降低了30.51%、52.54%,而Si+Se处理的相对表达量增加了83.62%,与单施Si、Se相比,Si+Se的表达水平分别显著增加了164.23% 和286.90%(图5-B)。此外,与Cd处理相比,Si、Se和Si+Se的添加均显著抑制了表达,其相对表达量降幅分别为55.17%、64.66%、32.76%(图5-C)。与其他基因的表达趋势不同,与Cd处理相比,Si、Se的添加均降低了的相对表达量,而Si+Se处理使该基因相对表达量显著增加了1.25倍(图5-D)。 土壤Cd浓度超标已成为影响农业安全发展的制约因子,Si、Se是植物生长发育的有益元素,大量研究表明,胁迫环境下Si、Se对作物生理的影响存在积极作用。本研究结果表明,镉胁迫下,植物株高、根长及干物质累积量受到显著影响,在此基础上添加Si、Se可有效缓解上述生长参数,同时在Si与Se结合处理(Cd+Si+Se)下得到进一步促进,其长势与CK处理大体相当。此外,各处理小麦叶片的净光合速率、胞间CO浓度、气孔导度、蒸腾速率和叶绿素含量整体表现为Cd处理 重金属的亚细胞分布是植物中最主要的重金属解毒机制之一,植物细胞壁是保护原生质体免受镉毒害的首道屏障,它可提供羟基或羧基等极性物质与重金属离子结合,细胞壁上的离子交换位点饱和后,进入细胞的重金属会被隔离到液泡中,从而降低游离重金属离子的活性,进一步降低镉对植物细胞器的干扰。本研究中,在Cd含量较高的根系中,Cd主要集中在细胞壁中,且与细胞器相比,Si处理的镉在细胞质中的分布比例更小。这与Zhang的研究结论趋于一致,即硅可以通过增加细胞壁中Cd的含量和比例来减少Cd在细胞质中的分布。本研究中,与相应部位的CK相比,添加硒对根系细胞器中的Cd浓度没有影响,但却增加了地上部细胞器的浓度和比例;因此,地上部细胞器沉淀重金属可能是硒减轻小麦根系镉毒性的主要机制之一。当Si和Se结合使用时,小麦根系Cd浓度最高,转移系数最低,且与其他处理差异显著(图2);此外,地上部、根系细胞质中的Cd浓度及比例进一步降低,地上部细胞器的Cd含量增加(图3)。这说明Si和Se的协同作用可以显著抑制Cd上移,且促进地上部细胞壁螯合Cd。 氧化还原失衡是镉胁迫的另一个重要毒性表现。丙二醛是胁迫下过氧化反应的产物,会对植物细胞膜造成严重破坏;抗坏血酸是缓解细胞高度氧化的重要还原物质,可中和过氧化物质;而谷胱甘肽和植物螯合肽含有硫醇基团(—SH),在重金属解毒的螯合和分离过程中起重要作用。本研究中,Cd处理增加了小麦中的MDA含量,这表明Cd存在氧化损伤,在此基础上Si、Se单一或结合使用均显著降低了地上部、根系MDA含量(图4-A),各处理AsA含量变化趋势与MDA相反,这意味着Si、Se结合使用可有效调控MDA与AsA的区室化中和作用以减轻Cd毒性。此外,Si、Se单一或结合使用皆增加了GSH和PC含量(图4-C、图4-D),GSH分泌增加可以直接螯合重金属,且较高的GSH含量会增加PC的产生,进而螯合细胞中的Cd形成PC-Cd复合物,这些复合物通过液泡膜ATP酶转运至液泡,从而降低细胞质中的Cd浓度。 金属转运基因转录水平的调控对于调控植物中重金属的积累和分布至关重要。在小麦植株中,位于根皮和内胚层末端的是吸收Cd的主要转运蛋白,参与细胞Cd植物内的吸收和Cd运输,于小麦根细胞的液泡膜,负责将Cd螯合到根液泡中,于细胞质膜上,介导Cd从根系向地上部的转运。本研究中,Cd处理降低了小麦根系的表达(图5-A),添加Si、Se后,的相对表达量增加,与根系中Cd浓度变化趋势一致(图2),同时Cd处理上调了的表达,添加Si、Se后,的相对表达量显著降低(图 5-C),这会导致较少的Cd转移到地上部,这与小麦地上部的Cd浓度趋势一致(图2)。值得注意的是,Si+Se处理后,的相对表达量较单一Si、Se处理发生显著上调,而地上部Cd浓度和转运系数显著降低。此外,Cd处理抑制了和的表达,在此基础上Si+Se处理的表达水平显著上调(图5-B、图5-D),这与根中Cd亚细胞分布一致(图4)。这些发现进一步表明,Si和Se的协同作用可以通过调节和的表达来促进Cd向液泡的转运,从而降低根系Cd毒性。1.2 试验设计
1.3 样品采集及测定分析

1.4 数据处理与统计分析
2 结果与分析
2.1 硒、硅对镉胁迫下小麦生长及叶绿素含量的影响

2.2 硒、硅对镉胁迫下小麦光合特征参数的影响
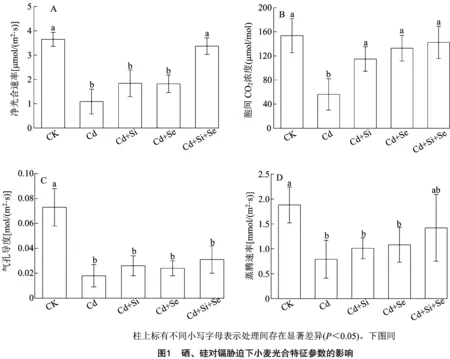
2.3 硒、硅对镉胁迫下小麦镉吸收及转移的影响

2.4 硒、硅对镉胁迫下小麦镉亚细胞分布的影响
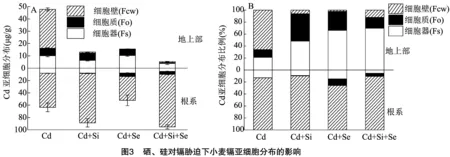
2.5 硒、硅对镉胁迫下小麦抗氧化系统指标的影响
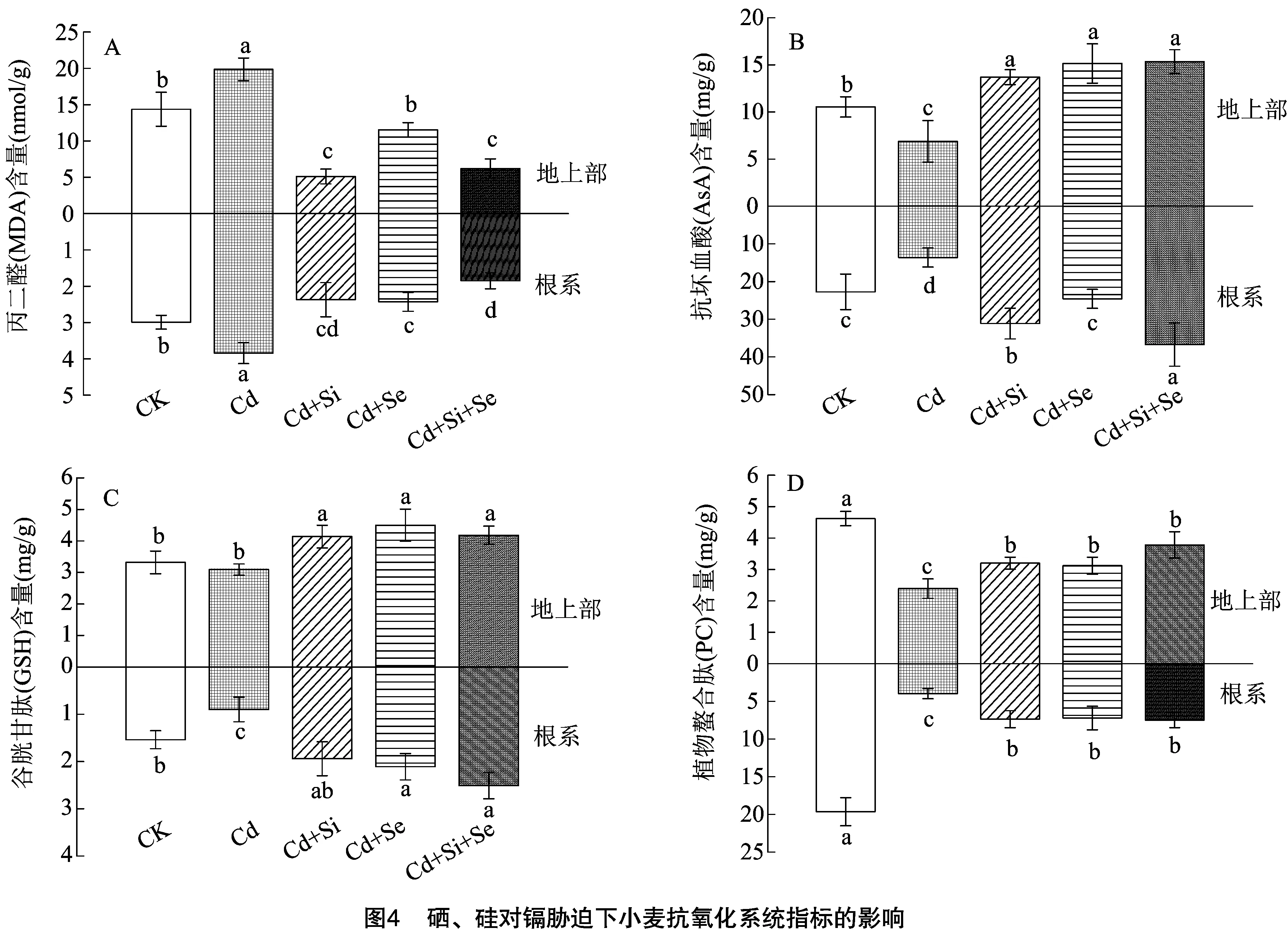
2.6 硒、硅对镉胁迫下小麦镉转运基因表达的影响
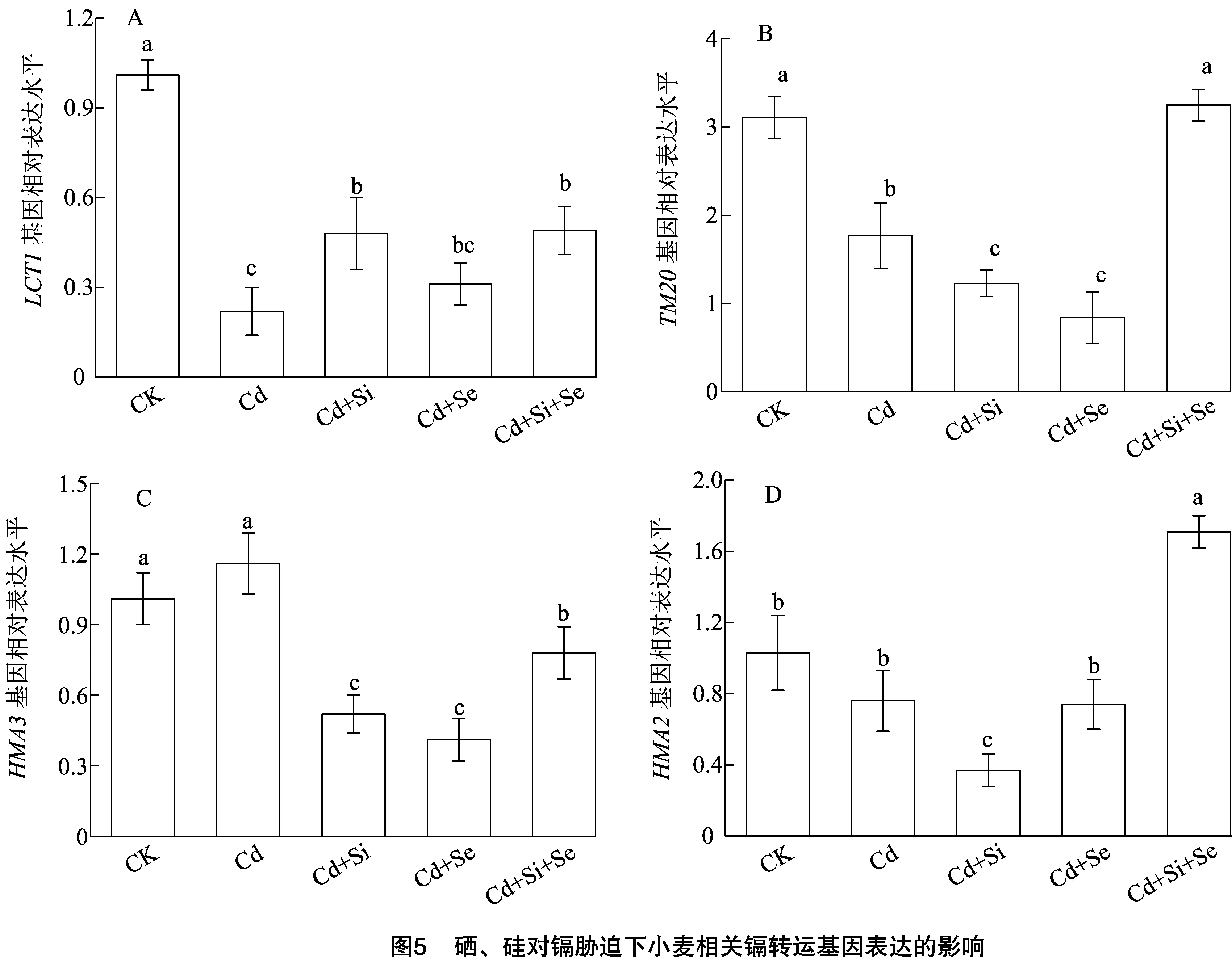
3 结论与讨论
猜你喜欢
四川劳动保障(2021年3期)2021-06-09矿产综合利用(2020年1期)2020-07-24中成药(2018年8期)2018-08-29浙江工业大学学报(2017年5期)2018-01-22现代园艺(2017年13期)2018-01-19广东农业科学(2017年5期)2017-08-29西南农业学报(2016年5期)2016-05-17中国农业文摘-农业工程(2016年5期)2016-04-12当代化工研究(2016年6期)2016-03-20中国资源综合利用(2016年3期)2016-01-22
