
硇洲岛大型海藻场软体动物群落的季节演替及其与环境因子的关系*
2022-09-21 02:37:12孙省利张才学廖海晴彭宇航
海洋与湖沼 2022年5期
姚 昕 孙省利, 2 张才学, 2 廖海晴 彭宇航
硇洲岛大型海藻场软体动物群落的季节演替及其与环境因子的关系*
姚 昕1孙省利1, 2张才学1, 2①廖海晴1彭宇航1
(1. 广东海洋大学化学与环境学院 广东湛江 524088; 2. 广东海洋大学海洋资源与环境监测中心 广东湛江 524088)
广东湛江硇洲岛是典型的潮间带生态系统, 属于热带气候, 适宜底栖生物生存。底栖动物作为生态指标的一部分, 可以反映出地理环境的多样性。于2011~2012年在硇洲岛潮间带五个位点进行底栖软体动物采样, 以研究软体动物的季节演替变化, 结果检出软体动物有4纲49科71属共104种。其中双壳纲39种, 占总数的37.50%; 腹足纲59种, 占总数的56.73%; 多板纲4种, 占总数的3.85%; 头足纲2种, 占总数的1.92%。种类数秋季最多共61种, 春季32种, 夏季51种, 冬季55种。各季节间共有种类数为19~33种。有8个种类为3个季节共有种, 有12个种类为4个季节共有种, 季节种间更替率为0.58~0.76, 春秋季种间更替率最高, 夏秋季种间更替率最低。优势种10种, 其中1个物种为全年优势种, 有2个物种为3个季节共有种。不同物种在潮间带分异明显, 中潮区软体动物有58种, 低潮区软体动物有44种, 高潮区软体动物只有3种。调查显示各季节软体动物栖息密度变化明显, 按降序排列为春季、夏季, 秋季、冬季。季度Shannon-Wiener多样性指数变化范围为2.45~3.42, 年均值为2.99; 季度Pielou物种均匀度指数变化范围为0.53~0.66, 年均值为0.58; 季度Margalef物种丰富度指数变化范围为2.42~4.61, 年均值为3.73; Simpson指数变化范围为0.69~0.80, 年均值为0.76。调查显示各断面均受到不同程度的人为干扰, 相关性分析发现栖息密度与悬浮物和总有机碳正相关(<0.05)。优势种的转变与沿岸上升流的强弱以及养殖废水的不规则排放有关。软体动物的栖息密度与大型海藻的分布呈相反趋势, 低潮区软体动物栖息密度小, 与其结构和食性有关。
软体动物; 季节演替; 硇洲岛; 潮间带; 大型海藻场
硇洲岛位于粤西湛江市东南部的海域上, 距离湛江市区约40 km, 岛屿陆地面积为56 km2, 北临东海岛, 西临雷州半岛, 是典型的潮间带生态系统, 潮间带是海洋与陆地物质循环和能量交换不可或缺的桥梁。其东部沿岸长期受海浪侵蚀, 只有岛南部海岸是砂质岸, 其余岛上地貌均以玄武岩台地为主, 属于火山岛屿(张文杰等, 2012)。其地势为四周向中间逐渐隆起, 因此岛上充足的降雨绝大部分流入海洋, 岛屿周围并无河流, 海水盐度变化不大。气候上属于热带气候, 此地适宜底栖生物生存, 四周水产品丰富。硇洲岛东部也是天然海藻场, 其中以马尾藻、羊栖菜和石莼等居多(张才学等, 2020)。底栖生物被誉为“生态系统工程师”, 其通过生物作用对整个生态系统做出重要贡献(Covich, 1999), 因此也可以作为生态指标的一部分, 用于监控滨海水质的状态(Rossaro, 2007; Miler, 2013; Lazaridou, 2018), 底栖生物的多样性反映出地理环境的多样性。但近几十年发现大型海藻的生物量及丰度大量减少, 同时伴随着底栖软体生物的减少。底栖生物的减少量与生态受到的危害程度成正比, 某些无脊椎动物会将致病菌传播给脊椎动物, 导致其生物量下降, 而部分鱼类的死亡导致生态系统的恶性循环(Covich, 1999), 对底栖生物的研究对于掌握海洋生态系统具有重要作用(李亚芳等, 2018)。目前对于硇洲岛底栖软体生物的研究较少, 对此进行调查有助于海洋生态修复的研究。
1 材料与方法
1.1 样品的采集和分析
1.1.1 采集地点与方法 硇洲岛为火山爆发后产生的岛屿, 属于北热带海洋性气候。从地质来看, 自下而上为第四纪广组滨海沉积物和风化残积土, 表层为大量玄武岩掺杂红色亚黏土(孔中恒, 2004; 张文杰等, 2012)。其潮汐属于半日不规则潮型, 潮高基准为190 cm, 全年平均为192 cm (国家海洋信息中心, 2020)。于2011~2012年在湛江市硇洲岛采集底栖软体动物, 根据岛屿地形以及当地人们活动影响设置5个采样断面, 5个断面地理位置分别为D1 (110.64°E, 20.93°N)、D2 (110.63°E, 20.92°N)、D3 (110.62°E, 20.89°N)、D4 (110.61°E, 20.87°N)和D5 (110.62°E, 20.84°N) (图1)。根据岛屿的地貌设置每条断面6个采样位点, 其中高潮区1个, 所在位置平均潮高为3.50 m; 中潮区分为中上潮区、中潮区和中下潮区共3个, 所在位置平均潮高分别为2.26 m、1.90 m和1.40 m; 低潮区分为低上潮区和低下潮区共2个, 所在位置平均潮高分别为1.10 m和0.80 m。在春季、夏季、秋季和冬季大潮时分别进行底栖软体动物的调查, 定量取样面积为0.25 m×0.25 m, 确定取样具体地点应当是在经过观察之后选取的具有代表性的地点, 采集方法按照《海洋调查规范》(GB/T 12763.6- 2007) (中华人民共和国国家质量监督检验检疫总局等, 2008)进行, 取样时需先将取样框内清理干净, 采集后现场用海水简单清洗, 将取得样品装入密封袋中并加入甲醇固定液, 带回实验室进行分析。
水质样品在每个断面退潮时水深1 m处用5 L聚乙烯塑料桶采集; 叶绿素(chl)样品用1 L聚乙烯瓶在表层采集, 加入10 g/L的MgCO3共5 mL保存, 并在24 h内完成检测; 水温(WT)和pH值采用Thermo Orion520M-01A多参数测量仪现场测定; 盐度(SAL)采用Orion130A盐度计现场测定。
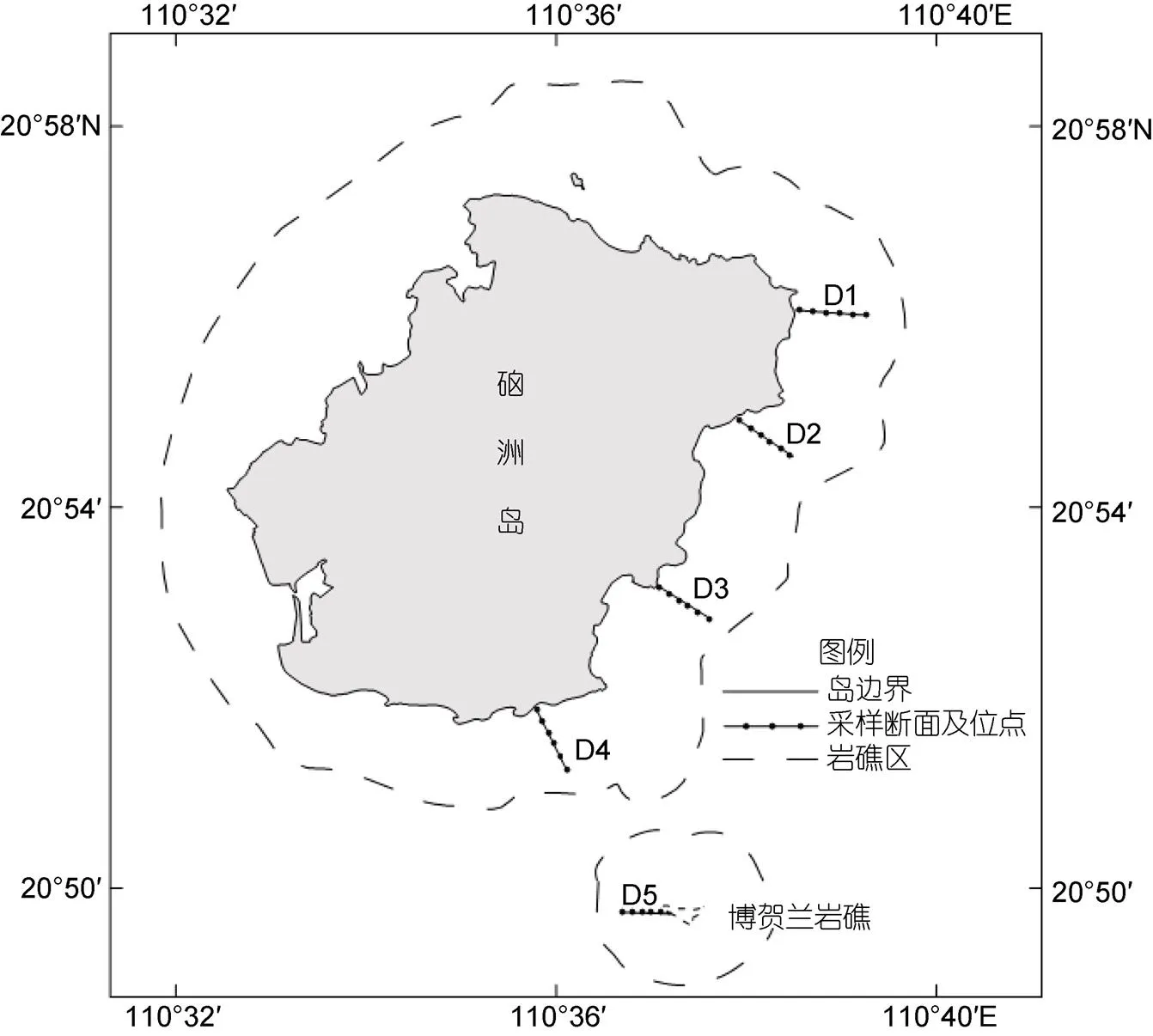
图1 采样断面及其站位
1.1.2 数据处理方法 使用Microsoft Office 2016进行数据初步统计和分析, Origin Pro 2017进行图片绘制, 相关性分析使用SPSS 26.0与Canoco5.0进行分析。
相对重要性指数IRI (Pianka, 1971), 其计算公式为:
IRI=(+)××104, (1)
物种多样性指数采用Shannon-Wiener多样性指数() (徐兆礼等,1989), 其计算公式为:
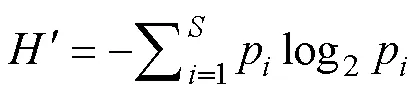
Pielou均匀度指数() (Shannon, 2001), 其计算公式为:

Margalef物种丰富度指数() (Ulanowicz, 2001), 其计算公式为:

Simpson优势度指数() (Pielou, 1966), 其计算公式为:
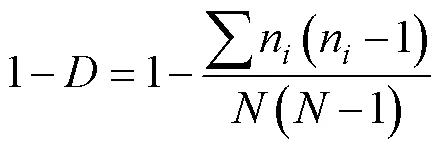
种间更替率(周伟男等, 2013), 其计算公式为:

以上式中,为某一种类的生物量占总生物量的比,为某一种类的个体数占总个数的比,代表物种总数,n为第种的个体数,为第某一种在各站位或各季节间出现的频率, 本文采用季节频率, 视IRI≥1 000为优势种,p为第种在全部采样中的比例,和为两季节的种类数,为两季节的共有种类数,值在0~1之间, 数越大代表群落更替越明显,值为0表示两季节相同,值为1表示两季节无相同种。
2 结果
2.1 硇洲岛软体动物种类组成及优势种演替规律
此次调查发现硇洲岛共有软体动物104种, 其中双壳纲39种, 占总数的37.50%, 栖息密度占总密度的7.76%; 腹足纲59种, 占总数的56.73%, 栖息密度占比为91.32%, 是软体动物的中主要物种; 多板纲4种, 占总数的3.85%, 栖息密度占比为0.88%; 头足纲2种, 占总数的1.92%, 栖息密度占比为0.04%。

表1 硇洲岛底栖软体动物季节种间更替率
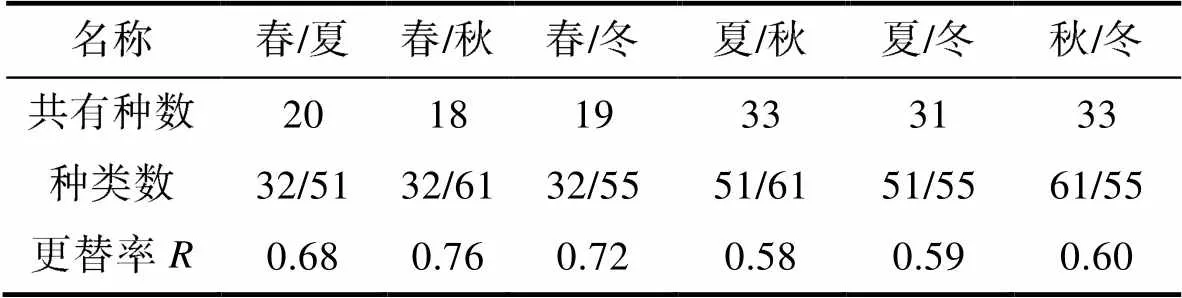
Tab.1 Seasonal interspecies replacement rate of benthic mollusks on Naozhou Island


表2 硇洲岛软体动物季节优势种
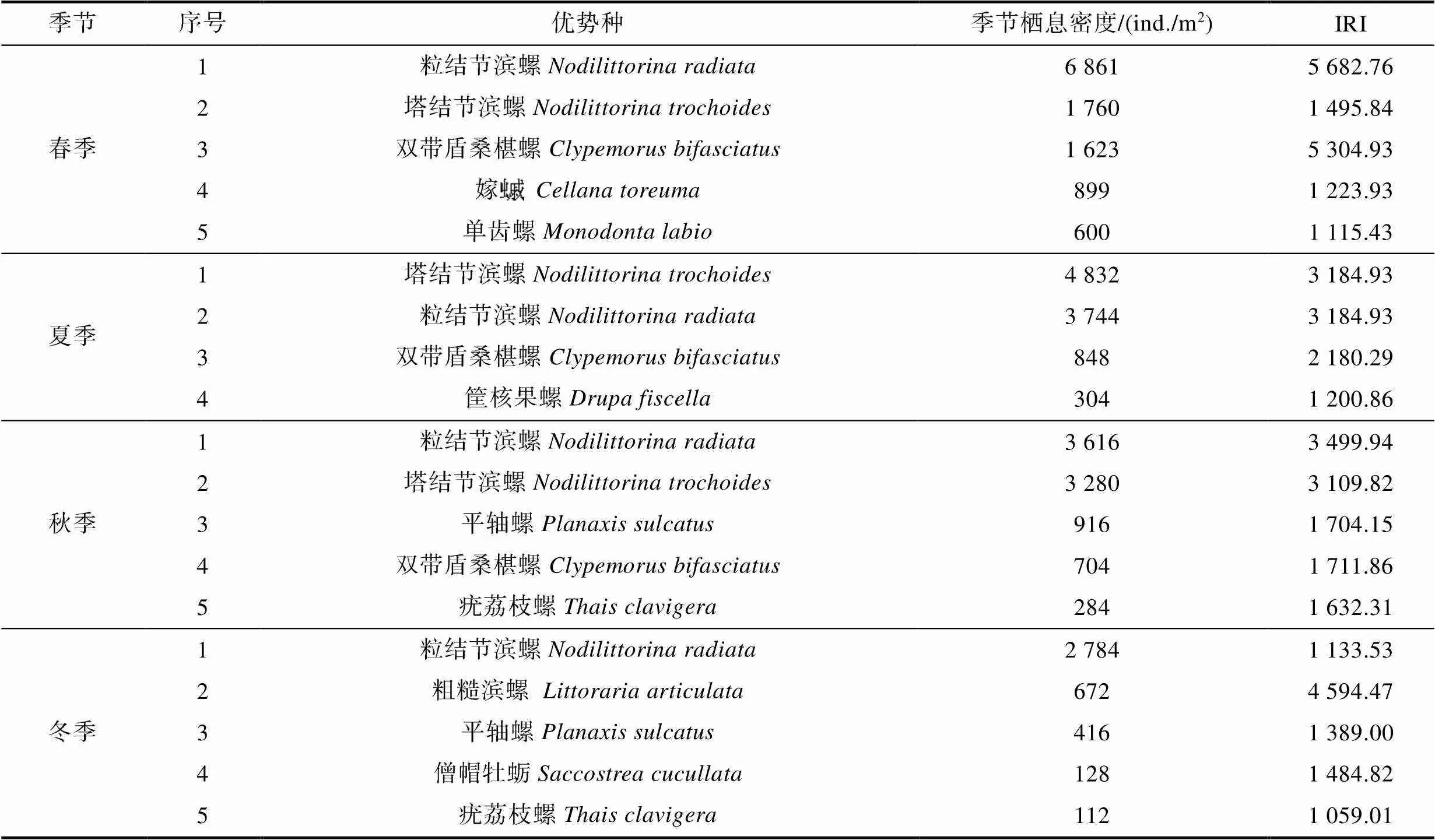
Tab.2 Seasonal dominant species of mollusks in Naozhou Island
2.2 硇洲岛软体动物时空分布与优势群落划分
软体动物分布特征较为明显, 其高潮区种类数为3种, 中上区32种, 中中区32种, 中下区41种, 低上区28种以及低下区34种。其栖息密度存在季节变化: 大体上由高潮区向低潮区逐渐降低, 高潮区栖息密度最大, 低潮区栖息密度最少。春季变化量最大, 高潮区密度最大, 中中区位居第二, 中上区密度较低; 夏季与春季相比略有相似, 在中中区向中下区有一定的回弹; 秋季与冬季相似, 栖息密度由高潮区向低潮区逐渐降低(图2)。

图2 硇洲岛软体动物栖息密度潮间带分布与季节的关系
不同季节的软体动物群落有所不同, 除秋季外的3个季节分为3个群落且相似性均在40%以上, 秋季在35%的相似性上分为3个群落(图3)。


图3 硇洲岛软体动物各季节聚类分析及MDS分析图
秋季群落1为粗糙滨螺-粒结节滨螺-双带盾桑椹螺群落, 优势种为粗糙滨螺、粒结节滨螺、双带盾桑椹螺、疣荔枝螺、筐核果螺、塔结节滨螺、平轴螺、僧帽牡蛎和单齿螺, 包括D1、D3和D4断面; 群落2为疣荔枝螺-筐核果螺-平轴螺群落, 优势种为疣荔枝螺、筐核果螺、平轴螺和单齿螺, 仅包括D2断面; 群落3为粗糙滨螺-疣荔枝螺-筐核果螺群落, 优势种为粗糙滨螺、疣荔枝螺、筐核果螺和僧帽牡蛎, 仅包括D5断面。

2.3 硇洲岛软体动物的物种多样性分析和稳定性
对硇洲岛底栖软体动物的物种多样性分析发现, 各季节软体动物栖息密度存在季节变化趋势, 按降序排列为春季(441.26 ind./m2)、夏季(431.55 ind./m2)、秋季(374.24 ind./m2)和冬季(215.60 ind./m2), 年平均值为365.66 ind./m2(图4)。季度Shannon-Wiener多样性指数变化范围为2.45~3.42, 按降序排列为冬季、秋季、夏季和春季, 年均值为2.99; 季度Pielou物种均匀度指数变化范围为0.53~0.66, 按降序排列为冬季、秋季、夏季和春季, 年均值为0.58; 季度Margalef物种丰富度指数变化范围为2.42~4.61, 按降序排列为秋季、冬季、夏季和春季, 年均值为3.73; Simpson指数变化范围为0.69~0.80, 按降序排列为秋季、冬季、夏季和春季, 年均值为0.76 (表3)。
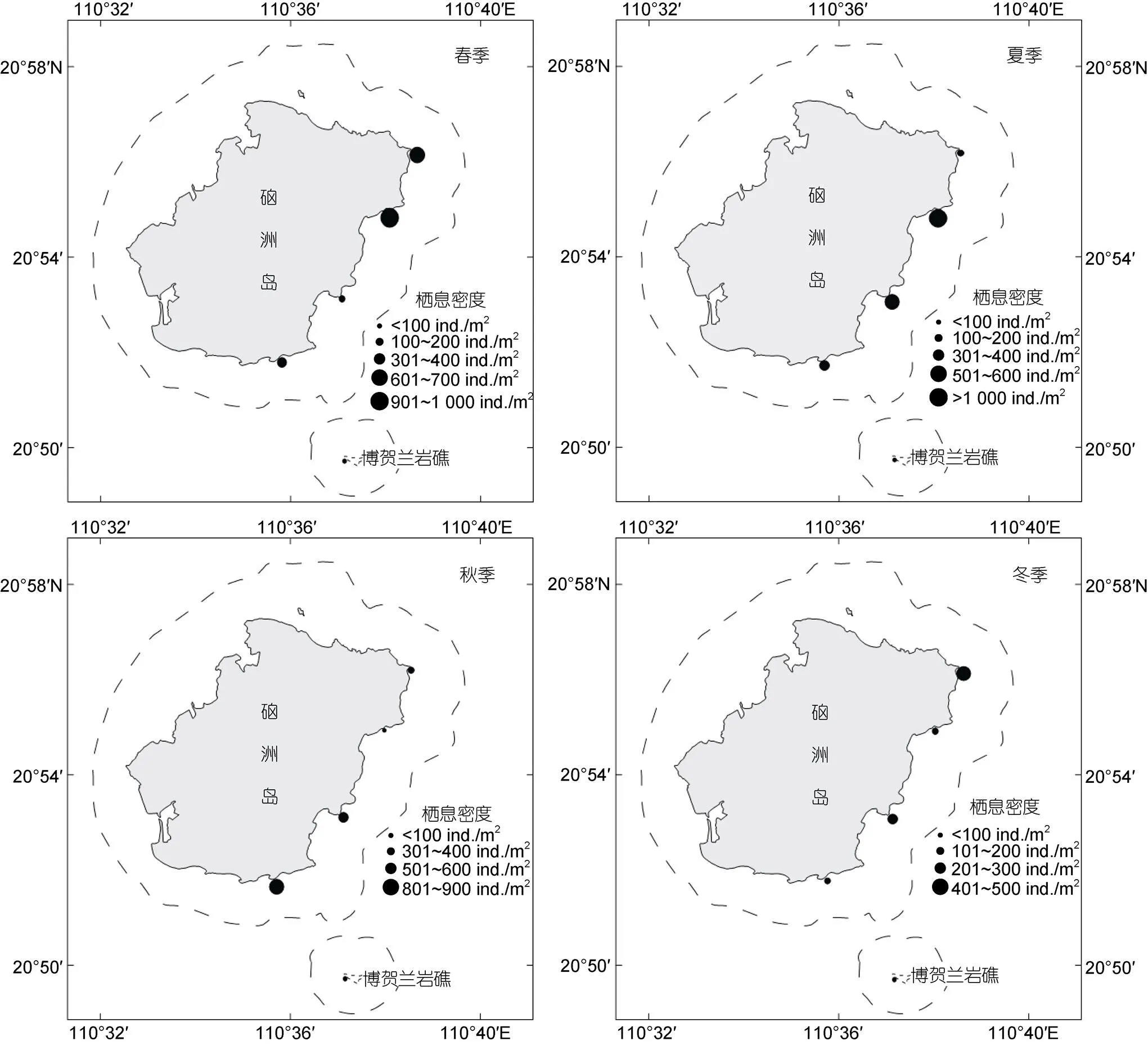
图4 硇洲岛软体动物栖息密度分布气泡图
表3 软体动物多样性分析

Tab.3 Mollusk diversity analysis
通过ABC曲线分析硇洲岛底栖软体动物的稳定性,值(-statistic)反应生物量与栖息密度的关系, 为正值表示群落稳定未受到干扰或者受到微弱干扰, 反之为负值。调查发现D1~D4断面的生物均受到不同程度的干扰, 其栖息密度曲线除D4断面个别交叉点之外, 均在生物量曲线之上, 其值均小于零, D5断面离人类活动区域较远,为正值, 但丰度曲线与生物量曲线有小部分交叉, 表示受到干扰, 只是程度远不及其他断面(图5)。
2.4 硇洲岛软体动物与环境因子的关系
2.4.2 软体动物分布与大型海藻场的关系 硇洲岛东部为大型海藻场, 通过张才学等(2020)的研究综合发现, 底栖软体动物在垂直分布上与大型海藻成相反趋势, 即软体动物栖息密度由高潮区向低潮区逐渐降低, 大型海藻的分布与此相反(图7)。
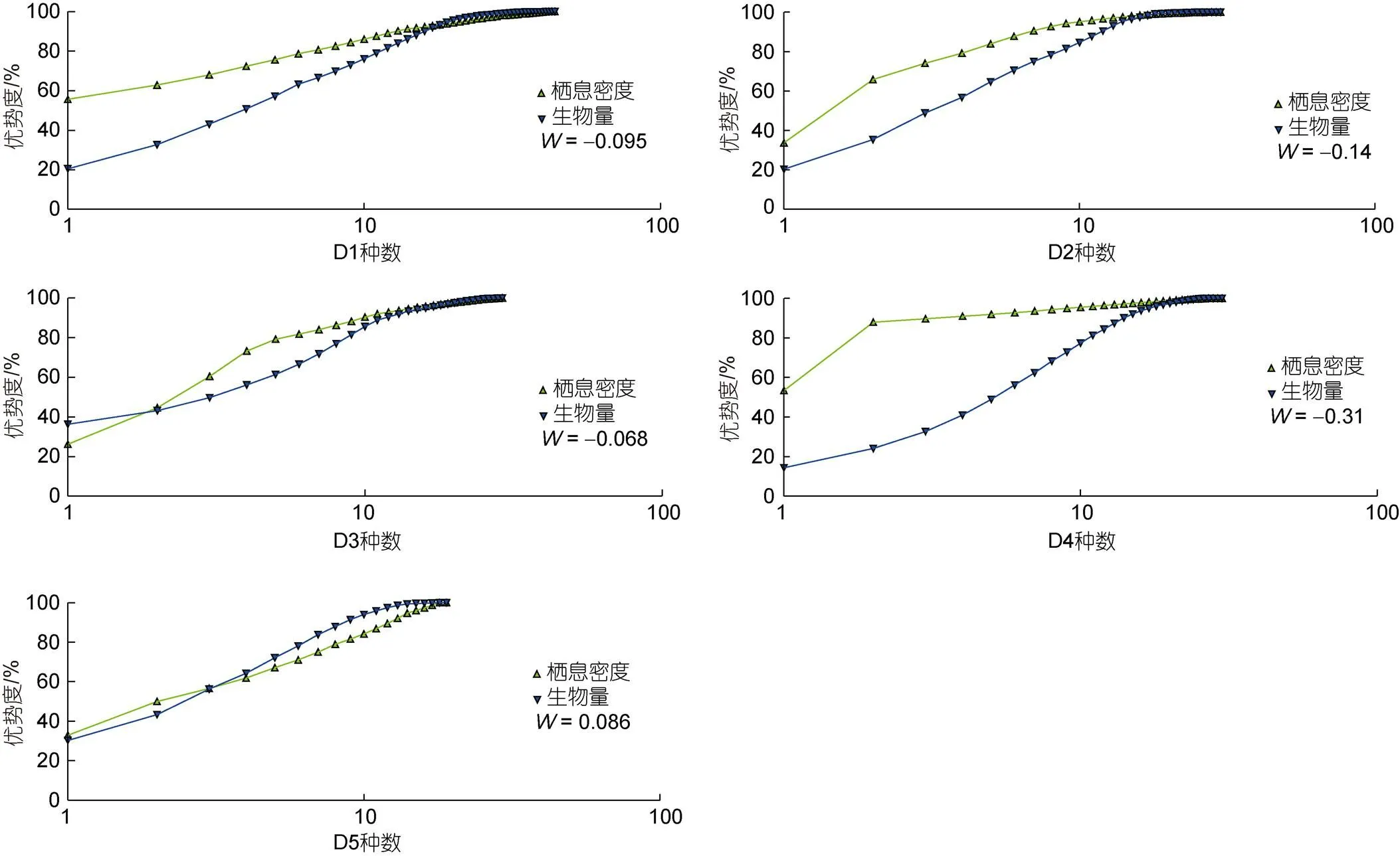
图5 各断面全年ABC曲线图
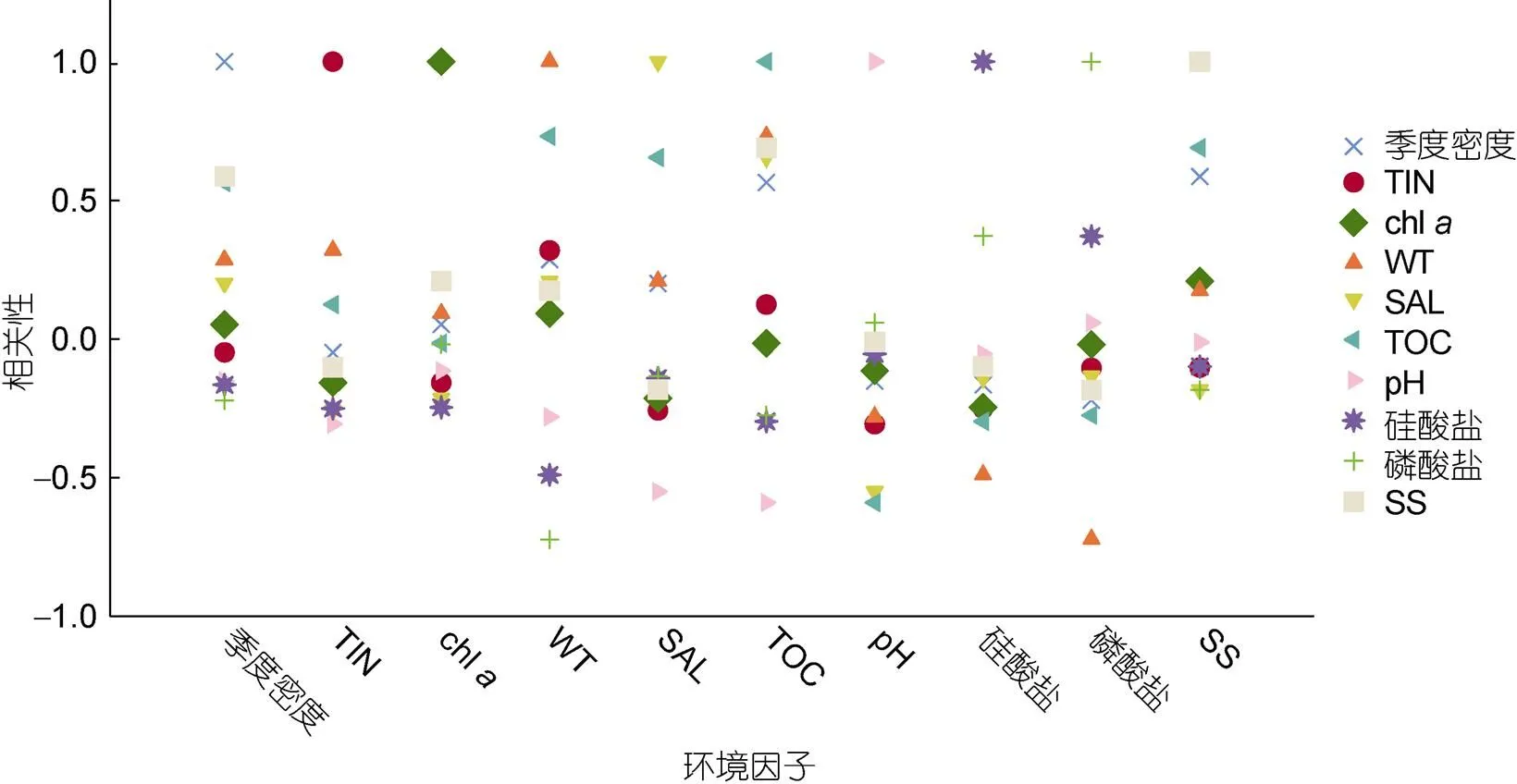
图6 软体动物密度和环境因子的关系

图7 各季节大型海藻生物量的垂直分布和季节变化
3 讨论
3.1 硇洲岛大型海藻场底栖软体动物组成及其分布
硇洲岛地质的表层主要为湖光岩组及其风化残积土(张文杰等, 2012), 不同的地质环境造成底栖生物的构成、栖息密度以及生物量的不同, 本次调查发现底栖软体动物104种, 共4纲, 双壳纲39种, 腹足纲59种, 多板纲4种以及头足纲2种。其种类数量高于国内其他大多数潮间带岛屿(表5)。与林炜等(2002)的研究相比, 软体动物的种类有很大程度的增加, 一方面是因为采集位置的不同, 在硇洲岛的西侧与东海岛隔海相望, 当地通过轮渡和小型渔船的形式通行, 进而对底栖生物产生很大影响, 另一方面采集条件限制导致一定影响。
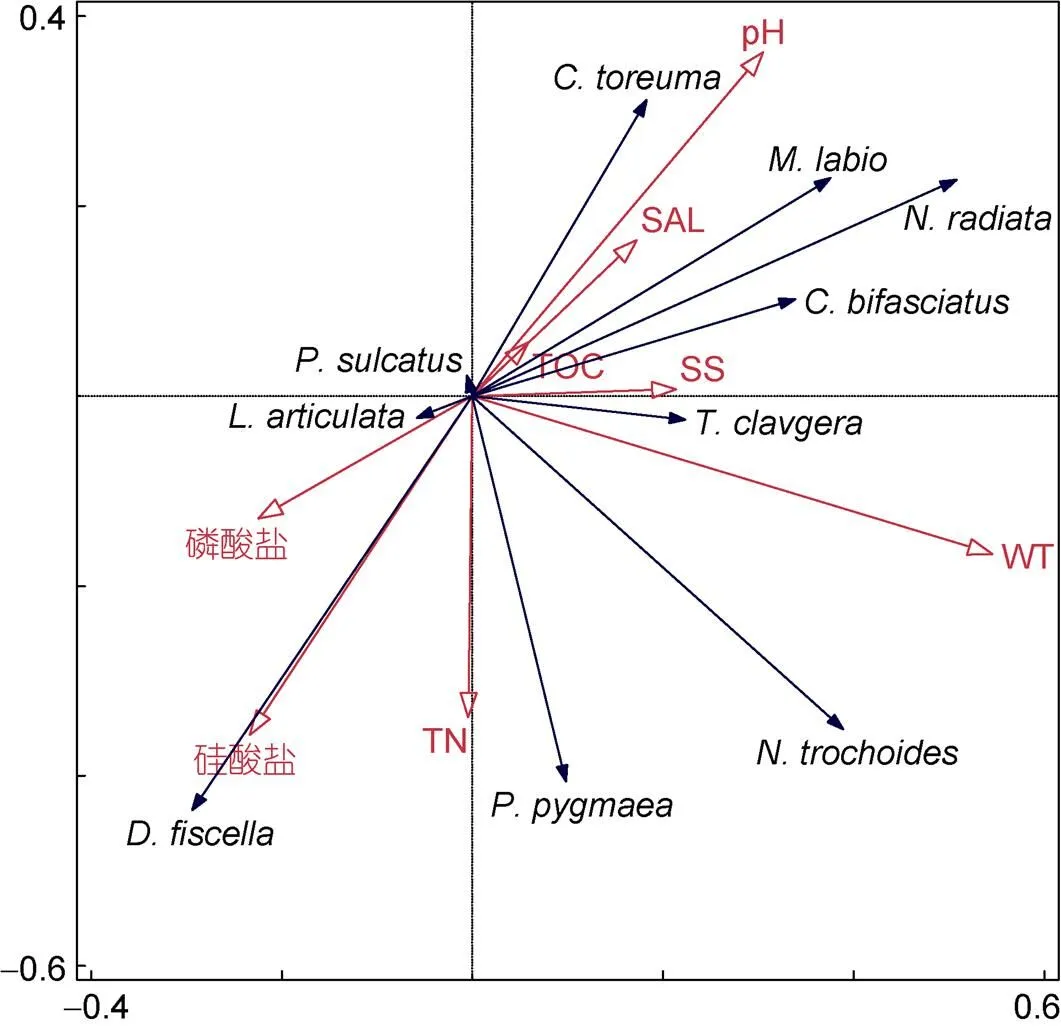
图8 软体动物优势种RDA分析
表5 硇洲岛软体动物与其他海域的比较
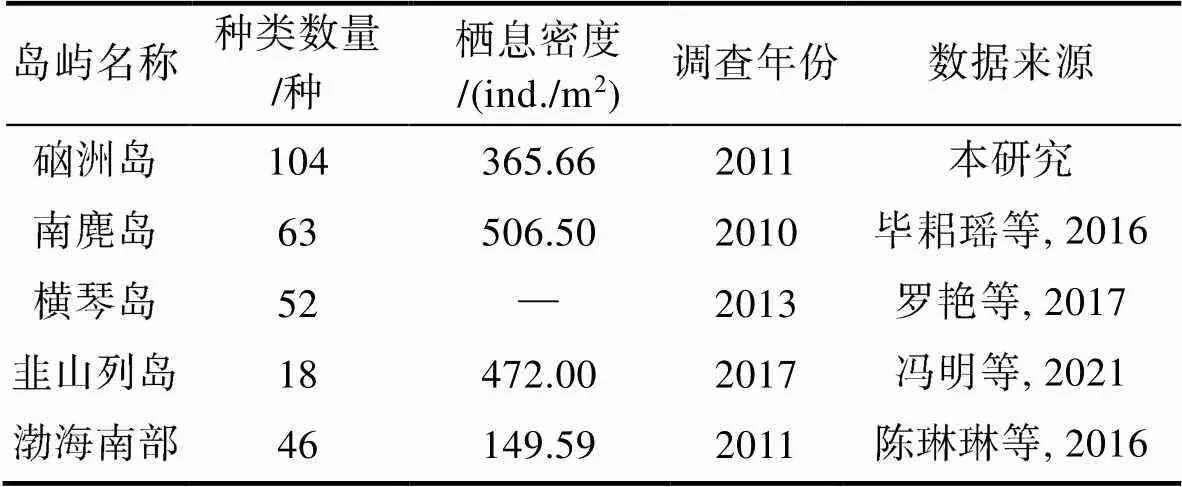
Tab.5 Comparison of mollusks on Naozhou Island and other sea areas
注: “—”表示未发现数据
3.2 硇洲岛大型海藻场底栖软体动物分布影响因素
底栖生物的分布与当地的地形、地质以及覆盖水有着直接的关系(Lucatelli, 2020), 这些非生物属性导致当地环境因子与海洋整体环境的差异。粤西的近岸海流较为特殊, 此区域潮汐模式为不规则半日潮(丁文兰, 1986; Xie, 2015), 整个粤西沿岸通过多因素表明为上升流区域(Lopes, 2005), 在硇洲岛附近其海流运动方向不规则, 但可以看出尺度为100 km的气旋涡特征, 表层海水大致为自东向西的环流, 夏季自下而上产生较弱的上升流(谢玲玲等, 2012; 许金电等, 2013; 王新星等, 2015), 秋季会产生较强的上升流(杨士瑛等, 2003), 上升流会带来SS含量的升高, 在一定程度上影响TOC含量的变化, 为底栖软体动物提供营养物质, 因此在整体上通过相关性分析发现栖息密度与这些因素相关。
硇洲岛海域与其他海域相比, 底栖生物丰富, 人类活动的影响较小。人类活动也会对软体动物产生影响, 岛上没有大型企业, 但在岛屿东部岸线附近有人工池塘, 其养殖废水即使经过处理排放也会对环境产生影响。从图4可看出D5断面由于离硇洲岛有一定距离, 因此受到人类活动影响较小,为正值, 但也有部分生物其栖息密度曲线在生物量曲线之上; 其余断面由于养殖废水的原因, 均受到不同程度的影响, 尤其D2和D4断面受影响严重, 两个断面为养殖废水直排点, 每个断面显示出不同的栖息密度。
4 结论

(2) 软体动物整体上栖息密度与海水悬浮物含量和总有机碳含量呈显著正相关, 优势种的季节变化与上升流相关, 而人类活动对两者均有影响。
(3) 软体动物在垂直分布上与大型海藻成相反趋势, 在低潮区大型海藻占优, 主要藻类为褐藻门中的马尾藻和羊栖菜, 软体动物栖息密度小, 这与动物本身的结构和食性有关, 腹足纲大部分动物依靠齿舌和鳃丝纤毛以褐藻碎片、褐藻代谢产物以及部分小型藻类摄食。
丁文兰, 1986. 南海潮汐和潮流的分布特征[J]. 海洋与湖沼, 17(6): 468-480.
王新星, 于杰, 李永振, 等, 2015. 南海主要上升流及其与渔场的关系[J]. 海洋科学, 39(9): 131-137.
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. 海洋调查规范第6部分: 海洋生物调查: GB/T 12763.6-2007[S]. 北京: 中国标准出版社: 1-157.
孔中恒, 2004. 雷州半岛火山岩的水文地质特征与富水规律[J]. 热带地理, 24(2): 136-139.
冯明, 韩庆喜, 严润玄, 2021. 浙江韭山列岛岩礁潮间带大型底栖动物群落特征和功能性状[J]. 生态学杂志, 40(5): 1469-1477.
毕耜瑶, 蔡厚才, 陈万东, 等, 2016. 南麂岛岩礁潮间带软体动物种类数量变化及其演替[J]. 渔业现代化, 43(3): 65-73.
许金电, 蔡尚湛, 宣莉莉, 等, 2013. 2006年夏季琼东、粤西沿岸上升流研究[J]. 海洋学报, 35(4): 11-18.
杜飞雁, 王雪辉, 贾晓平, 等, 2011. 大亚湾海域大型底栖生物种类组成及特征种[J]. 中国水产科学, 18(4): 877-892.
李亚芳, 杜飞雁, 王亮根, 等, 2018. 粤西海陵湾养殖区邻近海域大型底栖动物生态学特征[J]. 海洋与湖沼, 49(6): 1294-1307.
杨士瑛, 鲍献文, 陈长胜, 等, 2003. 夏季粤西沿岸流特征及其产生机制[J]. 海洋学报, 25(6): 1-8.
张才学, 周伟男, 孙省利, 等, 2020. 硇洲岛大型海藻群落的季节演替[J]. 热带海洋学报, 39(1): 74-84.
张文杰, 谭红兵, 陈喜, 等, 2012. 广东硇洲岛地下水化学演化及成因机理[J]. 水文, 32(3): 51-59.
陈琳琳, 王全超, 李晓静, 等, 2016. 渤海南部海域大型底栖动物群落演变特征及原因探讨[J]. 中国科学: 生命科学, 46(9): 1121-1134.
林炜, 钟海莹, 唐以杰, 2002. 硇洲岛潮间带不同生境底栖软体动物物种多样性研究[J]. 热带海洋学报, 21(3): 14-22.
国家海洋信息中心, 2020. 数据可视化潮汐潮流站位点[EB/OL]. (2022-01-05) [2022-02-08]. http://mds.nmdis.org.cn/pages/ visualization.html.
罗艳, 林丽华, 张翠萍, 等, 2017. 珠海横琴岛海域大型底栖生物的生态特征[J]. 海洋湖沼通报(5): 69-79.
周伟男, 孙省利, 李荣冠, 等, 2013. 湛江湾大型底栖动物的群落结构和多样性特征[J]. 广东海洋大学学报, 33(1): 1-8.
徐兆礼, 陈亚瞿, 1989. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J]. 生态学杂志, 8(4): 13-15, 19.
谢玲玲, 曹瑞雪, 尚庆通, 2012. 粤西近岸环流研究进展[J]. 广东海洋大学学报, 32(4): 94-98.
COVICH A P, PALMER M A, CROWL T A, 1999. The role of benthic invertebrate species in freshwater ecosystems: zoobenthic species influence energy flows and nutrient cycling [J]. BioScience, 49(2): 119-127.
FLORES L N, BARONE R, 1998. Phytoplankton dynamics in two reservoirs with different trophic state (Lake Rosamarina and Lake Arancio, Sicily, Italy) [J]. Hydrobiologia, 369: 163-178.
LAZARIDOU M, NTISLIDOU C, KARAOUZAS I,, 2018. Harmonisation of a new assessment method for estimating the ecological quality status of Greek running waters [J]. Ecological Indicators, 84: 683-694.
LOPES M R M, DE M BICUDO C E, CARLA FERRAGUT M, 2005. Short term spatial and temporal variation of phytoplankton in a shallow tropical oligotrophic reservoir, Southeast Brazil [J]. Hydrobiologia, 542(1): 235-247.
LUCATELLI D, GOES E R, BROWN C J,, 2020. Geodiversity as an indicator to benthic habitat distribution: an integrative approach in a tropical continental shelf [J]. Geo-Marine Letters, 40(6): 911-923.
MILER O, PORST G, MCGOFF E,, 2013. Morphological alterations of lake shores in Europe: a multimetric ecological assessment approach using benthic macroinvertebrates [J]. Ecological Indicators, 34: 398-410.
MILLER R J, PAGE H M, 2012. Kelp as a trophic resource for marine suspension feeders: a review of isotope-based evidence [J]. Marine Biology, 159(7): 1391-1402.
MUYLAERT K, SABBE K, VYVERMAN W, 2000. Spatial and temporal dynamics of phytoplankton communities in a freshwater tidal estuary (Schelde, Belgium) [J]. Estuarine, Coastal and Shelf Science, 50(5): 673-687.
PIANKA E R, 1971. Ecology of the agamid lizardin western Australia [J]. Copeia, 1971(3): 527-536.
PIELOU E C, 1966. Species-diversity and pattern-diversity in the study of ecological succession [J]. Journal of Theoretical Biology, 10(2): 370-383.
ROSSARO B, MARZIALI L, CARDOSO A C,, 2007. A biotic index using benthic macroinvertebrates for Italian lakes [J]. Ecological Indicators, 7(2): 412-429.
SHANNON C E, 2001. A mathematical theory of communication [J]. ACM SIGMOBILE Mobile Computing and Communications Review, 5(1): 3-55.
ULANOWICZ R E, 2001. Information theory in ecology [J]. Computers & Chemistry, 25(4): 393-399.
XIE M X, YAO S S, LI W D,, 2015. Investigation of the movement characteristics of West Guangdong Longshore Ocean Current System, China [J]. Journal of Coastal Research, 73(sp1): 364-368.
RELATIONSHIP BETWEEN SEASONAL SUCCESSION OF MOLLUSC COMMUNITIES AND ENVIRONMENTAL FACTORS IN NAOZHOU ISLAND, SOUTH CHINA SEA
YAO Xin1, SUN Xing-Li1, 2, ZHANG Cai-Xue1, 2, LIAO Hai-Qing1, PENG Yu-Hang1
(1. School of Chemistry and Environment, Guangdong Ocean University, Zhanjiang 524088, China; 2. Marine Resources and Environmental Monitoring Center, Guangdong Ocean University, Zhanjiang 524088, China)
Benthic molluscs were sampled at five locations in the intertidal zone of Naozhou Island in Zhanjiang, Guangdong, in South China Sea from 2011 to 2012 to study their seasonal succession. 104 species of molluscs were identified including 4 classes, 49 families, and 71 genera, of which 59 species were Gastropoda (56.73% of the total species), 39 species of Bivalvia (37.50%), 4 species of Polyplateae (3.85%), and 2 species of Cephalopods (1.92%). The number of species was up to 61 species in autumn, 32 species in spring, 51 species in summer, and 55 species in winter. Only 10 species were seen throughout the whole year, of which 2 species were annual dominant species, and 3 species were common in 3 seasons. There are 19~33 species in each season, 8 species were common in 3 seasons, and 12 species were common in 4 seasons. The seasonal interspecific turnover rate was 0.58~0.76, the highest in spring and autumn, and the lowest in summer and autumn. They were distributed in different tidal zones: 58 molluscs in the mid-tide zone, 44 in the low-tide zone, and only 3 in the high-tide zone. The habitat density of molluscs varied significantly in season in a descending order of spring, summer, autumn, and winter. The Shannon-Wiener diversity index varied from 2.45~3.42 on annual average of 2.99; the Pielou species evenness index ranged 0.53~0.66 on annual average of 0.58; the Margalef species richness index ranged 2.42~4.61 on average annual value of 3.73; the range of Simpson index was 0.69~0.80, and the annual average value was 0.76. Result show that each section was subject to human disturbance in different degrees. Correlation analysis revealed that the habitat density was positively correlated with suspended solids and total organic carbon (<0.05). The change of dominant species was related to the strength of the coastal upwelling and the disorderly discharge of aquaculture wastewater. The abundance of molluscs is inversely proportional to the biomass of large seaweeds. The low tide area of molluscs has a low habitat density, which is related to its structure and feeding habits.
mollusk; seasonal succession; Naozhou Island; intertidal zone; large seaweed field
Q178.1; Q958.8; X171
10.11693/hyhz20211200355
* 国家海洋公益性行业科研专项, 201505027号。姚 昕, 硕士研究生, E-mail: yaoxinnmg@163.com
张才学, E-mail: gdzhangcx@126.com
2021-12-31,
2022-03-13
猜你喜欢
军事文摘(2023年18期)2023-10-31 08:10:50
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
作文周刊·小学四年级版(2022年8期)2022-03-11 07:42:32
食品安全导刊(2021年20期)2021-08-30 06:40:50
今日农业(2020年19期)2020-12-14 14:16:52
小学生必读(低年级版)(2019年5期)2019-08-30 03:22:28
食品与生活(2017年12期)2018-01-09 22:12:34
中国科技信息(2015年2期)2015-11-16 08:18:32
水生生物学报(2015年1期)2015-02-28 16:01:05
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
