
国外梢斑螟属害虫研究进展
2022-09-22 01:29:54聂振业宋丽文迟德富
中国森林病虫 2022年5期
聂振业宋丽文迟德富
(1.东北林业大学,黑龙江哈尔滨 150040;2.吉林省林业科学研究院,吉林长春 130117)
梢斑螟属Dioryctria隶属于鳞翅目Lepidoptera螟蛾科Pyralidae螟蛾亚科Phycitinae,是全球重要的针叶树害虫,以幼虫取食嫩芽、针叶、形成层和球果,生活隐蔽,生活史主要在枝梢与球果内完成,不易人工饲养与观察[1]。梢斑螟的侵染不仅对针叶林树种更新构成巨大威胁,也对种子园的培育造成了严重危害。多年来,国内外学者对梢斑螟属种、种团划分开展了大量基础性研究工作,也对具有一定危害的梢斑螟种类进行了生物学、生态学、发生规律及研究方法探索,取得了许多重要的研究进展。近年来,梢斑螟属害虫在我国大面积发生,严重危害红松Pinus koraiensis、樟子松Pinus sylvestrisvar.mongolica等针叶树,给资源、经济造成重大损失。国内一些学者对我国梢斑螟属害虫的种类鉴别、生物学特性、发生危害特点和防治技术进行了系统总结[2-5],但该类群国际上研究进展的总结未见报道,笔者对国际上梢斑螟属类群不同种团的主要种类、危害特点、生物学特性和防治方法加以总结,以提供研究借鉴。
1 国外梢斑螟属重要种类概述
螟蛾科为鳞翅目中的一个大科,全世界记载1万多种,大部分为农林业的重要害虫。其中螟蛾亚科为螟蛾科中的第二大亚科,该亚科的昆虫个体通常较小,并且种间形态特征十分相近,常会造成鉴别错误与种名混淆。目前全球已发现的梢斑螟属的种类约78~86个种,归属于11个种团(其中1个为中国特有种团)。
由于梢斑螟成虫鉴定特征有重叠,如翅颜色、生殖结构以及寄主选择关系,单纯依靠形态特征及外生殖器特征鉴定梢斑螟属昆虫十分困难[2,6]。但是形态特征分类依然是梢斑螟划分种团、确定物种的最重要手段,而分子生物学技术也成为了该属昆虫相似物种鉴定、种团辅助划分、系统发育研究等的重要技术。如在NCBI网站(https://www.ncbi.nlm.nih.gov/)上分别有梢斑螟线粒体细胞色素氧化酶I(COI)基因序列336条和II(COII)6条,分别涵盖33种和24种。通过序列分析,对团的划分以及种团之间的系统发育关系有了更深入的理解[3-4,7-8]。
在11个梢斑螟种团中,有9个种团广泛分布在北半球亚热带地区至亚北极区,在新热带区和印度马来西亚区(Indo-Malaysian)也有少数物种。其中1个种团(Pygmaeella group)不以松树为食。在国外危害严重的有6个种团,危害较轻的有Erythropasa种团、Ponderosae种团、Taiella种团和Pygmaeella种团。这些种团中有部分种类以树木韧皮部、针叶等为食,但对针叶树生长发育不会造成重大影响。因此,关于这些危害较轻的种团的生物学、生态学特性、危害及防治技术的研究较少。
在我国有记录的15种梢斑螟,分别属于5个种团,即冷杉梢斑螟种团(Abietella group)、松梢斑螟种团(Auranticella group)、云杉梢斑螟种团(Schuetzeella group)、赤松梢斑螟种团(Sylvestrella group)和樟子松梢斑螟种团(Mongolicella group)。其中,樟子松梢斑螟种团为我国特有种团,且只记录了樟子松梢斑螟D.mongolicella1个物种[2]。
本文综合除我国特有种团(樟子松梢斑螟种团)以外的10个种团,对相关资料记载的54种梢斑螟(含同时分布在我国的11种)的地理分布、危害部位进行了总结,并着重对危害相对较重、研究较多的6个梢斑螟种团的分布和危害等进行分析。
因为樟子松梢斑螟、云南松梢斑螟D.yuennanella、昆明松梢斑螟D.kunmingnella和梵净梢斑螟D.fanjingshana等4个种,仅记载在我国有分布,国外几乎没有研究,因此未在本文中记述。
1.1 冷杉梢斑螟种团
该种团有10种梢斑螟,其中多数分布于新北区或古北区,在北美洲和中美洲、欧洲、北非和亚洲均有分布(表1)。寄主植物为针叶树,主要以幼虫寄生枝梢、取食球果危害,大部分为重要的经济害虫。
该种团在国外危害较重的2个种为云杉球果螟D.abietivorella和D.ebeli。云杉球果螟分布范围较广,横跨加拿大南部和整个美国西部,而D.ebeli分布仅限于美国东南部;云杉球果螟寄主植物范围与侵害部位均较广,对寄主有较强的适应性[1,9-12]。

表1 冷杉梢斑螟种团的种类、寄主植物、侵害部位及分布范围Tab.1 Species,host plants,infestation parts and distribution of Abietella group abroad
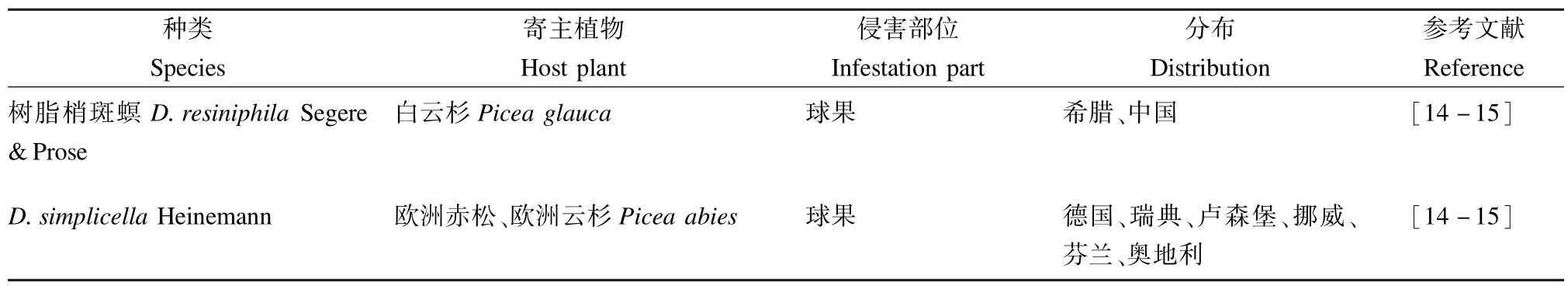
续表1Tab.1(Continued)
1.2 松梢斑螟种团
该种团中D.auranticella、D.rossi和D.disclusa均为重要的松树果实害虫(表2)。
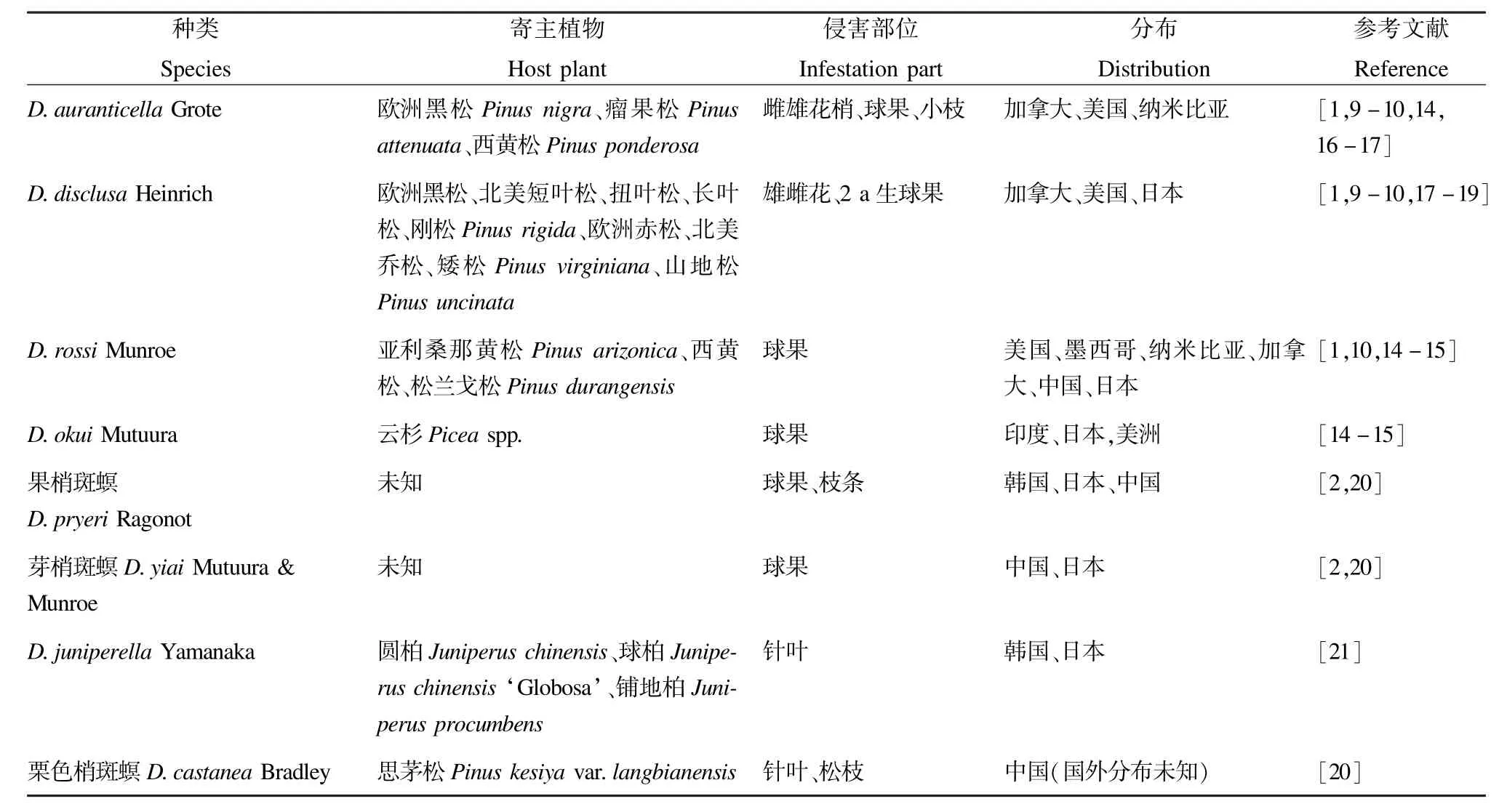
表2 松梢斑螟种团的种类、寄主植物、侵害部位及分布范围Tab.2 Species,host plants,infestation parts and distribution of Auranticella group abroad
在加拿大不列颠哥伦比亚省南部和美国新墨西哥州之间的大部分范围内,D.auranticella和D.rossi共域分布[9-10]。该种团中的D.disclusa、D.rossi、D.okui、果梢斑螟D.pryeri、芽梢斑螟D.yiai和D.juniperella均在亚洲有分布(表2)。
1.3 Baumhoferi group
该种团均分布于美国和加拿大。寄生部位以松树球果与嫩枝为主,该种团中只有2种侵染球果,是松疱球果梢斑螟D.clarioralis和D.pentictonella。松疱球果梢斑螟分布在美国东南部,是美国南部种子园中一种重要的经济害虫,而D.pentictonella分布在西部,未在种子园中造成严重危害(表3)。
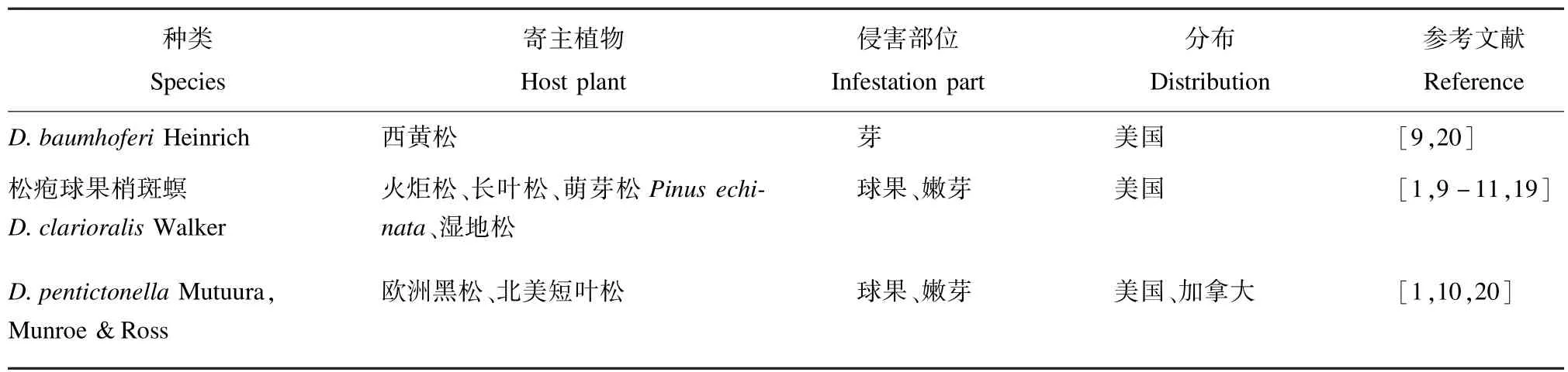
表3 Baumhoferi种团的种类、寄主植物、侵害部位及分布范围Tab.3 Species,host plants,infestation parts and distribution of Baumhoferi group abroad
1.4 云杉梢斑螟种团
该种团记载有3个种,其中针枞梢斑螟D.reniculelloides以及伪节梢斑螟D.pseudotsugella寄生在各种针叶树上,但很少在松树上[22]。针枞梢斑螟广泛分布于加拿大南部和美国北部,在加拿大不列颠哥伦比亚省和美国西北部有记录;伪节梢斑螟的分布范围相对于针枞梢斑螟较窄。Roe和Sperling对这两个物种形态、地理分布等进行了诊断和细致划分[23]。云杉梢斑螟D.schuetzeella在欧洲主要寄生在杉科植物上(表4)。

表4 云杉梢斑螟种团的种类、寄主植物、侵害部位及分布范围Tab.4 Species,host plants,infestation parts and distribution of Schuetzeella group abroad
1.5 赤松梢斑螟种团(Sylvestrella group)
该种团分布于欧洲和亚洲。幼虫主要以松属植物枝梢和球果为食,有时也生长在其他针叶树上[12],如赤松梢斑螟D.sylvestrella在云杉属Picea和冷杉属Abies植物上均有发现(表5)。
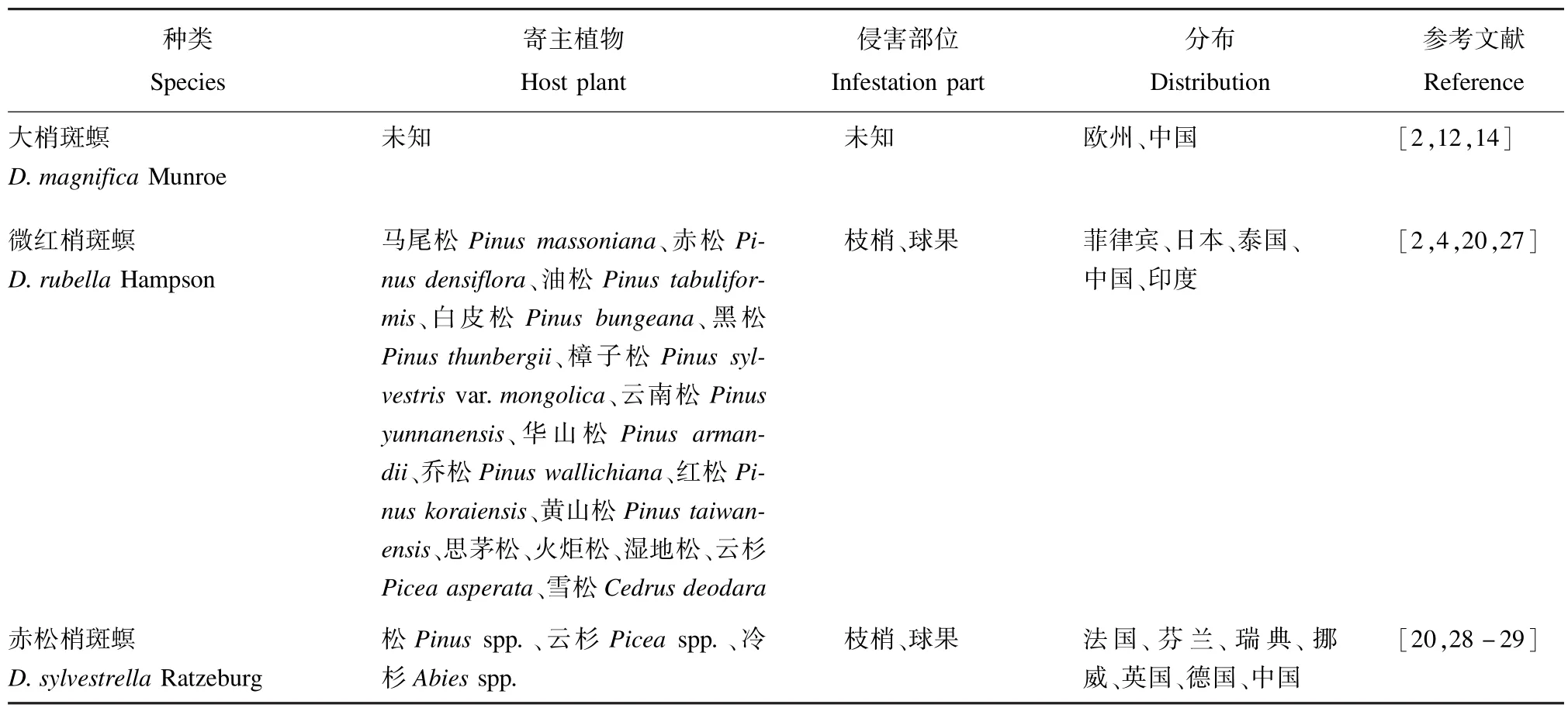
表5 赤松梢斑螟种团的种类、寄主植物、侵害部位及分布范围Tab.5 Species,host plants,infestation parts and distribution of Sylvestrella group abroad
1.6 Zimmermani group
该种团包含18种,其中白带梢斑螟D.albovittella、南方松梢斑螟D.amatella、D.cambiicola、火炬松球果螟D.merkeli、D.resinosella、D.taedae、小火炬松梢斑螟D.taedivorella以及D.yatesi等8种侵染球果(表6)。这些种中,危害最严重的是南方松梢斑螟和火炬松球果螟。与冷杉梢斑螟种团一样,该种团由于分类特征不明显以及和幼虫寄主的重叠,Zimmermani种团内的分类比较混乱。如D.taedae是火炬松的一种重要害虫,分布于美国特拉华州到乔治亚州,该种通常与形态相似的南方松梢斑螟和火炬松球果螟物种混淆[1]。南方松梢斑螟分布于美国东南部,在部分范围内与D.taedae同域。虽然南方松梢斑螟被认为是长叶松P.palustris的主要害虫,但已知该区域的其他梢斑螟也侵染长叶松,因此不能利用幼虫寄主作为该区域梢斑螟种类的区别特征[1,10]。小火炬松梢斑螟分布于美国弗吉尼亚州和北卡罗莱纳州到阿拉巴马州北部和密西西比州[10,30],危害火炬松P.taeda。该物种曾与火炬松球果螟、美洲松梢斑螟D.zimmermani混淆,尽管后者现在被发现是寄主植物形成层组织的蛀虫[10]。火炬松球果螟在形态上与该生活范围内的许多其他物种相似,其寄主是佛罗里达州北部、乔治亚州南部和密西西比州南部的火炬松、湿地松P.elliottii和长叶松。D.yatesi仅限于美国东南部沿海和美国田纳西州的山区,是Zimmermani种团中唯一侵染刺松P.pungens的种类。D.resinosella以多脂松P.resinosa为食,遍布加拿大安大略省南部和美国东北部,从美国缅因州到明尼苏达州。D.cambiicola是北美洲西部Zimmermani种团中已知的唯一危害球果的物种。Mutuura等在1969年报道,在加拿大不列颠哥伦比亚省和美国西部的发现D.cambiicola,只在大果松P.coulteri、扭叶松P.contorta上记录到了该种[24]。
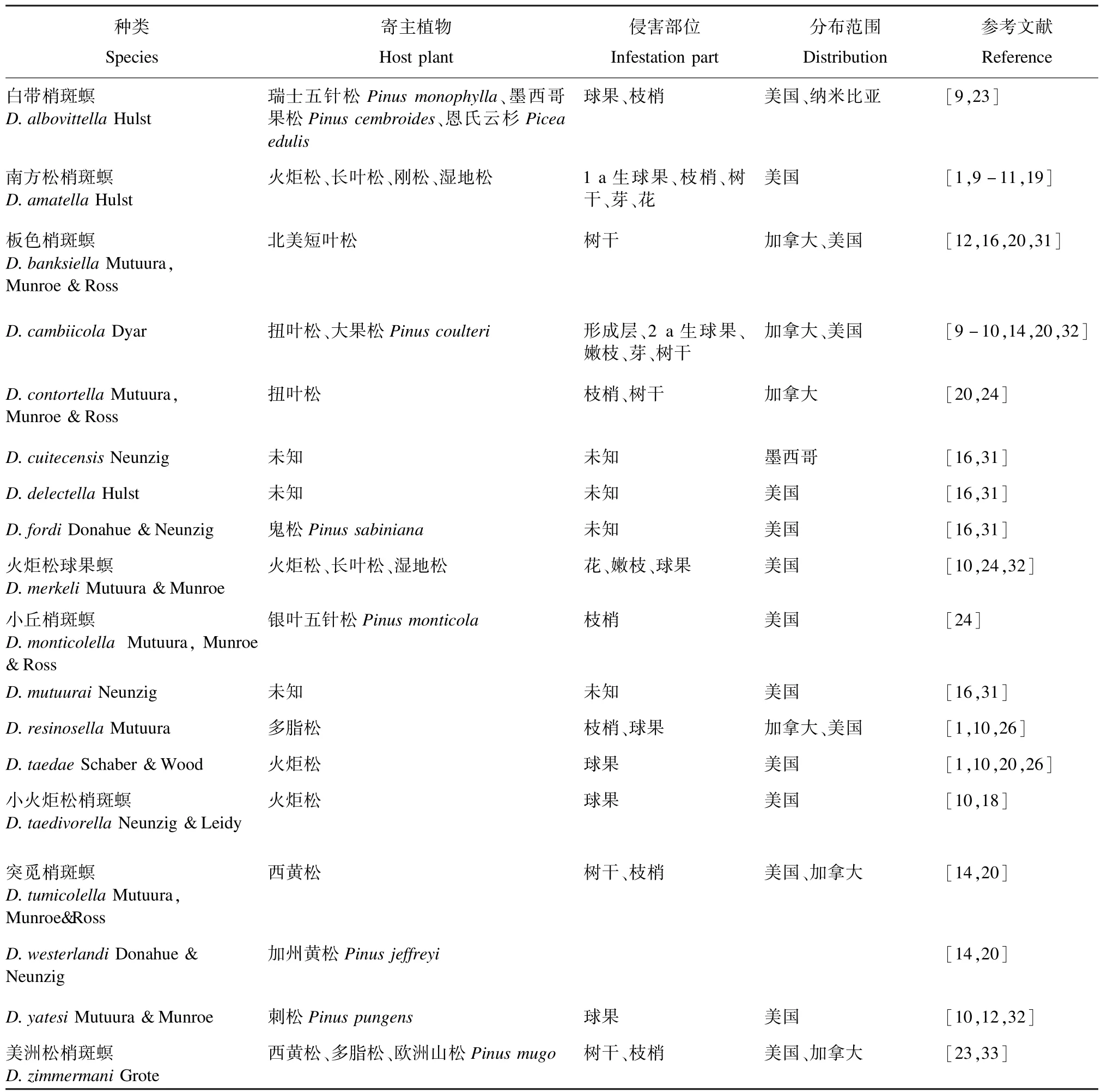
表6 Zimmermani种团的种类、寄主植物、侵害部位及分布范围Tab.6 Species,host plants,infestation parts and distribution of Zimmermani group abroad
在国外危害较轻的4个种团的部分梢斑螟种类名称、寄主、危害部位及分布见表7。
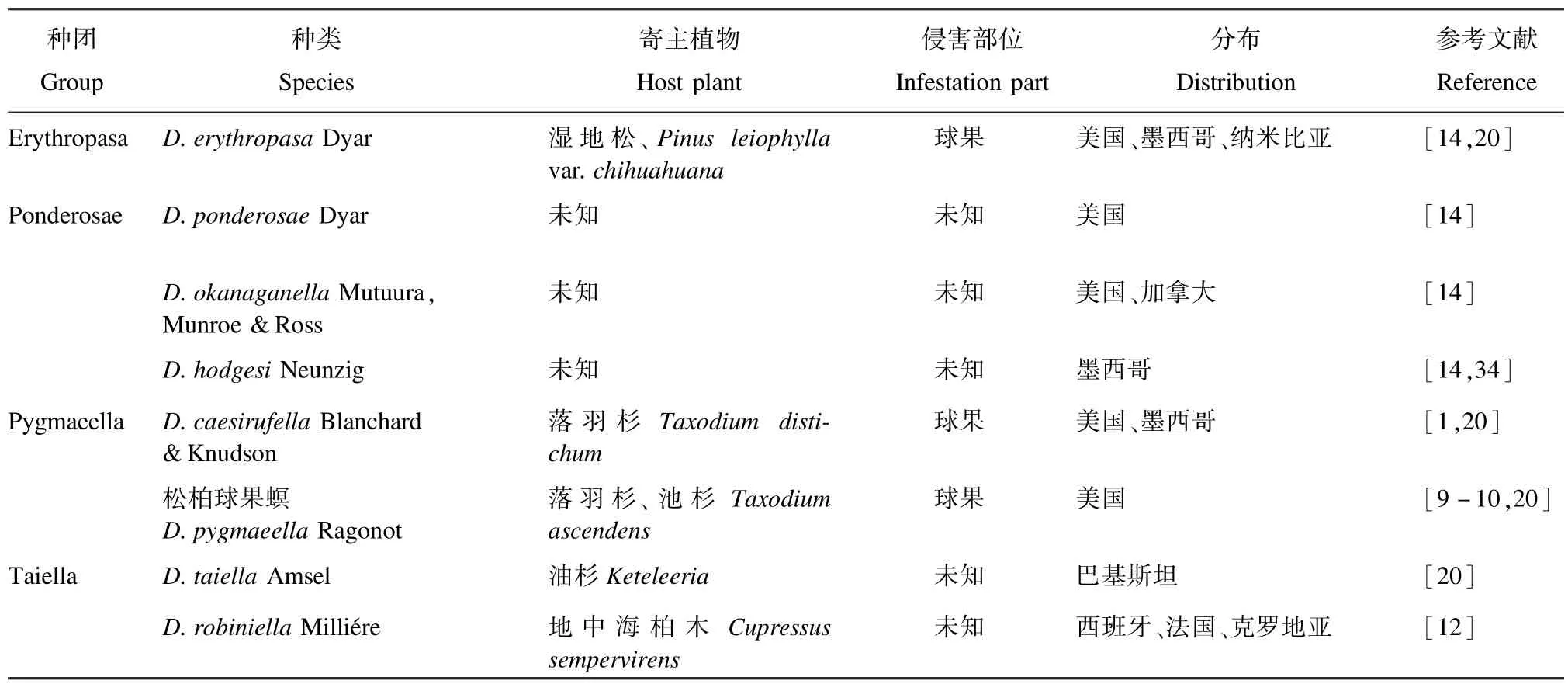
表7 国外梢斑螟危害较轻的种类、寄主植物、侵害部位及分布范围Tab.7 Species,host plants,infestation parts and distribution of harm slightly of Dioryctria pests abroad
2 主要生物学和生态学特点
2.1 生物学要点
1)有效积温影响发育进度和世代数。在(26±2)℃下,D.auranticella卵的孵化期为6~9 d,23 ℃时孵化期约为15 d[17]。不同地点的世代数也不尽相同,尤其是在温暖地区,常会出现世代重叠现象,如松疱球果梢斑螟D.clarioralis通常是1 a 1代[18],但在美国南部可能是1 a多世代[35]。
2)幼虫常利用已有虫道转移危害。梢斑螟常通过其他球果害虫的虫道侵入寄主,造成二次伤害,从而扩大损失[1,9,14,36]。 在一些梢斑螟中,当取食部位资源耗尽时,幼虫会在球果间迁移,从而危害多个球果[9,14]。
3)越冬和化蛹场所多样。大多数梢斑螟均在其寄主危害部位(或附近)化蛹,但也有部分梢斑螟在寄主附近土壤中化蛹,如D.auranticella在球果内化蛹,而冷杉梢斑螟D.abietella在土壤中越冬、化蛹[1,9-10]。梢斑螟的多样越冬和化蛹习性值得关注。
4)成虫多具有趋光性。梢斑螟成虫像许多螟蛾科昆虫一样,具有夜行与紫外光敏感特征[37-39]。Fatzinger等发现天黑后3 h内冷杉梢斑螟蛹的羽化速度最快,并于夜间交配行为增加[14,37]。Yates等在美国乔治亚州采用黑光灯确定了4种梢斑螟的飞行物候[35]。
5)雌雄成虫羽化时间差异。雌雄成虫羽化期具有差异,如D.auranticella[17]和D.disclusa[14]的雄性比雌性分别早出现2~3 d和5 d。这些信息可以帮助我们更好认识种内互作规律。
6)求偶和交尾多在夜间进行。对梢斑螟的繁殖行为已有一些较为详尽的描述。梢斑螟属昆虫大部分以性信息素为介导,以雌性召唤(calling)来吸引雄性。Fatzinger等详细描述了D.ebeli(冷杉梢斑螟种团)和南方松梢斑螟D.amtella(Zimmermani种团)的交尾行为在召唤行为与交尾频率上存在着显著的种内差异[37,40],D.ebeli雌虫以 12L∶12D 人工光周期饲养,在天黑后5~6 h后开始召唤,9 h后达到召唤峰值,并在光照开始前停止;雌性会采取一种特有的召唤姿势,腹部尖端在翅间向上弯曲,从产卵器顶端挤出信息素;在羽化后的第1天,大约30%的D.ebeli会召唤并接受交尾[37]。在人工黑夜期,召唤雌性数量翻倍,D.disclusa在天黑后3~5 h开始召唤,雄蛾会追随性信息素气味寻找雌蛾[41]。
7)雌虫常与多雄交尾。Auranticella group、Pounderosae group、Zimmermani group和冷杉梢斑螟种团等4个种团中的雌性,均具有与多雄交尾现象。其中云杉球果螟个体通常交尾8次,其他物种交尾2~3次;但目前尚未能确定交尾对雌虫的健康及再次交尾的影响,对雌雄后续交尾的影响可能因种团或物种而异,因此交尾频率因物种而异[41-42]。
2.2 植物源信息化合物影响成虫行为和产卵选择
Shu等发现从北美乔松P.strobus松枝上释放的单萜烯-香月桂烯和3-蒈烯,会引起云杉球果螟雌性成虫的触角和产卵反应,虽然(2)-柠檬烯是另一种促卵剂,但它不会引起触角的显著电生理反应;与未交尾的梢斑螟相比,这些单萜引起交尾后的雌性成虫的触角反应更强烈;成虫对于不同位置的挥发物具有不同的反应,如Asher等发现D.ebeli雄虫和雌虫对从湿地松第1年球果基部释放挥发物的触角反应大于对第2年球果或第1年球果的顶端和中部释放挥发物的触角反应,这表明球果基部位置可能是一个重要的产卵地点[40,43]。
不同梢斑螟属昆虫产卵地点与寄主单萜排放量似乎具有相关性。Fatzinger等发现,在实验室中南方松梢斑螟(Zimmermani种团)雌虫仅在柱锈菌Cronartium quercuumf.sp.fusiforme危害诱导的虫瘿(fusiform rust galls)情况下才开始召唤、交尾和产卵[40]。这些瘿瘤是柱锈菌的锈孢子产生阶段,诱导松树产生的肿瘤状突起,被诱导部位产生大量的α-蒎烯为其主要促卵剂;尽管1龄幼虫取食量与单萜含量呈负相关,但雌虫更喜欢在单萜含量较高的地方产卵[40]。
2.3 性信息素及其对行为的影响
目前已知鉴定出了11种梢斑螟属昆虫的性信息素(表8)。其中7种[44-45]以Z9-tetradecenyl acetate(Z9-14:Ac)为其主要成分,在云杉球果螟等种类的性信息素中也均有一定含量,但不是含量最高的组分;(Z9,E11)-tetradecadienyl acetate(Z9,E11-14:Ac) 和(Z3,Z6,Z9,Z12,Z15)-pentacosapentaene(C25 pentaene)这两种成分在梢斑螟性信息素中也比较常见;在触角电位图分析中,C25 pentaene只能微弱地刺激雄蛾触角,但在北美洲西部的野外试验中发现,添加该成分对于吸引雄蛾至关重要。此外,研究发现雌蛾释放的各信息素成分的比例,与在野外有效诱捕雄蛾时添加到诱芯中的各成分比例有很大不同;这种现象是否受到了诱芯中各种成分释放速率的影响,以及雄虫如何区分这种微量的差别尚需深入探究;在不同地区的同种雄性梢斑螟对不同性信息素成分比例的吸引力具有差异,合成的Z9,E11-14:Ac与C25 pentaene的比例为1∶10时,会吸引雄性云杉球果螟,但云杉球果螟雄性对雌性性信息素的反应在地理上有所不同[46-47]。由于各成分具有不同的结构类型,它们很可能是通过独立的生物途径产生的[47]。

表8 几种梢斑螟的性信息素主要成分Tab.8 Main components of sex pheromone of several Dioryctria

续表8Tab.8(Continued)
3 监测与防治方法
梢斑螟属害虫部分物种对全球范围内的松科植物造成了巨大危害,由于其生活史不规律并且幼虫生活较隐蔽,在被害处常会有保护性泌脂反应,因此选择正确的防治措施是十分必要的。笔者仅从人工物理、化学防治与生物防治等几个方面,对在国外危害较重、防治效果良好的事例进行简要总结,以期为我国梢斑螟属害虫防治提供借鉴。
关于性信息素诱捕监测,目前国外鉴定出了11种梢斑螟性信息素组分及比例,而且探索出了针对8种主要梢斑螟的野外诱捕配方。Strong等通过在树冠不同位置放置诱捕器,发现树顶附近可以捕获更多的云杉球果螟成虫[47]。单一诱捕器的诱捕效果高于多种诱芯诱捕器一起使用的效果[42]。
灯光诱捕可以确定成虫羽化进度、雌雄成虫比例,并预测卵或低龄幼虫的发育进度。Mcleod等在美国阿肯色州使用黑光灯诱捕器,监测了5种梢斑螟,发现诱捕到的南方松梢斑螟成虫数量波动规律,与羽化进度一致,证明羽化后的成虫具有良好的趋光性[26]。Roe等使用黑光灯诱捕器在加州收集了云杉梢斑螟种团的成虫[31],除松柏球果螟D.pygmaeella(Pygmaeella种团)外,多数种类在光诱结果中,捕获量雄性超过雌性[26],为正确分析灯诱数据提供了参考。
3.1 人工物理防治
采用黑光灯诱捕的方法,可以诱捕冷杉梢斑螟种团的云杉球果螟成虫,诱捕到的雄成虫数多于雌成虫;亦可诱捕松梢斑螟种团的D.auranticella[56]和云杉梢斑螟种团的D.pseudotsugella,以及Zimmermani种团的南方松梢斑螟成虫[56]。
3.2 化学防治
国外主要报道了用化学防治方法控制冷杉梢斑螟种团的种类,如在喜马拉雅冷杉Abies pindrow胸径处,以100 mL/20 cm或100 g/20 cm周长的剂量,采用内吸性噻虫胺滴灌法或林木注射剂乐果等药剂后,冷杉梢斑螟D.abietella的危害率可降低到16.66% ~33.33%[57]。 在印度部分地区通过对云杉树干根部注射硫脲、呋喃和乐果,优于在土壤中施用颗粒剂,对胸径20 cm的云杉防治冷杉梢斑螟最优效果可达86.49%[58]。
3.3 生物防治
对冷杉梢斑螟种团,性信息素诱捕松果梢斑螟D.mendacella成虫时,(3Z,6Z,9Z,12Z,15Z)-pentacosapentaene与(9Z,11E)-tetradecadienyl acetate的比例至少为10∶1,具有较强吸引力;性信息素干扰交尾方法,可以有效降低冷杉梢斑螟的危害,如性信息素干扰后,采用黑光灯诱捕,诱捕量降低94% ~100%[59]。山地松P.uncinata球果喷施欧洲五针松P.cembra球果挥发物,可显著降低棘梢斑螟D.mutatella对球果侵害[60]。
通过在挪威云杉P.abies上喷施苏云金芽孢杆菌kurstaki×aizawai品系,冷杉梢斑螟危害从30%降低到15%[61]。狭面姬小蜂属的Elachertus spinola等7种姬小峰在云杉球果螟幼虫中存有寄生[14,24]。
对Zimmermani种团,性信息素诱捕南方松梢斑螟、火炬松球果螟D.merkeli成虫。室内生测发现南方松梢斑螟对于苏云金杆菌高度敏感,苏云金杆菌悬液对南方松梢斑螟2龄幼虫的 LC50为42.3 IU/mL[23]。用信息素诱捕器监测到南方松梢斑螟成虫7 d后,施用苏云金杆菌柏林亚种,球果保存率增加约18%,结籽量增加48%[24]。从美国火炬松种子园中被侵染的南方松梢斑螟幼虫上,采集到了3株白僵菌,室内生测发现其致死率均达到60%以上[61]。在火炬松种子园中,南方松梢斑螟的膜翅目与双翅目天敌的寄生率达18.8%[60]。
4 展望
4.1 加强梢斑螟属害虫的物种鉴定与生物生态学研究
梢斑螟属害虫形态差异较小,部分种团的种类分布与寄主重叠现象比较严重,目前国外已将危害严重的梢斑螟种团的物种,进行了比较细致的鉴定和种团划分。借鉴国外的研究进展,需要注意近源种鉴定、系统发育和生物生态学研究。对物种准确鉴定及对其生物生态学习性的全面了解,是高效安全防治的基础。
1)应通过分析形态特征(如成虫翅图案颜色、生殖器官差异)、物种的分子生物学信息(如线粒体COI基因序列标记)以及生物生态学行为特征(幼虫与植物间危害关系)来综合描述和划分梢斑螟属种团、物种和种群。梢斑螟属许多种类均分布较广,且常有几种混合发生的情况。由于不同分布区的气候与寄主差异,造成物种的种群分化明显、形态差别明显、生物学特性差距较大。如在不同分布区冷杉梢斑螟外部形态、寄主、侵害部位与世代均呈现明显不同[8,14,37]。 单靠形态来鉴定种类,对多数研究者和生产单位技术人员都很难实现,采集的标本都送由从事梢斑螟分类的专业人员鉴定也不现实,这也是目前文献中梢斑螟种类及相关生物学习性记载较乱的原因之一。如能采用形态、分子生物学及生物生态学特征综合方法,建立某区域梢斑螟种类的鉴定数据库、手册、指南,可加快鉴定进程、提升鉴定的准确性。另外,应考虑由地理因素引起的物种种下分化及与寄主植物的关系。最好在一种害虫的分布区内,对其进行彻底的遗传调查,量化种内变异。例如冷杉梢斑螟种团中,冷杉梢斑螟种内变异大,不形成一个单系群,而该种团的其它种类种内变异较小;种内变异大小会直接影响物种对环境、寄主等的适应性;在区分物种间差异时,更有必要借助分子生物学技术[8,14,37]。
2)对于不同种团梢斑螟性信息素的研究还需进一步深入。国外针对性信息素研究已较为深入,特别是对分布在北美洲东部和西部的云杉球果螟雌虫释放的性信息素组分的不同比例,以及针对能有效吸引东、西部种群雄虫的有效比例的探究给我们较大的启发。应进一步分析我国不同分布区雌虫性信息素主要成分配比对成虫生理过程的影响,如交配中断或者引诱,而不仅是通过引进国外性信息素产品进行尝试。因梢斑螟属昆虫对性信息素微量成分有较强的感知机制,所以在我国进行性信息素研究时更应对微量成分进行关注,这对更好监测与防治梢斑螟属害虫具有重大意义。
4.2 深入探索高效规范监测技术
国内开展了一些梢斑螟种类的灯诱监测研究,但由于同一地区多种梢斑螟混合发生,且成虫体小、近源种间外观差异不明显等原因,尚没能建立起规范灯诱、准确鉴定、监测该类害虫种群的技术体系。
我国对梢斑螟属害虫性信息素研发较少,目前仅鉴定了微红梢斑螟和樟子松梢斑螟2种的性信息素成分和释放比例。但对这2种梢斑螟的性信息素最适诱捕组分比例、诱捕效果等都需进一步深入研究。可以认为,我国尚未实现利用性信息素对主要梢斑螟种类的高效精准监测,应尽快建立主要梢斑螟的监测技术体系和标准。成功防治梢斑螟属害虫依赖于完善的监测方案、适当的控制策略和高效安全的控制技术。监测预报能准确地掌握当地梢斑螟的发生期及发生量,在此基础上制定出合理的防治方案。梢斑螟属害虫的监测应该包括取样解剖观察、灯光诱捕和信息素诱捕监测等环节。其中,取样解剖观察可以通过观察被害状、排粪孔、虫粪、流脂、幼虫(或蛹)特征、幼虫(或蛹)数量等准确检测害虫种类、虫口密度和发育进度等。但目前国内外均未制定针对主要梢斑螟种类监测的规范与技术标准。
4.3 加强营林和生物防治研究工作
营林措施不仅影响寄主林木的质量,也对目标害虫种群具有重要影响。到目前为止,国内外均未能充分揭示梢斑螟成灾与林分组成、结构、多样性的关系。但是已经发现樟子松梢斑螟、赤松梢斑螟、D.contortella喜欢选择旧坑道、树木伤口、病斑部位侵入寄主[12,23-24]。 如 Merkel等[51]发现,在受病害的球果中D.ebeli幼虫数量较无病害的球果中更多。在球果因虫和病联合作用而衰弱死亡后,梢斑螟会转移到其他球果中,此时虫体携带的病原菌也会随幼虫转移到新球果中,使侵染加剧。当病害得到控制后,球果虫害率也相应地降低。因此,针对主要梢斑螟种类,应探明其暴发成灾与林分状况等环境关系,并建立有效的营林控制技术体系。
生物防治是可持续控制力强、环境友好的防治手段。值得借鉴的是国外在梢斑螟防控中采用苏云金杆菌取得了良好的防治效果。在实验室情况下,球孢白僵菌和绿僵菌对幼虫具有毒性,这2种真菌有很大潜力开发成生物农药。国外研究发现,梢斑螟属的寄生性天敌很多,包括茧蜂科Braconidae、姬蜂科Ichneumonidae和赤眼蜂科Trichogrammatidae以及蝇科Muscidae,寄生率达18% ~50%[61],但关于寄生蜂(蝇)的生物学、生活史、饲养防治技术的研究较少,在自然状态下对于梢斑螟属的种群控制效果评价也相对较少。尽管我国发现了梢斑螟啮小峰Baryscapus dioryctriae,并实现了规模饲养和释放,也开展过一些利用蒲螨Pyemotes和昆虫病原微生物防控梢斑螟的研究,但尚未实现对梢斑螟属害虫的高效防控。因此,有必要加强梢斑螟寄生和捕食性天敌的挖掘,天敌规模化繁育、释放、效果评价技术的深入研究。同时,从虫尸及其生活环境中分离鉴定梢斑螟虫生真菌、细菌或病毒等病原微生物,加强高致病力病原微生物的开发及应用技术研究。
综上所述,为更加有效地控制梢斑螟属害虫,应首先准确鉴定并确定关注区域内的梢斑螟种类,通过监测手段深入了解其生活史,充分利用其生物学特性中的薄弱环节加以调控。通过营林措施进一步增加林分自然控制能力和降低寄主的易感性,进一步探索利用白僵菌、苏云金杆菌、寄生及捕食性天敌等生物防治技术,适时精准地开展化学调控,全面提高我国安全高效可持续防控梢斑螟属害虫的能力。
猜你喜欢
现代农村科技(2020年7期)2020-12-19 03:12:08
广西林业科学(2019年2期)2019-07-25 02:28:46
中国林副特产(2019年3期)2019-01-10 17:17:11
福建林业(2018年3期)2018-11-07 05:34:36
收藏界(2018年1期)2018-10-10 05:23:08
创新作文(小学版)(2018年31期)2018-05-16 03:00:30
启迪与智慧·教育版(2018年12期)2018-02-26 12:24:06
山西林业科技(2017年4期)2018-01-23 05:45:05
摄影之友(影像视觉)(2017年1期)2017-07-18 11:12:16
甘肃林业(2016年2期)2016-11-07 08:56:12
