
城市绿地中鸟类对海南蒲桃的取食和传播作用
2022-09-17 05:11汪国海董佩佩韦丽娟黄秋婵韩巧唐创斌
生态科学 2022年6期
汪国海, 董佩佩, 韦丽娟, 黄秋婵, 韩巧, 唐创斌,*
城市绿地中鸟类对海南蒲桃的取食和传播作用
汪国海1, 董佩佩1, 韦丽娟2, 黄秋婵1, 韩巧1, 唐创斌1,*
1. 广西民族师范学院 化学与生物工程学院, 广西崇左 532200 2. 广西民族师范学院 数理与电子信息工程学院广西崇左 532200
海南蒲桃()是亚热带地区城市绿地中常见的绿化植物, 其果实数量多、果期长, 可为鸟类提供大量食物资源。2020年6月—8月借助 Safari 10×26 变焦双筒望远镜, 采用焦点扫描法对访问海南蒲桃果实(种子)的鸟类行为进行观察, 详细记录鸟类的种类、取食基质、访问频次、取食时间、取食数量和取食方式等信息, 探讨鸟类在海南蒲桃种子传播及种群更新中的生态作用。结果表明: 成熟的海南蒲桃能吸引7种食果鸟类对其种子进行取食,其中白头鹎()、红耳鹎()、白喉红臀鹎()和黄臀鹎()4种鸟类以整吞的方式取食海南蒲桃的种子, 属于种子潜在传播者。不同种鸟类对海南蒲桃果实的取食频次间存在显著差异(t=4.310,=6,< 0.01), 平均访问只数、平均取食时间和平均取食量间存在极显著差异(< 0.001)。食果鸟类的形态特征(体长、体重和嘴峰大小)与取食数量间均呈正相关。鸟类主要以呕吐的方式传播海南蒲桃的种子, 观察期间在鸟类取食后停歇的树木下方地面上常散落着较多表面洁净、无果肉包裹的裸露种子, 且有大量的海南蒲桃幼苗成功更新, 说明城市绿地中的植物是可以借助鸟类的取食和传播来实现其种群的更新。
海南蒲桃; 食果鸟类; 种子传播; 城市绿地
0 前言
种子传播是决定植物种群空间分布、结构组成及遗传多样性的一个重要过程[1-2]。许多研究表明, 大约70%—90%热带和亚热带区域中的植物种子都是依靠陆生脊椎动物进行取食和传播[3-4], 尤其是数量和种类众多的鸟类。由于鸟类具有较强的飞翔能力且能同时取食和传播多种植物, 加之不同鸟类间的形态特征、取食方式、活动模式和取食后的生境选择偏好等存在明显差异[5], 这些生态功能差异性能提高不同传播者间的互补性[6-7], 提高种子到达适宜萌发微生境中的机会以占据新的生态位[8-9]。同时避免后代因母树周边高强度的种内竞争和自然天敌(病原体、微生物、食草动物和种子捕食者)而造成的高密度死亡率, 从而影响后期植物种群的空间分布和群落多样性的维系[10]。
海南蒲桃()又称乌墨、乌楣, 隶属于桃金娘科(Myrtaceae)、蒲桃属()的常绿乔木, 主要分布于马来西亚、印度、印度尼西亚等国家[11], 而我国主要分布于台湾、福建、广东、广西和云南等地。海南蒲桃果实为椭圆形浆果, 果实期为6—9月[12]; 研究期间对成熟的海南蒲桃果实和种子特征进行了测量, 具体参数如下: 果实的长度(14.583±0.155) mm、宽度(11.472±0.109) mm和重量(1.419±0.038) g, 种子的长度(10.892±0.119) mm、宽度(8.760±0.675) mm和重量(0.491±0.014) g (=100)。较大的外形和重量使其难以借助风力和重力作用离开母树到达较远的地方, 同时其果实成熟后会由青色变为紫黑色, 这符合吸引食果鸟类取食后传播的特征[13-14], 但目前还未见有关鸟类在其种子传播中作用的相关报道。因此, 本研究以分布在城市绿地中的海南蒲桃为研究对象, 通过观察鸟类对海南蒲桃种子的取食行为, 探讨鸟类在其种子传播和种群更新中的生态作用, 以期为城市生态系统中的动植物协同进化取食网络的研究及绿地建设提供基础数据参考。
1 研究区域与方法
1.1 研究区域概况
研究区域位于崇左市境内的广西民族师范学院校园内(22°23'N, 107°23'E)。该区域属于亚热带季风气候区, 气候温和、雨量充沛, 常年光热充足。年平均温度为22 ℃, 7月最高温度为36 ℃, 1月最低温度为 1 ℃; 年降雨量为1200 mm, 有明显的旱季和雨季之分(雨季: 4—9月, 旱季: 10月至翌年3月), 其中80%的降雨量都集中在雨季; 年无霜期达340 d, 年日照时数高达1600 h。校园内种植有大量的果实植物包括海南蒲桃、秋枫()、桂花()、萍婆()、假萍婆()、香樟树()、斜叶榕()和海桐()等。
1.2 研究方法
2020年6月底至8月中旬, 对取食海南蒲桃果实的鸟类行为进行连续观察。选取结实率高且易于观察的5株母树作为目标观察树, 每天上午6:30— 10:30和下午15:30—17:30 时借助 Safari10×26 变焦双筒望远镜(产地: 德国)采用焦点动物扫描法对访问目标母树的鸟类取食行为进行观察, 并详细记录鸟类种类、访问频次、取食时间、取食数量、取食基质(树上或地面)和取食方式(整吞或啄食), 直至其离开观察树为止[15-16]。若一群同种鸟类同时访问目标母树, 又无法对所有鸟类的取食行为进行同步观察时, 则选择最适宜观察的一只个体进行记录[17]。将以整吞的方式取食海南蒲桃果实的鸟类定义为种子传播者[18]。观察周期为种子开始成熟至种子完全掉落, 所有的观察都在晴朗的天气中进行。
采用 One-Sample T-test 对鸟类的访问频次、平均访问只数、平均取食时间及平均取食数量差异进行分析; 鸟类形态特征(体重、体长和嘴峰)的所有参数来自《中国鸟类野外手册》[19]和《中国鸟类志》[20], 并对鸟类形态特征与取食量间的关系进行相关性分析。所有数据的分析均在SPSS 20.00上完成, 并将显著水平设定为< 0.05。采用R语言(3.6.1)进行作图[21]。
2 结果与分析
研究期间共观察到7种鸟类(1目4科)访问目标母树308次, 不同种鸟类对海南蒲桃果实的访问频次存在显著差异(t=4.310,=6,< 0.01), 而平均访问只数(t=10.478,=6,< 0.001)、平均取食时间(t=12.846,=6,< 0.001)和平均取食量(t=8.817,=6,< 0.001)都存在极显著差异 (表 1)。
从访问频次上来看, 白头鹎()、暗绿绣眼鸟()和红耳鹎() 3种鸟类的访问频次最高, 分别占总访问频次的21.75%、21.10%和19.81%(表1)。
从取食方式上来看, 4种鹎科鸟类同时以啄食和整吞的方式取食海南蒲桃的果实, 对海南蒲桃的种子具有潜在的传播作用; 而其余3种鸟类仅啄食果肉, 不起传播作用(表1; 图1)。从取食基质上来看, 仅1种鸟类即鹊鸲()在地面上取食海南蒲桃的果实。在鸟类常停歇的树木下方地面上散落着大量表面洁净、无果肉包裹的海南蒲桃种子, 但在鸟粪中未发现海南蒲桃的种子, 因此推断这些整吞的食果鸟类均以呕吐的方式传播海南蒲桃的种子。
相关分析表明, 食果鸟类的体长、体重和嘴峰长与果实平均取食量间均呈正相关, 说明鸟类的形态越大, 取食的果实数量越多(图2)。
3 讨论
自然界中的植物为尽可能的吸引更多的食果鸟类对其果实或种子进行取食和传播, 会不断的进化出一系列的适应特征, 如颜色、大小、形状、营养组成和成熟时间等[22-23], 其中颜色尤其是红色和黑色是植物最常用的视觉吸引剂[24]。本研究中, 海南蒲桃果实成熟后由青色转变为紫黑色, 使其能与周围植物的绿色背景形成强烈的视觉反差, 从而对空中飞行的鸟类起着强烈的视觉吸引作用[25]。此外, 鸟类对植物果实或种子的取食方式常会随果实或种子尺寸的大小而发生改变[16]。如鸟类会直接整吞小型种子后以粪便的形式将其排出体外, 而常以呕吐的方式传播尺寸较大的种子, 从而有利于植物种子到达适宜萌发的生境中以实现其种群的自然更新[26]。本研究中, 4种鹎科鸟类均以整吞的方式取食海南蒲桃的果实, 属于种子潜在传播者(表1; 图1)。大多数鸟类取食完海南蒲桃的果实后, 会直接停歇在母树周边的木棉()、海桐和假萍婆等树上, 待消化完果肉部分后再以呕吐的方式将种子排出体外; 同时在鸟类停歇的树木下方也发现有大量表面洁净、无果肉包裹的种子和少量的更新幼苗, 说明鸟类的取食行为的确能促进海南蒲桃种群的更新, 二者间能形成稳定的互利互惠关系。
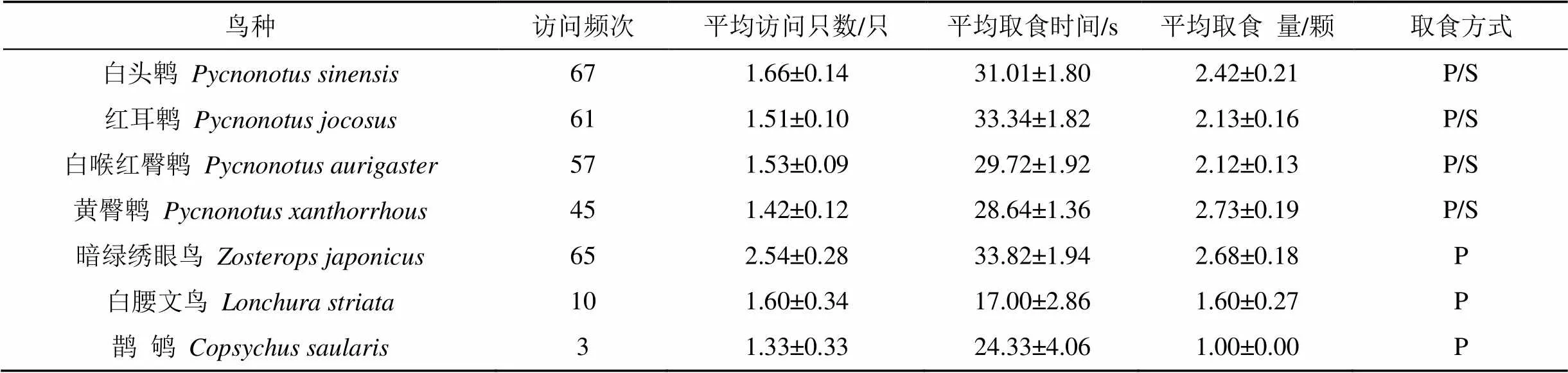
表1 取食海南蒲桃果实的鸟类种类
注: 取食方式: P表示啄食; S表示整吞; 所有数据均为平均值±标准误(Mean±SE)。

注: 按其访问频次的比例从小到大依次排列; 中间线条的粗细表示两者间对应关系的强度; a.白头鹎; b.红耳鹎; c.白喉红臀鹎; d.黄臀鹎; e.暗绿绣眼鸟; f.白腰文鸟; g.鹊鸲。
Figure 1 Foraging network between frugivorous birds andfruits
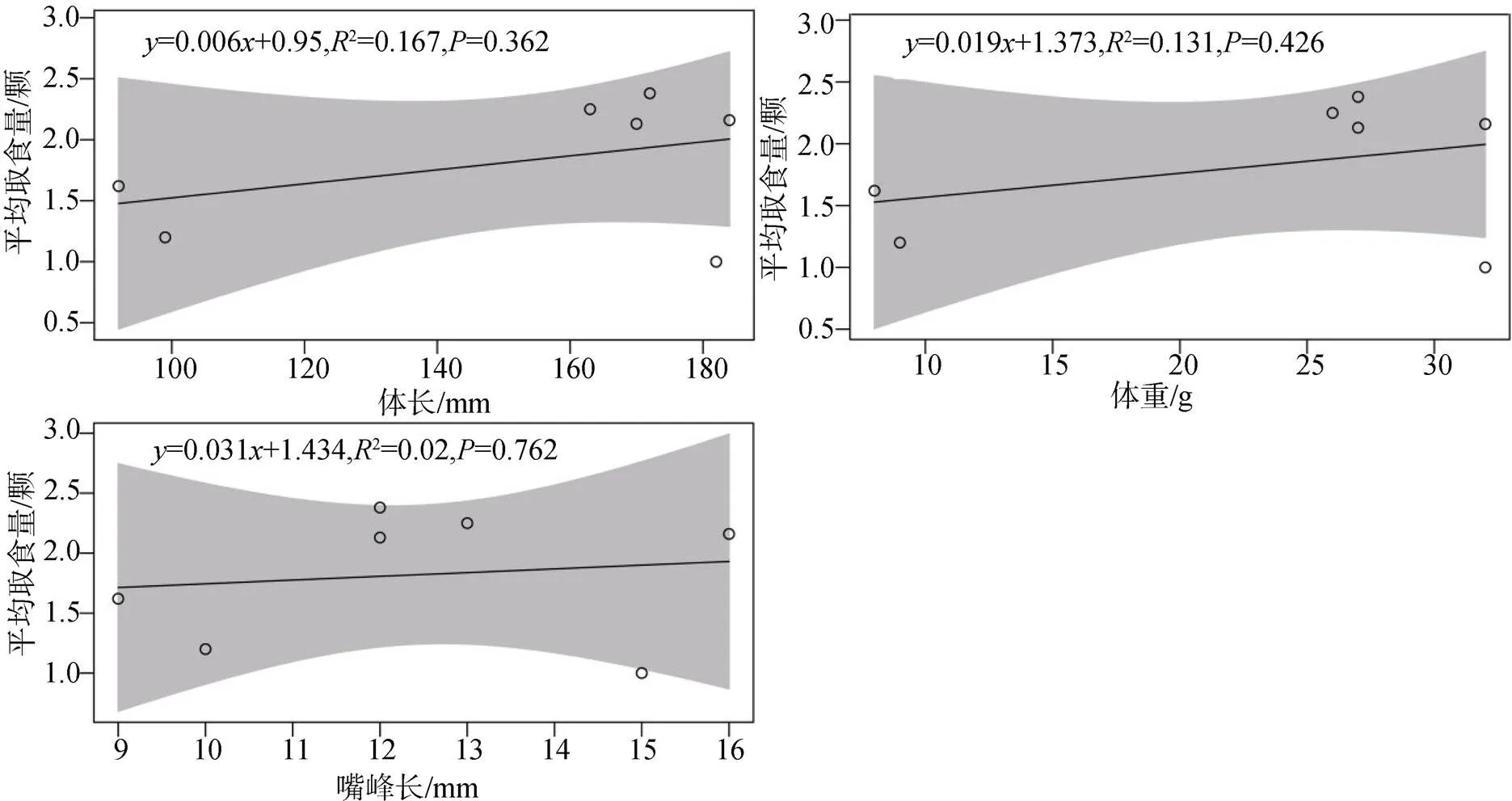
图 2 取食量与鸟类形态特征的相关性
Figure 2 Relationship between foraging quantity and morphological characteristics of birds
鸟类的形态特征(体重、体长和嘴峰)会影响其对植物果实或种子的取食数量[27]。许多研究表明, 鸟类对种子的取食数量与其形态特征间呈显著正相关[28-29]。这是因为鸟类的形态越大意味着其具有更大的肠道容量, 能一次性取食更多的植物种子, 加之大型鸟类具有较强的飞行能力和活动范围, 能将植物种子传播到不同的生境中, 从而提高植物种群的空间分布[30]。本研究中, 鸟类的体重、体长和嘴峰与鸟类的取食数量间都呈正相关关系(图2), 说明形态特征数据越大, 鸟类的取食数量越多。
本研究结果表明, 城市绿地中的海南蒲桃是有能力与当地鸟类形成稳定的协同进化关系, 这为后期研究城市绿地中的动植物协同进化网络及绿地建设具有重要意义。同时, 成熟的海南蒲桃果实富含大量的营养, 除了鸟类外是否有其他动物(啮齿动物、蚂蚁等)对其果实进行取食和搬运还需进一步研究。
[1] HELENO R H, RAMOSJ A, MEMMOTT J. Integration of exotic seeds into an Azorean seed dispersal network [J]. Biological Invasions, 2013, 15: 1143–1154.
[2] FONTURBRL F E, JORDANO P, MEDEL R. Scale- dependent responses of pollination and seed dispersal mutualisms in a habitat transformation scenario [J]. Journal of Ecology, 2015, 103: 1334–1343.
[3] ARROYO R V, AGUILAR B E, GONZALEZ Z A, et al. Parent-parent and parent-offspring distances inseeds suggest long-distance pollen and seed dispersal: evidence from latrines of the spider monkey [J]. Journal of Tropical Ecology, 2017, 33(2): 95–106.
[4] CARLO T A, TEWKSBURY J J, REES M. Directness and tempo of avian seed dispersal increases emergence of wild chiltepins in desert grasslands [J]. Journal of Ecology, 2014, 102(1): 248–255.
[5] LI N, BAI B, Li X H, et al. Dispersal of remnant endangered trees in a fragmented and disturbed forest by frugivorous birds [J]. Journal of Plant Research, 2017, 130: 669–676.
[6] BREGMAN T P, LEES A C, MACGREGOR H E A, et al. Using avian functional traits to assess the impact of land-cover change on ecosystem processes linked to resilience in tropical forests [J]. Proceedings of the Royal Society B: Biological Sciences, 2016, 283: 20161289.
[7] SAAVEDRA F, HENSEN I, BECK S G, et al. Functional importance of avian seed dispersers changes in response to human-induced forest edges in tropical seed-dispersal networks [J]. Oecologia, 2014, 176(3): 837–848.
[8] BECKMAN N G, ROGERS H S. Consequences of seed dispersal for plant recruitment in tropical forests: interactions within the seedscape [J]. Biotropica, 2013, 45(6): 666–681.
[9] DURON Q, GARCIA I O, BRESCIA F, et al. Comparative effects of native frugivores and introduced rodents on seed germination in New-Caledonian rainforest plants [J]. Biological Invasions, 2017, 19(1): 351–363.
[10] COMITA L S, QUEENBOROUGH S A, MURPHY S J, et al. Testing predictions of the Janzen-Connell hypothesis: a meta-analysis of experimental evidence for distance-and density-dependent seed and seedling survival [J]. Journal of Ecology, 2014, 102(4): 845–856.
[11] AZIZ A, BANERJEE S. Phytochemical Screening and Antibacterial Activity study of(Myrtaceae) Seed Extracts[J]. Pharma Tutor, 2018, 6(4): 70–73.
[12] 周菊珍, 杜铃, 林榕庚. 海南蒲桃的栽培技术 [J]. 广西林业科学, 2001, 30(2): 99–100.
[13] DUAN Q, QUAN R. The effect of color on fruit selection in six Tropical Asian birds [J]. The Condor, 2013, 115(3): 623–629.
[14] DU Y J, MI X C, Liu X J, et al. Seed dispersal phenology and dispersal syndromes in a subtropical broad-leaved forest of China [J]. Forest Ecology and Management, 2009, 258(7): 1147–1152.
[15] 陆彩虹, 鲁长虎. 南京中山植物园鸟类对香樟果实(种子)的取食 [J]. 动物学杂志, 2019, 54(6): 784–792.
[16] 潘扬, 徐丹, 鲁长虎, 等. 食果鸟类对红楠种子的传播作用[J]. 生态科学, 2017, 36(2): 63–67.
[17] 李宁, 王征, 鲁长虎, 等. 斑块生境中食果鸟类对南方红豆杉种子的取食和传播 [J]. 生态学报, 2014, 34(7): 1681–1689.
[18] BREITBACH N, LAUBE I, STEFFAN D I, et al. Bird diversity and seed dispersal along a human land-use gradient: high seed removal in structurally simple farmland [J]. Oecologia, 2010, 162(4): 965–976.
[19] 约翰.马敬能, 卡伦.菲利普斯,等. 中国鸟类野外手册 [M]. 长沙: 湖南教育出版社, 2000.
[20] 赵正阶. 中国鸟类志 [M]. 长春: 吉林科学技术出版社, 2001.
[21] R Core team. R: A language and environment for Statistical Comupting [M]. R Foundation for Statistical computing, Vienna, Austria, 2019.
[22] BURNS K C, DALEN J L. Foliage color contrasts and adaptive fruit color variation in a bird-dispersed plant community [J]. Oikos, 2002, 96: 463–469.
[23] HERRERA C M. Plant-vertebrate seed dispersal systems in the Mediterranean: ecological, evolutionary, and historical determinants [J]. Annual Review of Ecology and Systematics, 1995: 705–727.
[24] SCHAEFER H M, MCGRAW K, CATONI C. Birds use fruit color as honest signal of dietary antioxidant rewards [J]. Functional Ecology, 2008, 22: 303–310.
[25] DUAN Q, GOODALE E, QUAN R C. Bird fruit preferences match the frequency of fruit colours in tropical Asia [J]. Scientific Reports, 2014, 4: 5627.
[26] JORDANO P, GARCIA C, GODOY J A, et al. Differential contribution of frugivores to complex seed dispersal patterns [J]. Proceedings of the National Academy of Sciences, 2007, 104(9): 3278–3282.
[27] SIMMONS B I, SUTHERLAND W J, DICKS L V, et al. Moving from frugivory to seed dispersal: incorporating the functional outcomes of interactions in plant-frugivore networks [J]. Journal of Animal Ecology, 2018, 87(4): 995–1007.
[28] DONOSO I, SCHLEUNING M, GARCIA, et al. Defaunation effects on plant recruitment depend on size matching and size trade-offs in seed-dispersal networks [J]. Proceedings of the Royal Society B: Biological Sciences, 2017, 284(1855): 20162664.
[29] LI N, WANG Z, Li X H, et al. Bird functional traits affect seed dispersal patterns of China's endangered trees across different disturbed habitats [J]. Avian Research, 2018, 9(1): 13.
[30] PEREZ M N, JORDANO P, VALIDO A. Downsized mutualisms: Consequences of seed dispersers' body-size reduction for early plant recruitment [J]. Perspectives in Plant Ecology, Evolution and Systematics, 2015, 17(2): 151–159.
Fruit foraging and dispersal ofby frugivorous birds in urban green space
WANG Guohai1, DONG Peipei1, WEI Lijuan2, HUANG Qiuchan1, HAN Qiao1, TANG Chuangbin1,*
1. College of Chemistry and Bioengineering, Guangxi Normal University for Nationalities, Chongzuo 532200, Guangxi, China 2. College of Mathematics, Physics and Electronic Information Engineering, Guangxi Normal University for Nationalities, Chongzuo 532200, Guangxi, China
Jambolan jam () is a common greening plant in urban green space with abundant fruits and a long fruit period, which is widely distributed in subtropical area. With Safari l0×26 zoom binoculars, the focus scanning method was adopted to observe the bird behavior of visiting Jambolan jam from June to August 2020. The information of bird species, foraging substrates, visiting frequency, foraging time, foraging amount and foraging methods was recorded to explore the ecological role of birds on seed dispersal and population regeneration of Jambolan jam. The results showed that Jambolan jam cloud attracted 7 species of frugivorous birds to feed on its fruit, and there were significant differences in the visiting frequency among different bird species (t=4.310,=6,<0.01). There were significant differences in the average number of visitors, average foraging time andaverage foraging amount among different birds (<0.001). Chinese Bulbul (), Red-whiskered Bulbul (),Sooty-headed Bulbul () and Brown-breasted Bulbul () swallowed the whole fruit, which were considered the potential seed dispersers of Jambolan jam. The morphological characteristics (body weight, body length and bill length) of birds were positively correlated with the foraging amount of the fruits. These birds dispersed seeds primarily by regurgitating them to the ground, many seeds with clean surface and no pulp wrapped, and regeneration seedlings were found under the trees acting as roosting site of birds during the observation period, which indicated that the plants in urban green space could realize the population regeneration by feeding and dispersal of birds.
Jambolan jam (); frugivorous bird; seed dispersal;urban green space
10.14108/j.cnki.1008-8873.2022.06.012
Q958.1
A
1008-8873(2022)06-100-05
2020-09-30;
2020-12-05
广西民族师范学院博士科研启动项目(2018FG008, 2021BS002); 珍稀濒危动植物生态与环境保护重点实验室研究基金(20201125);广西壮族自治区教育厅第四批民族院校特色学科建设立项建设学科项目(民族生态学)
汪国海(1986—), 男, 博士, 主要从事动植物协同进化研究, E-mail:1016729581@qq.com
通信作者:唐创斌, 男, 博士, 讲师, 主要从事野生动物生态与保护研究, E-mail: 1198407963@qq.com
汪国海, 董佩佩, 韦丽娟, 等. 城市绿地中鸟类对海南蒲桃的取食和传播作用[J]. 生态科学, 2022, 41(6): 100–104.
WANG Guohai, DONG Peipei, WEI Lijuan, et al. Fruit foraging and dispersal ofby frugivorous birds in urban green space[J]. Ecological Science, 2022, 41(6): 100–104.
猜你喜欢
山东林业科技(2021年4期)2021-08-31
花火彩版A(2021年10期)2021-01-03
现代园艺(2020年17期)2020-08-31
天然产物研究与开发(2018年11期)2018-11-30
飞魔幻A(2018年9期)2018-10-26
现代园艺(2018年1期)2018-03-15
飞魔幻B(2018年11期)2018-02-26
现代园艺(2017年19期)2018-01-19
现代园艺(2017年23期)2018-01-18
现代园艺(2017年21期)2018-01-03
