
凤眼莲入侵程度对金鱼藻和黑藻生长及种间关系的影响
2022-09-13 08:40蔡竟芳高海燕沈一栾阿斯哈李红丽
生态学报 2022年16期
张 羽,蔡竟芳,万 云,高海燕,孙 凯,沈一栾,阿斯哈,李红丽 ,*
1 北京林业大学生态与自然保护学院,北京 100083 2 南京大学生命科学院,南京 210073 3 鄂尔多斯生态环境职业学院,鄂尔多斯 017010
生物入侵已成为破坏湿地生态系统日益严重的生态问题,是继生境破坏和破碎化之外对生物多样性有严重影响的又一威胁因素[1],外来入侵种能够在短时间内占据广泛的空间快速繁殖,引起本地物种种群特性的变化,造成本地物种的消失甚至灭绝,破坏生态系统稳定性[2]。外来入侵种往往会导致生境破碎化,通过竞争或者直接占据本地种生态位从而排挤本地种,或者与本地种竞争资源,使本地物种的种类和数量减少,物种多样性降低,一些种群的遗传多样性降低,甚至导致本土物种濒危或灭绝[3—4]。凤眼莲(Eichhorniacrassipes)被称为水域的“癌细胞”,扩展蔓延速度快,可以在很短时间内渡过适应期,依靠其繁殖优势和扩展能力,迅速增加种群数量,很快在入侵成功的生态系统中占据重要生态位,对原生态系统造成破坏[5]。研究表明凤眼莲可以在水面扩散形成致密层,降低水下植物的可利用光资源,占据生态位和生境,使本地种失去竞争能力[6—7],进而导致沉水植物种间关系改变,甚至会引发沉水植物的死亡。沉水植物的死亡又使水体中溶解氧含量降低,导致其他鱼类死亡,使生态链失去平衡[8]。
沉水植物是河流、湖泊等湿地生态系统中的重要初级生产者,为湿地动物提供丰富的食物,还可以为它们提供栖息环境和繁殖场所[9],对生态系统的健康和稳定有着重要作用[10—11]。沉水植物在水体中占据的生态位相同,生长期也大致相近,必然存在种间竞争关系[12]。植物间的相互作用直接影响植物个体的生长发育、形态特征,同时也影响植物群落的分布乃至整个生态系统的结构和功能[13—14]。并且,植物间相互作用的关系和强度也会随着环境因子的变化而变化[15—16]。其中湿地植物入侵也是影响沉水植物生长和种间关系变化的环境因子之一。外来植物入侵过程通常被分为引入阶段、建群阶段和扩散阶段,但在实际调查中很难界定这不同阶段的界限和范畴。故此,研究中更多使用入侵种覆盖范围、植株密度等指标来实现入侵程度的衡量[17],入侵植物重要值涵盖入侵植物的数量、高度、盖度,众多研究采用此指标来分析入侵程度对本地物种的影响[18—19]。而在外来植物入侵过程中对本地种的影响并非单纯的抑制作用[20—21],李安定等人对空心莲子草(Alternantheraphiloxeroides)的研究发现本地种多样性指数和均匀度指数均表现为随空心莲子草重要值先增加后减少,并且在水生和湿生环境中莲子草的临界重要值也不同[22]。而关于凤眼莲入侵程度对沉水植物生长和种间关系的研究鲜见报道。
目前针对外来植物凤眼莲的入侵机制,以及入侵植物对本地单一沉水植物的影响有一定的报道,而关于凤眼莲入侵程度对沉水植物生长及种间关系影响的研究相对缺乏。本文通过控制实验,设计入侵植物凤眼莲不同入侵程度(以凤眼莲不同株数形成水面覆盖程度设定),定量分析研究凤眼莲入侵程度对沉水植物生长和种间关系的影响。试图解决以下问题:(1)凤眼莲不同入侵程度对沉水植物生长指标的影响如何变化?(2)凤眼莲不同入侵程度对沉水植物种间关系有什么影响?(3)凤眼莲不同入侵程度对水质如何影响?(4)凤眼莲入侵、沉水植物生长及水体理化性质之间如何相互影响?
1 材料与方法
1.1 实验材料
金鱼藻(Ceratophyllumdemersum)是金鱼藻科金鱼藻属的沉水性多年生水草,花期6—7月,果期8—9月。金鱼藻多生长于小湖泊静水处,于池塘、水沟等处常见,生命力较强,适温性较广,在水温低至4℃时也能生长良好。为鱼类的饵料,也可作为猪的饲料[23]。
黑藻(Hydrillaverticillata)是水鳖科黑藻属的单子叶多年生沉水植物,花果期5—10月。广布于池塘、湖泊和水沟中。喜阳光充足的环境,环境荫蔽则植株生长受阻,在15—30℃的温度范围内生长良好,越冬不低于4℃。适宜浅水绿化、室内水体绿化,是良好的沉水观赏植物[24]。
凤眼莲(Eichhorniacrassipes)是雨久花科凤眼蓝属的漂浮性水生植物,被列入世界百大外来入侵种之一,广布于中国长江、黄河流域及华南各省。喜欢温暖湿润、阳光充足的环境,适应性很强。适宜水温18—23℃,超过35℃也可生长,气温低于10℃停止生长,具有一定耐寒性。喜欢生于浅水中,随水漂流,繁殖迅速。生命力顽强,在干旱环境下种子也可保存15—20a[25]。
1.2 实验设计
实验所需要的沉水植物及相关实验材料均取自北京奥林匹克森林公园,并协同水体,带回实验区域进行培养。
实验采用双因素设计,包括凤眼莲入侵程度3种模式和邻株状态3种定植模式。设置凤眼莲入侵程度3种模式,包括无入侵,入侵程度盖度约25%(以凤眼莲入侵植物1株),入侵程度盖度约75%(以凤眼莲入侵植物3株)。邻株状态3种定植模式,分别为沉水植物金鱼藻无种间竞争模式、沉水植物黑藻无种间竞争模式、两个物种混种存在种间竞争模式。实验共9种处理,分别是:(1)无入侵金鱼藻单种模式;(2)无入侵黑藻单种模式;(3)无入侵金鱼藻和黑藻混种模式;(4) 凤眼莲盖度约25%金鱼藻单种模式;(5) 凤眼莲入侵盖度约25%黑藻单种模式;(6) 凤眼莲入侵盖度约25%金鱼藻和黑藻混种模式;(7) 凤眼莲盖度约75%金鱼藻单种模式;(8) 凤眼莲入侵盖度约75%黑藻单种模式;(9) 凤眼莲入侵盖度约75%金鱼藻和黑藻混种模式(图1)。
2018年8月20日,实验植株均取自北京奥林匹克森林公园。将生长健壮,长势均一的金鱼藻与黑藻分别剪取约8 cm长的带芽茎段作为实验材料,定植在的实验容器(上径29 cm,下径27 cm,高38 cm)中,实验基质底泥采自北京市野鸭湖湿地公园。基质深度约为10 cm,有机质含量为 (0.797±0.050) g/kg,总氮含量为 (0.105±0.030) g/kg,总磷含量为 (0.146±0.060) g/kg。植株种植完毕在桶内水体表面放置0株、1株或3株入侵植物凤眼莲,分别模拟凤眼莲入侵程度为0、25%和75%的沉水植物生长环境(图1)。实验设置5组重复。每2—3天浇水一次,水体为未经处理的普通自来水。
实验于北京市海淀区北京林业大学三顷园苗圃内进行,实验时间为2018年8月27日至10月13日,共48天。
实验结束收获前首先用取水管于每个处理取15 mL水样用于水质检测,然后小心取出植物进行生长指标测定。
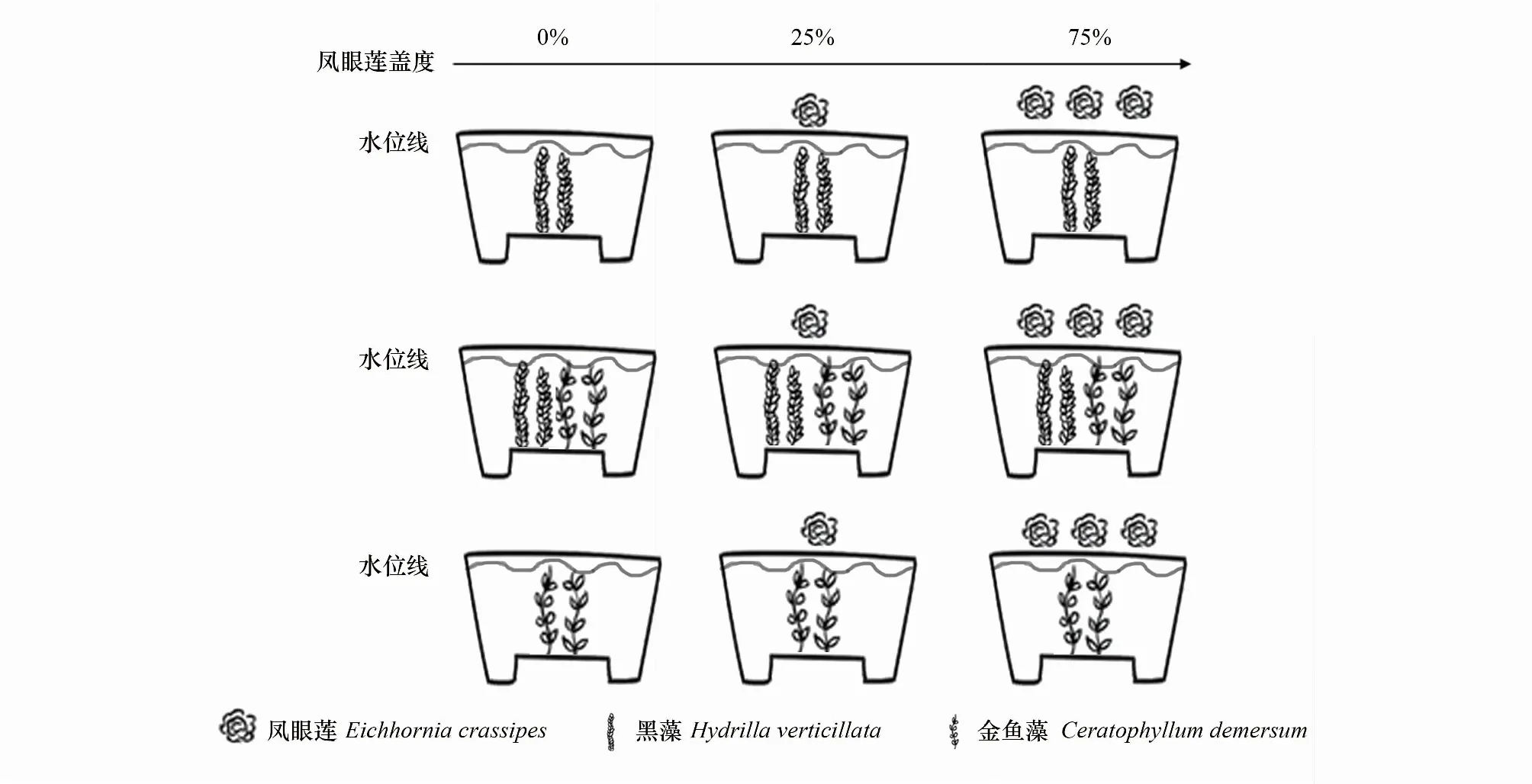
图1 实验设计图Fig.1 Experimental design 实验设计9个处理,每个处理设置5组重复,共45组样品
1.3 数据分析
实验结束后,分别收获入侵植物凤眼莲,及各个处理容器中的沉水植物。沉水植物测量生长指标包括株高,分节数和分枝数。将收获的沉水植物整株放入信封内,于烘箱中70゜C烘至恒重(超过48 h),分别称量各部分生物量并计算总生物量。由于入侵植物凤眼莲主要作为处理环境所用,没有进行细分各个指标。
对于各个实验容器的水体,进行测定水体中的总氮、总磷和水体透明度。其中利用碱性过硫酸钾消解紫外分光光度法测定水体中的总氮含量、利用钼酸铵分光光度法测定水体中总磷含量,利用塞氏盘法测定水体中的透明度。
根据烘干称重得到的单株生物量并采用对数响应比lnRR(ln response ratio)来计算沉水植物的种间竞争系数[26],采用公式如下:
lnRR=ln(B+/B0)
式中,B+为有竞争关系时平均单株生物量,B0为无竞争关系时平均单株生物量。
相对邻体效应指数的数值范围为-1—1,其中值小于0时,表示种间关系中邻株对植物的生长产生抑制作用,值越大表示邻体竞争效应越小;当RNE数值大于0时,则表示种间关系中邻株对植物的生长产生促进作用,值越大表示邻体促进效应越大。
采用双因素方差分析(Two-Way ANOVA)分别检验凤眼莲入侵程度与种植模式及两者交互作用对金鱼藻和黑藻生物量、株高、分枝数和分节数的影响,以及水体中总氮、总磷和透明度的影响。在进行方差分析前,对数据进行正态性检验和方差齐性检验。用单因素方差分析不同处理下相对邻体效应指数处理间是否存在显著性差异。数据统计分析用SPSS 20.0统计软件进行。使用Duncan方法进行显著性检验(P<0.05)。使用结构方程模型(structural equation model, SEM)分析凤眼莲入侵强度以及水体理化性质对沉水植物生长的影响大小,利用SEM的标准回归化系数,即指标间的路径系数量化凤眼莲入侵或水体性质对沉水植物生长的影响程度。在构建初始模型时,假设沉水植物群落、入侵植物群落、水体理化性质任意两个变量之间,及变量的指标之间都会产生相互作用,这样的模型为饱和模型。通过删除不显著的变量关系(P>0.05的路径),或者基于不同变量之间的残差协方差来增加变量关系,对初始模型进行修改。利用χ2(当P>0.05模型拟合较好),拟合指数(CFI; 当CFI>0.95模型拟合较好)和近似的均方根误差(RMSEA; 当RMSEA<0模型拟合较好)评估整体模型。在Sigma Plot 14.0中进行作图。
2 结果与分析
2.1 凤眼莲入侵程度和定植模式对沉水植物金鱼藻生长繁殖的影响
凤眼莲入侵程度对沉水植物金鱼藻生物量、分枝数具有显著影响,定植模式对金鱼藻生物量也具有显著影响,同时两者的交互作用对金鱼藻所有的生长繁殖指标如生物量、株高、分枝数和分节数均具有显著影响(表1)。
凤眼莲不存在时,金鱼藻生物量、株高、分枝数和分节数均为单种模式高于混合模式(图2);随凤眼莲盖度增加,金鱼藻生物量显著减少,株高有所降低,但变化不显著,金鱼藻分枝数则显著增加。同时,随凤眼莲盖度增加,金鱼藻各生长指标如株高、分枝数和分节数先是混合模式高于单种模式,之后趋势相反(图2)。
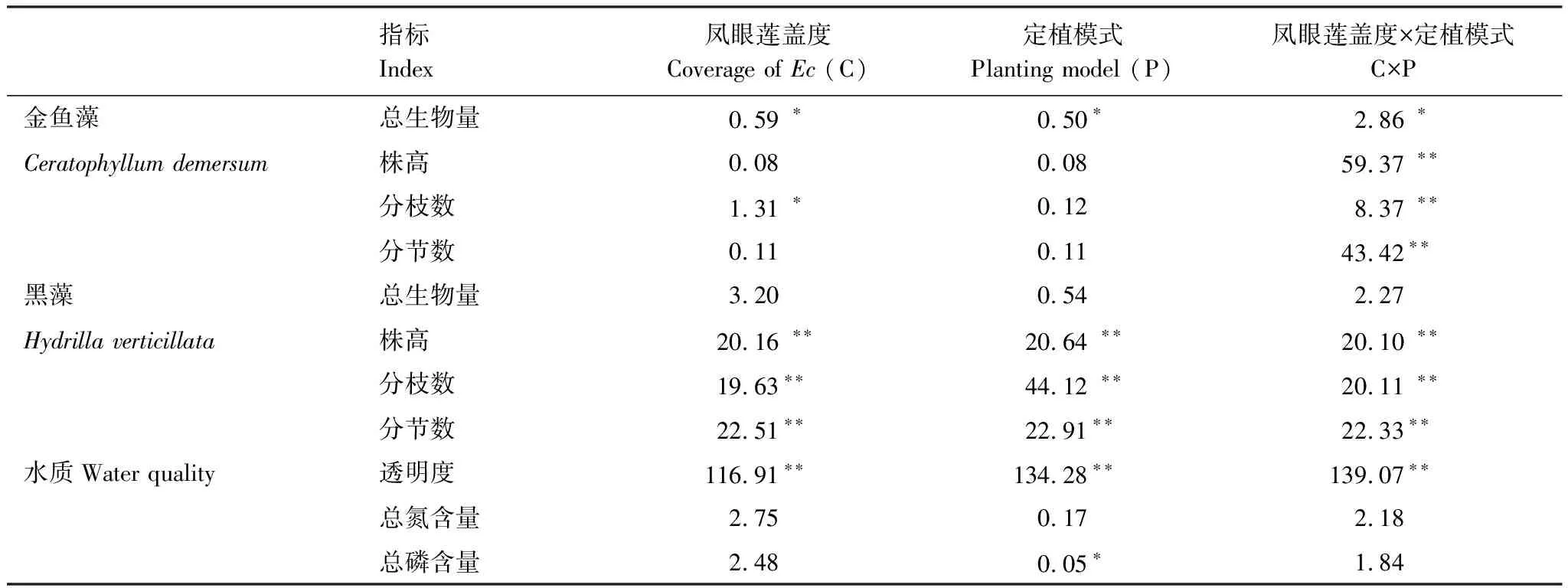
表1 凤眼莲入侵程度和定植模式对沉水植物生长指标及水质影响的方差分析表
2.2 凤眼莲入侵程度和定植模式对沉水植物黑藻生长繁殖的影响
凤眼莲入侵程度、定植模式和两者的交互作用对沉水植物黑藻株高、分枝数和分节数均显著影响,而对生物量无显著影响(表1)。
凤眼莲不存在时,黑藻生物量、株高、分枝数和分节数均为单种模式高于混合模式(图3);随着凤眼莲入侵,但是其盖度较小时,黑藻生物量有所降低,其生物量和株高单种模式小于混合模式,而分节数却趋势相反;随着凤眼莲盖度增加,黑藻的各项生长繁殖指标均降低,且均为单种模式高于混合模式(图3)。
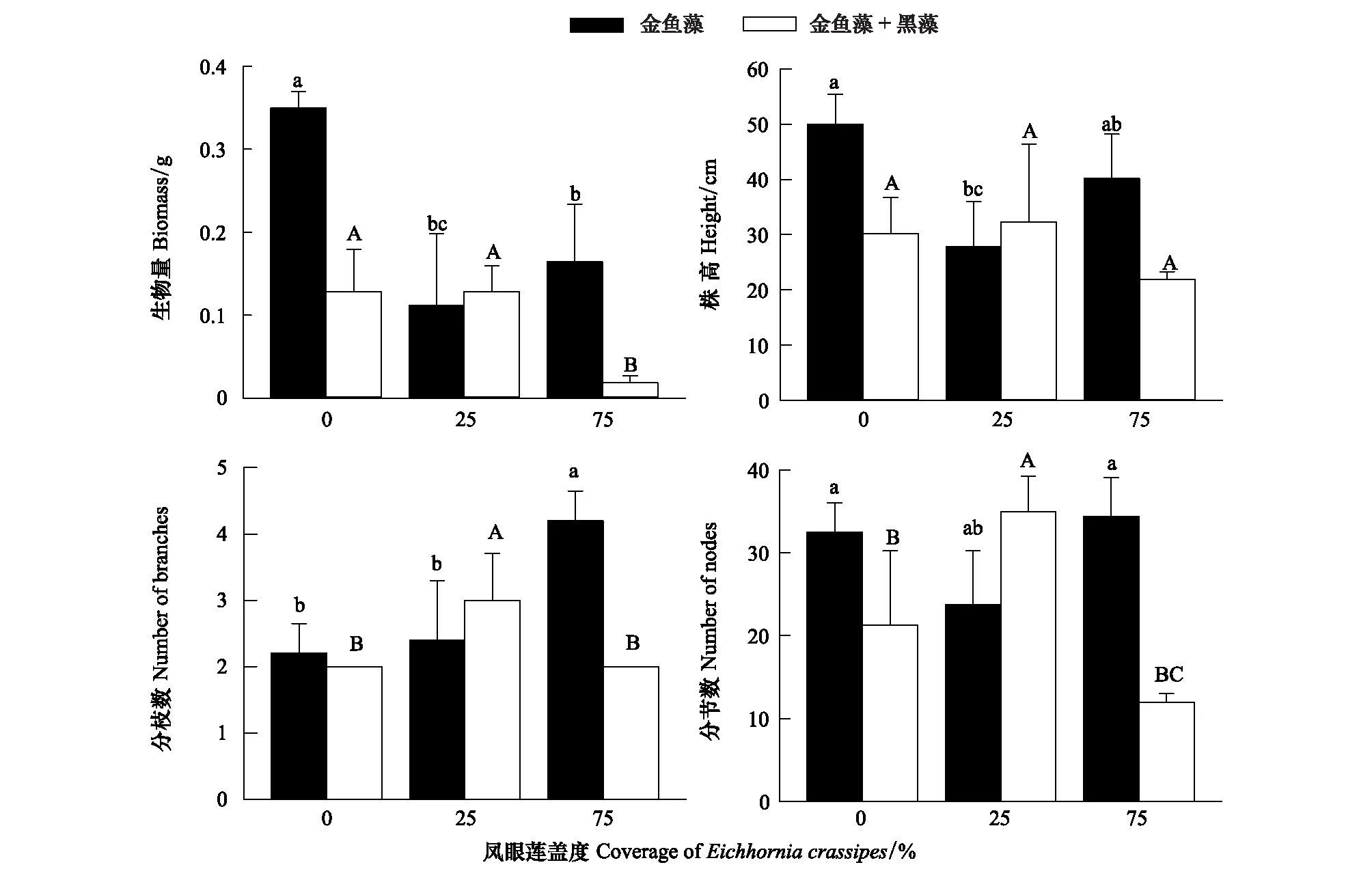
图2 凤眼莲入侵程度和定植方式对金鱼藻生长繁殖指标的影响 (平均值+标准误)Fig.2 Effects of different invasive degree of Eichhornia crassipes on the growth index of Ceratophyllum demersum (Mean+SE) 不同小写或大写字母表示不同入侵程度同一模式下差异显著(P<0.05)
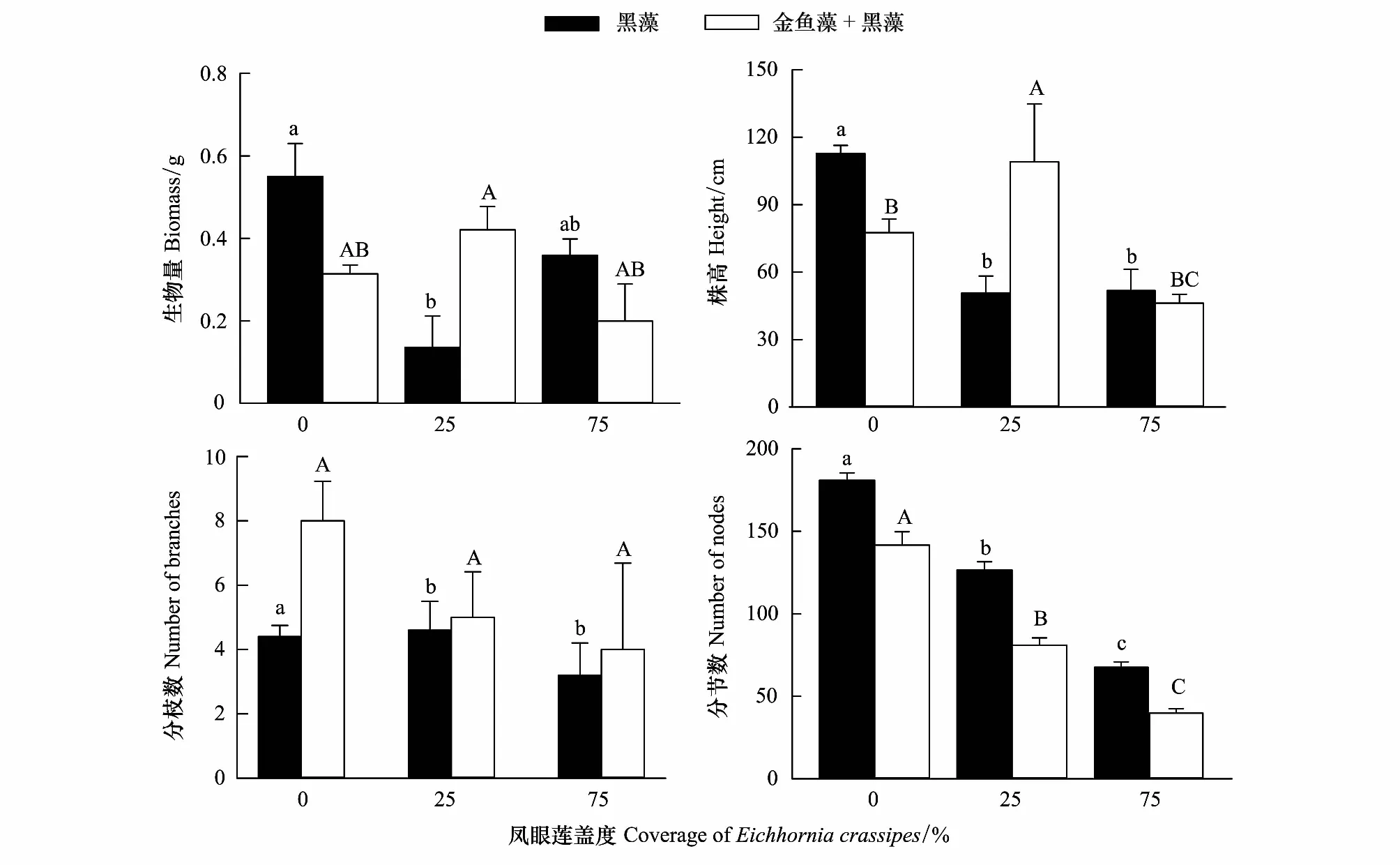
图3 凤眼莲入侵程度和定植方式对黑藻生长繁殖的影响 (平均值+标准误)Fig.3 Effects of different coverage of Eichhornia crassipes on the growth index of Hydrilla verticillate (Mean+SE) 不同小写或大写字母表示不同入侵程度同一模式下差异显著(P<0.05)
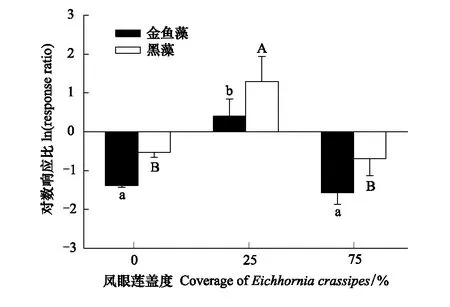
图4 凤眼莲入侵程度对沉水植物种间关系的影响 (平均值+标准误)Fig.4 Effects of different coverage of Eichhornia crassipes on the interspecies relationship of submerged plants (Mean+SE) 不同小写或大写字母表示不同入侵程度同一模式下差异显著(P<0.05)
2.3 凤眼莲入侵程度对金鱼藻和黑藻种间关系的影响
当凤眼莲不存在时,两种沉水植物金鱼藻和黑藻种间关系指数均小于0,说明此时两种沉水植物单种生物量高于混种,种间关系表现为竞争作用;随着凤眼莲入侵,两种沉水植物种间关系指数均大于0,表明其沉水植物单种模式生物量小于混种模式,种间关系表现为促进作用,说明随着入侵胁迫变化,物种间的竞争关系削弱,而转变为促进作用共同抵御其胁迫生境。但随着其凤眼莲入侵程度增加,两者的种间关系指数又变回小于0,表明两者又呈现了竞争作用(图4) 。
2.4 凤眼莲入侵程度和定植模式对水质的影响
凤眼莲入侵程度、定植方式和两者的交互作用均对水体的透明度显著影响,同时定植模式对水体的总磷含量具有显著影响,其余均无显著影响(表1)。
随凤眼莲盖度增加,水体中总氮TN含量无显著变化,总磷TP含量先略微上升后下降,但受凤眼莲入侵程度影响不显著。当凤眼莲不存在时,水体透明度较低;随凤眼莲盖度增加,水体透明度先增加后略微降低,但仍高于没有凤眼莲入侵的对照组,说明凤眼莲对水体透明度有轻度净化作用(图5)。
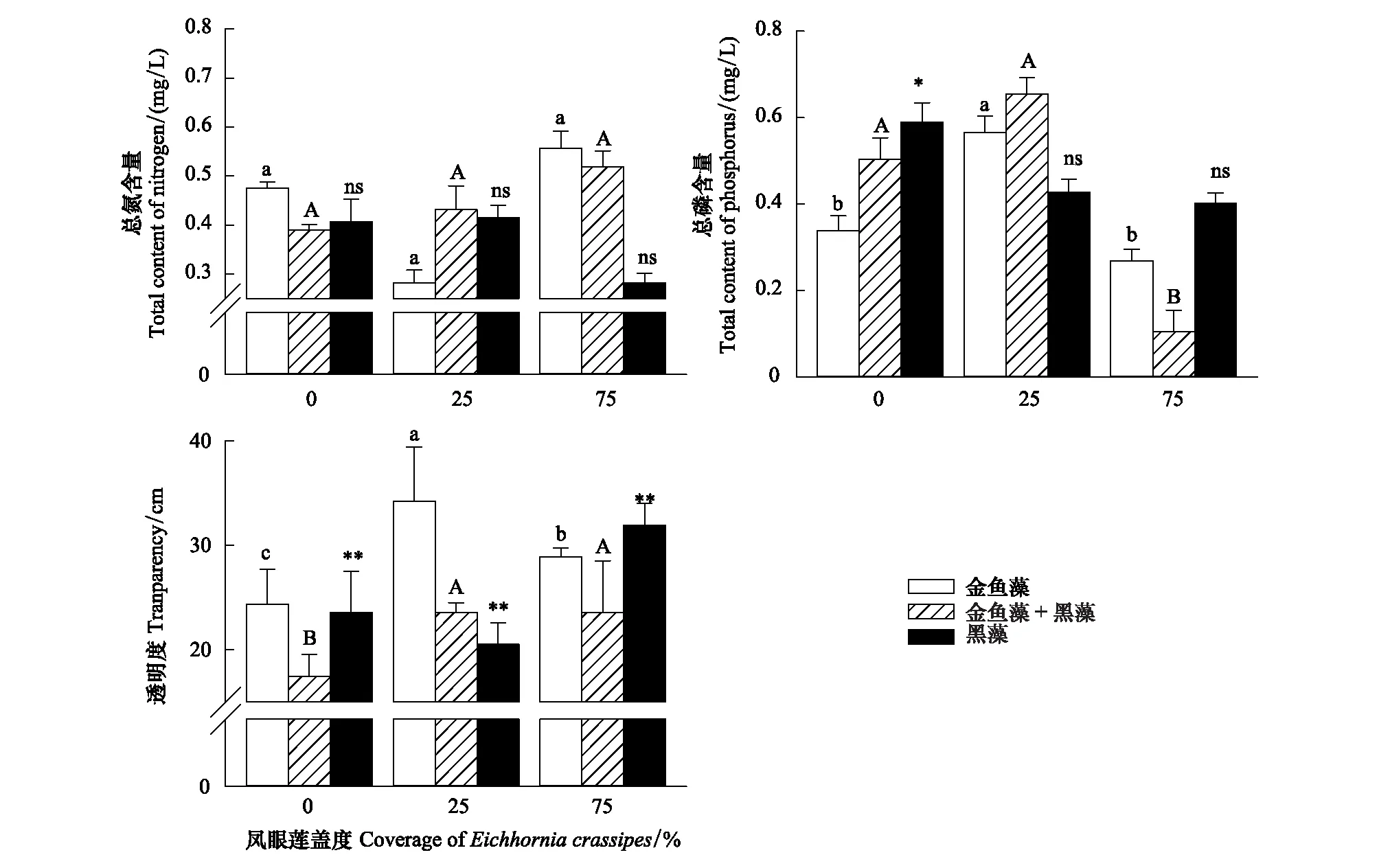
图5 凤眼莲入侵程度和定植方式对水质的影响 (平均值+标准误)Fig.5 Effects of different coverage of Eichhornia crassipes on water quality (Mean+SE) 不同小写或大写字母表示不同入侵程度下同一定植模式差异性显著(P<0.05);ns代表无显著性差异,*,**分别表示显著性差异P<0.05、P<0.01
2.5 凤眼莲入侵和水环境因子对沉水植物生长影响的结构方程模型分析
在相关性分析基础上,选取影响沉水植物群落特征的因素,通过建立模型、拟合模型、评价和修正模型后得到了沉水植物群落与入侵植物群落、水体理化性质相互关系的结构方程模型(图6)(χ2=4.512,CFI=0.98,RMSEA=0.002)。沉水植物群落特征与株高、生物量、分枝数、分节数呈显著正相关,其中与生物量相关系数最高,为0.439;其次为株高,相关系数为0.318。入侵植物群落特征与凤眼莲生物量呈显著正相关,相关系数为0.708。水体理化性质与透明度、TN、TP呈显著正相关,其中与TN相关系数最高,为0.625。植物入侵和TN等水体理化性质增加对沉水植物群落特征均有显著负向影响(P<0.05),路径系数分别为-0.487和-0.657。同时,入侵植物群落与水体理化性质也具有显著的相关关系,相关系数为0.246,说明入侵植物群落与水体理化性质相互影响,并且水体理化性质较入侵植物对沉水植物影响更大。
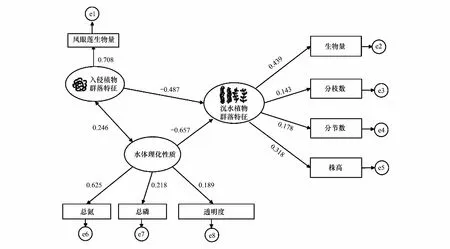
图6 入侵植物群落、沉水植物群落与水体理化性质相互关系的结构方程模型Fig.6 SEM of the relationship between invasive plant community, submerged plant community and the physical and chemical properties of water 椭圆代表潜在变量,长方形代表可测变量,圆代表残差,双箭头表示相关系数,单箭头表示路径系数
3 讨论
3.1 凤眼莲入侵程度和定植模式对沉水植物生长繁殖指标的影响
植物入侵程度的影响主要与植物本身入侵性,及本地物种群落的脆弱性,群落的空间尺度大小等密切相关。研究表明黄顶菊(Flaveriabidentis)的入侵显著降低了植物群落的物种多样性指数和丰富度指数[27],其它研究表明随着反枝苋(Amaranthusretroflexus)入侵程度的增加,本地群落植物的物种数、株高及生物量均显著降低[28];而在加拿大一枝黄花(Solidagocanadensis)中度和重度入侵区域群落生物多样性显著下降,而轻度入侵区域群落多样性则无显著变化[20]。本研究结果与以往研究类似。
入侵植物凤眼莲具有较大的生态位宽度及较强的繁殖能力,对环境的适应能力极强,大量繁殖后可占据极大的空间。较强的资源利用能力使得凤眼莲可以成功入侵本地沉水植物群落并进一步扩散造成危害。夏秋季是凤眼莲和沉水植物的生长旺盛期,在此时段外来物种入侵对本地物种多样性的负作用也相对显著[29]。本实验中凤眼莲的入侵使沉水植物的总生物量及株高明显降低,同时对不同种类的沉水植物影响效果也不同。凤眼莲入侵使金鱼藻生物量降低了约2.4倍,黑藻的生物量则降低了约1.5倍,对金鱼藻生长的抑制作用更加明显。说明黑藻对凤眼莲入侵的耐受能力更强。随着凤眼莲入侵程度增加,金鱼藻的分枝数也随之显著增加,分节数则没有显著变化,说明当入侵威胁加大时,金鱼藻更倾向于横向扩展,以寻求光照适应不利环境(图2)。而黑藻的分枝随凤眼莲盖度的增加略微下降,而分节数则呈现显著下降,说明虽然黑藻对凤眼莲有一定的耐受性,但随着凤眼莲入侵程度增强,凤眼莲入侵将削弱了黑藻的生长繁殖。
比较凤眼莲相同入侵程度下两种沉水植物的生长指标,可以明显看出黑藻的生物量及株高均显著高于金鱼藻,黑藻在群落生长中占据竞争优势。黑藻和金鱼藻均属于冠层型沉水植物,在水体中处于同一生态高度,不存在相互遮阴的可能,出现此现象的原因可能是由于黑藻强大的繁殖能力[30]。黑藻拥有断裂生殖、块茎、具鳞根出条以及种子繁殖四种繁殖方式,其中断裂生殖由于黑藻质地清脆易断,会产生大量断枝,可使黑藻大量繁殖。并且,黑藻无性繁殖体的生根能力也强于金鱼藻[31]。同时,黑藻在地下部分也有一定优势。黑藻的不定根能够迅速扎根在土壤底质中吸收营养,从而影响金鱼藻生长,因此黑藻在竞争中占据优势。
3.2 凤眼莲入侵程度对沉水植物种间关系的影响
在自然界中,物种不是孤立存在的,彼此间的相互作用普遍存在。种间关系是影响植物形态、生长、存活以及物种分布的重要因子,且在具有相似生态幅的物种间往往更容易发生激烈的竞争现象,植株的特征、资源水平和环境条件都会影响其竞争能力。在水生环境中,有效光合辐射和氮磷等营养物质也是影响植物竞争的重要环境因子[32]。而对于凤眼莲入侵会导致其光资源及营养物质等环境条件的变化,进而影响沉水植物的群落生产力,也可能导致其沉水植物的种间关系发生变化。研究表明当凤眼莲入侵显著降低了沉水植物群落的生产力,主要由于其降低了优势种黑藻的生物量[10],进而可能会导致沉水植物种间关系的改变。
无凤眼莲入侵时,金鱼藻与黑藻之间的种间关系均小于0,表现为竞争关系,且金鱼藻受到黑藻的竞争作用更强;随凤眼莲入侵盖度增加,两者的种间关系由先促进作用后又变回竞争作用,表明在受到轻微胁迫时,两者共同协助抵御其不良环境,但是随着胁迫程度加重,两者又呈现竞争关系,黑藻仍呈现较强的竞争作用,符合“胁迫梯度假说 (stress gradient hypothesis, SGH)”[33]。其他研究也表明当处于非胁迫条件时,辣蓼与苔草的地上部分对光的竞争显著;但在胁迫水位条件下,植物间的竞争作用则非常微弱[34]。可能是因为相比于种间或种内的竞争作用,外界的环境胁迫更能影响植物的生长状况[35],进而引起种间关系变化。
3.3 凤眼莲入侵对水质的影响
水体环境对湿地的结构和功能十分重要,其中营养物浓度是重要的环境控制因子,为植物的生长提供能量[36]。水体中营养盐的有效性决定了植物的初级生产量,而过量的营养盐会使水体中藻类以及其它水生生物异常繁殖,引起水体透明度和溶解氧的变化导致水质恶化和生态系统破坏。水体透明度下降时,光的通透量减少,喜光植物生长受到限制,沉水植物受到的影响则更大[37]。
本实验中随凤眼莲盖度增加,水体中营养浓度变化不显著。可能是凤眼莲对水体氮磷的去除主要依靠根系及根系微生物的吸附作用[38]、植株的吸收同化作用、微生物转化作用及沉淀作用[39],而本实验时间处于凤眼莲花期(9—11月),生长速率减缓,根系吸附作用不显著[40]。他人研究发现, 凤眼莲在富营养化的水体中能够使水体的透射光明显下降[41]。而本实验中凤眼莲入侵反而使水体透明度增加,可能是由于凤眼莲并未大量扩散繁殖, 没有在水面上形成草垫遮挡阳光, 净化作用占据主导,使水体更加清澈。植物对水体中氮磷的净化作用是以植物为中心的植物与微生物综合作用的结果,是一个持续进行的过程[42—43],本实验持续时间相对于植物的生长期偏短,只能反映一段时间内凤眼莲对水体中的氮磷去除的作用,具有一定局限性。
3.4 凤眼莲入侵程度和水环境因子对沉水植物生长影响的结构方程模型分析
凤眼莲的入侵程度这一生物因素和总氮、总磷等水体环境非生物因素对沉水植物生长具有不可忽视的影响。多项研究表明,凤眼莲在适宜的生境中迅速繁殖,在水面上方形成细密的植物垫,大大抑制了水下光照,从而限制沉水植物生长[44]。水体透明度是反映水下光照的重要指标,直接影响沉水植物可接受到的光照强度,进而影响光合作用[45]。营养盐浓度也会影响植物的生长,低浓度氮磷能促进沉水植物生长;高浓度氮磷则会导致藻类数量激增,水体透明度降低,不利于正常沉水植物生长。凤眼莲入侵程度和水体的理化性质对沉水植物生长均有显著影响(P<0.05),这与多数前人研究一致[46],而水体理化性质对沉水植物生长的影响略大于入侵植物,推测是因为水体理化性质不仅可以直接影响沉水植物的生长繁殖,还会通过影响入侵植物的生长间接影响沉水植物[47]。将凤眼莲入侵强度对沉水植物生长及种间关系影响制成一个概念框架图(图7)。根据概念框架图,无凤眼莲入侵时,沉水植物间表现为竞争关系;凤眼莲轻度入侵显著降低了两种沉水植物的生长繁殖,沉水植物种间关系表现促进。当凤眼莲重度入侵时,两种沉水植物“各自逃逸”,重新相互竞争。
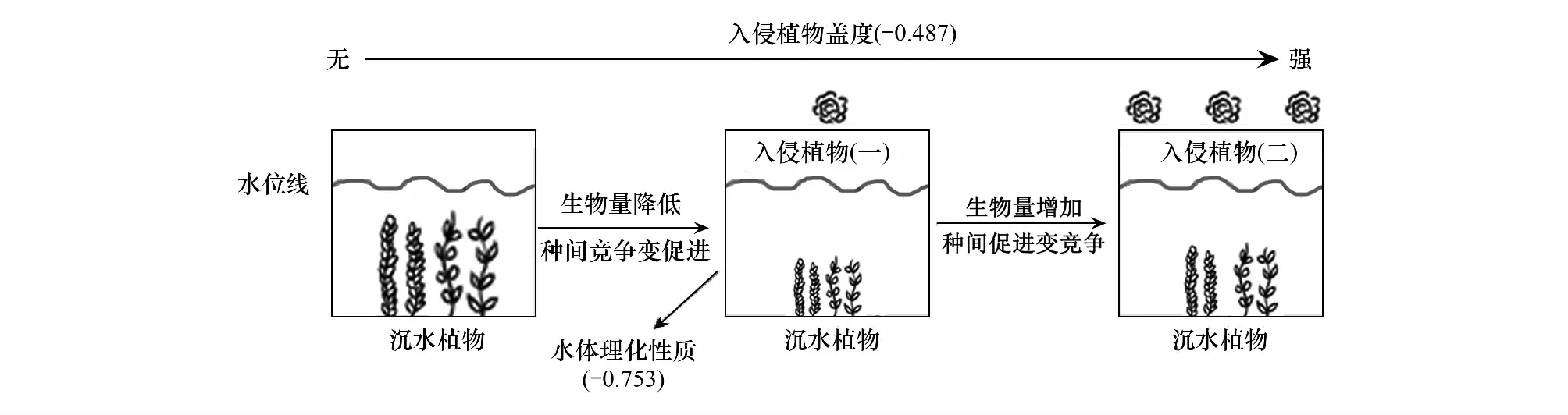
图7 沉水植物生物量及种间关系决定因子示意图Fig.7 Schematic diagram of determining factors of submerged plant biomass and interspecific relationships -0.487和-0.753分别为入侵植物盖度和水体理化性质对沉水植物影响的结构方程模型系数
4 结论
综上所述,凤眼莲的入侵使沉水植物的总生物量及株高均明显降低,金鱼藻生物量降低了约2.4倍,黑藻的生物量则降低了约1.5倍,对金鱼藻生长的抑制作用更加明显;当凤眼莲不存在时,金鱼藻与黑藻之间的种间竞争关系较强;随入侵盖度增加,种间关系则先变为促进作用后又变回竞争作用。黑藻在沉水植物共存环境中占据了明显的优势地位;随凤眼莲盖度入侵程度增加,水体中TN、TP浓度均无显著变化,水体透明度则略微降低,说明凤眼莲对水体透明度有轻度净化作用。植物入侵及TN等水体理化性质增加对沉水植物生长均有显著负向影响,且水体理化性质对沉水植物生长的影响略大于入侵植物。
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
湖泊科学(2021年4期)2021-07-07
生物学通报(2021年7期)2021-07-01
环境科学导刊(2020年6期)2020-12-07
花卉(2020年12期)2020-01-09
当代水产(2019年10期)2019-12-18
——以武汉紫阳湖为例
绿色科技(2019年2期)2019-05-21
蔬菜(2018年5期)2018-05-17
杂草学报(2015年2期)2016-01-04
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
