
石油烃-重金属复合污染共去除研究进展
2022-09-13 13:06:08夏如馨
石油学报(石油加工) 2022年5期
夏如馨, 姜 岩, 彭 蕾
(重庆工商大学 废油资源化技术与装备教育部工程研究中心,重庆 400067)
传统的油污染问题依旧广泛存在且不断加剧。此外,在政策驱使下,以油企为代表的产污企业遗留下来的污染土地发展成为新问题。主要体现在2个方面:其一,随着中国工业化园区的建设,出现了很多企业迁址“入园”事件,旧址土地一般都遭受了较为严重的油污染;其二,在国家严厉的环保政策及行业内部技术升级的驱使和推动下,部分“土炼油”和低产能等企业的关停也遗留了大面积油污染土壤。对于这些土壤的处置研究在以往基本忽略了重金属的干扰,而事实上油污染场地一定伴随着重金属污染。Adeniyi和Afolabi[1]较早关注了二者复合污染问题,发现在两个加油站、两个汽修车间和一个国家电站的污染土壤中,均存在石油烃以及Cd、Cr、Cu、Ni和Pb等5种重金属的混合污染。
发展至今,人们对石油烃与重金属复合污染问题已经有了较为清楚的认知。首先,原油本身就含有重金属,Osuji等[2]及祁鲁梁等[3]对国内外油田原油的灰分进行了定性分析,发现存在Zn、Cu、Pb、Cd、Ni、Mn、Co、V等34种微量金属。这些金属元素与石油烃一起构成了石油的组分,二者相伴相生[4];其次,钻井开采过程中由于钻井液添加剂、低品质重晶石的使用[5],或者与碎钻屑一同从地层中携带出来,从而引入重金属污染。在对钻井废弃物的毒性进行检测时,一些常见的具有危害性的金属几乎全部检测到了踪迹,如Zn、Pb、Cu、Cd、Ni、Hg、Ba、Cr等[6];最后,对于润滑油而言,由于二烷基二硫代磷酸锌(ZDDP)等各类含金属添加剂的使用,以及机械磨损等过程均会混入重金属。同时,部分重金属污染土壤也会伴随着由机加工和其他操作引起的油污染[7]。在对重庆某化工厂的Cr(Ⅵ)污染土壤进行采样分析时发现,除了Cr(Ⅵ)(质量分数580~940 mg/kg)以外,石油烃的质量分数最高达到18760 mg/kg,比一些油污染场地的含油量还高。可见,石油烃和重金属2种污染物经常同时存在于污染环境中,并产生复杂的底物交互作用。不同的重金属由于具有不同的键合度和迁移率而对微生物和环境表现出不同的毒性作用;同一重金属又能以胶体或可溶性络合物等形态存在,从而表现出不同的生物毒性和底物竞争特性,使二者的环境行为发生改变,影响污染物的跨膜运输及生物降解。
以往,对于石油类污染场地中的重金属缺乏足够的重视,生物修复研究以生物除油为主,而屏蔽了重金属的扰动。笔者针对石油烃-重金属复合污染问题,综述了当前复合污染微生物修复研究现状。重点论述了石油烃与重金属的交互作用,剖析了石油烃降解菌在复合污染环境中进化出的不同金属抗性机制,指出在生物修复领域中大量基础研究与实际应用的脱节,提出将重金属的影响和去除纳入到制定油污染场地修复策略中,推动生物修复技术走向实际应用是未来的发展方向。
1 重金属抑制石油烃的生物降解
重金属在土壤环境中呈现出复杂的作用结果,仅浓度变化就会对石油烃的生物降解产生不同影响,不过,抑制作用还是占据主导[8-14]。多数研究结果证实,随着金属浓度的增加,石油烃生物降解效率降低。Oriomah等[11]的研究就具有代表性,Achromobacterxylosoxidans对Cu(Ⅱ)和废机油都表现出耐受性,但当二者共存时,随着Cu(Ⅱ)质量浓度增加至200 mg/L时,A.xylosoxidans对废机油的降解能力由40%降低到5%。Baltrons等[12]也发现了相似的现象,Cr、Co、Pb、Mn、Ni、Cu和Zn等金属浓度越高,对微生物降解3~4环多环芳烃的抑制作用越明显。重金属抑制石油烃生物降解机制更多地表现为损伤细胞或抑制细胞活力[15]。在重金属环境中,细胞主动或被动地吸附和吸收重金属后会影响酶蛋白等生物活性物质的功能特性;细胞还可依靠代谢酶和有机酸等物质在胞内、胞外还原和络合重金属。这些作用将会占用细胞正常的代谢通道,形成与天然底物的竞争,细胞对重金属的应激响应扰动了原有的石油烃代谢途径。因此,重金属对石油烃抑制作用的复杂性体现在对细胞代谢的影响上,其作用结果再抑制石油烃的生物利用。
在现有认知中,除了重金属本身的毒理学效应以外,更多的研究集中在酶蛋白方面。重金属可以通过抑制石油烃降解酶系来影响其代谢通路。Liu等[16]发现在质量浓度100~300 mg/L的Pb(Ⅱ)作用下,Bacillusmalacitensis的邻苯二酚2,3-双加氧酶被诱导,使酶蛋白的空间构象发生改变、催化活性位点被掩蔽,这种作用是通过激发氧自由基的产生而造成酶蛋白损伤;此外,Pb(Ⅱ)的抑制作用也表现在影响细胞骨架的运动从而迟滞细胞的分裂增殖上。加氧酶是多环芳烃(PAHs)类化合物代谢的关键限速酶,在石油烃代谢通路中起到重要作用[17]。当前,对该酶的研究手段已较为成熟,相关报道较多。尽管对金属抑制酶蛋白活性的研究相对较多,但不同的研究体系以及研究角度的不同,对其机制的阐述不尽相同,形成普遍共识的是绝大多数重金属对酶蛋白会产生抑制作用。
此外,石油烃代谢过程中会产生大量水杨酸等有机酸,这一过程是有机物代谢的普遍规律,与重金属并不存在直接关联,但有机酸的累积会降低细胞对重金属逆性胁迫的抗性[18],导致重金属对微生物的抑制作用被放大,最终加剧了对石油烃生物降解的抑制。因此,在以往的生物修复研究中,屏蔽重金属的处理模式不会是最优解决方案。
2 重金属促进石油烃的生物降解
重金属抑制细胞代谢是普遍存在的一种生物毒理现象,不过,重金属的作用并非完全体现在其生物学毒性上,当前的部分研究也发现了其对石油烃降解具有促进作用。个别重金属在适宜浓度等特定环境中对细胞毒性效应小,而且可以调节微生物对石油烃的吸附作用,如通过群体感应来调控生物膜的形成和胞外多糖产生等过程[19]。在抑制与调控两种作用竞争下,使重金属表现出提高石油烃的可生物降解性。表1总结了当前发现的对石油烃降解具有促进作用的重金属。其中,该现象较多地发生在Cu(Ⅱ)上[19-22]。Ma等[20]发现,Cu(Ⅱ)的存在可以促进真菌Acremoniumsp. P0997对蒽的吸附,使蒽在水和菌丝之间的相分配发生改变,当Cu(Ⅱ)质量浓度增加至600 mg/L时,蒽吸附系数从0.75 L/g递增至2.25 L/g,吸附系数与Cu(Ⅱ)浓度呈现正相关。2种细胞吸附行为竞争的结果造成了这一现象。重金属与微生物表面之间存在静电吸引力,而PAHs则以范德华力吸附于菌体表面,显然,2种作用力差异明显,导致重金属优先占据细胞表面的吸附区[23];随着金属阳离子浓度的增加,细胞表面电荷逐渐被中和而呈现出近中性状态,此时疏水性的PAHs更易于被吸附。在重金属占据吸附区的初期,PAHs存在一个吸附抑制过程,只是细胞对PAHs的吸附作用原本就非常有限,其生物降解主要依赖于酶系的生物氧化[24],所以抑制结果并不明显。此外,对于PAHs与重金属的交互作用存在另外一种认知:由于阳离子-π[25]作用力使PAHs倾向于吸附在聚集于细胞表面的重金属上,提高了细胞对PAHs的吸附。不过,这一研究结论仅有个别报道,这种新型分子间作用力对复合污染物的去除机制需要进一步探究。
上述研究存在着一个共性问题,随着金属浓度的增加而呈现出一致的促进作用,但对于抑制浓度未做报道。而实际上,同一金属离子处于不同浓度水平时,很可能表现出不同的生物学效应。赵晓秀等[21]研究发现,当土壤中存在23.7 mg/kg的低浓度质量分数Cu(Ⅱ)时,正十六烷的降解率从不含Cu(Ⅱ)的59.0%提高到66.0%;而当Cu(Ⅱ)的质量分数提高到47.4和237.0 mg/kg时,正十六烷的降解率又分别下降至49.0%和39.0%。这一现象可能由于降解十六烷的优势微生物是Cu(Ⅱ)抗性菌株,能够耐受一定的Cu(Ⅱ)毒性,而非优势的其他土著微生物则是Cu(Ⅱ)敏感性菌株,其活性受到Cu(Ⅱ)抑制,减少了对土壤中生长型底物的竞争,从而有利于优势菌摄取各类营养物质,促进十六烷的降解;但当金属浓度超过阈值水平时,会对所有抗性和敏感性微生物产生各种生理、生化和遗传毒性效应,抑制石油烃的微生物降解。
从表1可见,除了Cu(Ⅱ)以外,Pb(Ⅱ)也对石油烃降解具有促进作用。Liu等[16]发现,当Pb(Ⅱ)的质量浓度从150 mg/L降低到50 mg/L时,B.malacitensis胞外和胞内蛋白质的质量分数分别增加了6.0×104和3.0×104。Pb(Ⅱ)诱导菲的降解基因簇表达,刺激了关键开环酶的释放;同时,低浓度的Pb(Ⅱ)刺激产生的毒物兴奋效应可引起蛋白质含量增加,促进了酶-金属-底物三元复合物的形成,最终提高了菲的降解速率;而当Pb(Ⅱ)过量时,则会与Mg(Ⅱ)、Ca(Ⅱ)等营养元素竞争蛋白质成分中的巯基(-SH),又抑制了酶的活性。对于芽孢杆菌属的研究,笔者所在课题组在近期也发现了一株Bacillustropicus,但该菌仅在Cu(Ⅱ)存在下对润滑油的降解具有一定促进作用,而当另外3种金属Pb(Ⅱ)、Cd(Ⅱ)、Cr(Ⅵ)存在时,均产生了抑制作用,体现了重金属生物学效应的复杂性。另外发现的一种有别于Cu(Ⅱ)和Pb(Ⅱ)的作用机制是在含Cd(Ⅱ)的体系中。Cd(Ⅱ)和芘会胁迫细胞产生过量的活性氧(ROS)来诱导O.radicata[27]产生氧化应激反应,损害细胞活性,但高浓度Cd(Ⅱ)又会诱导抗氧化酶(SOD、CAT、POD)更迅速地分泌,可以强烈抑制由于ROS产生的氧化应激,二者的竞争形成了随金属浓度变化的促进效应,因此,表1中芘的去除率提高了9%。总之,微生物为应对重金属毒性产生的不同抗性机制使复合污染环境中石油烃的降解变得更为复杂,利用和提高重金属积极作用,削弱和抵消其胁迫是生物修复技术得以发展的优选策略。
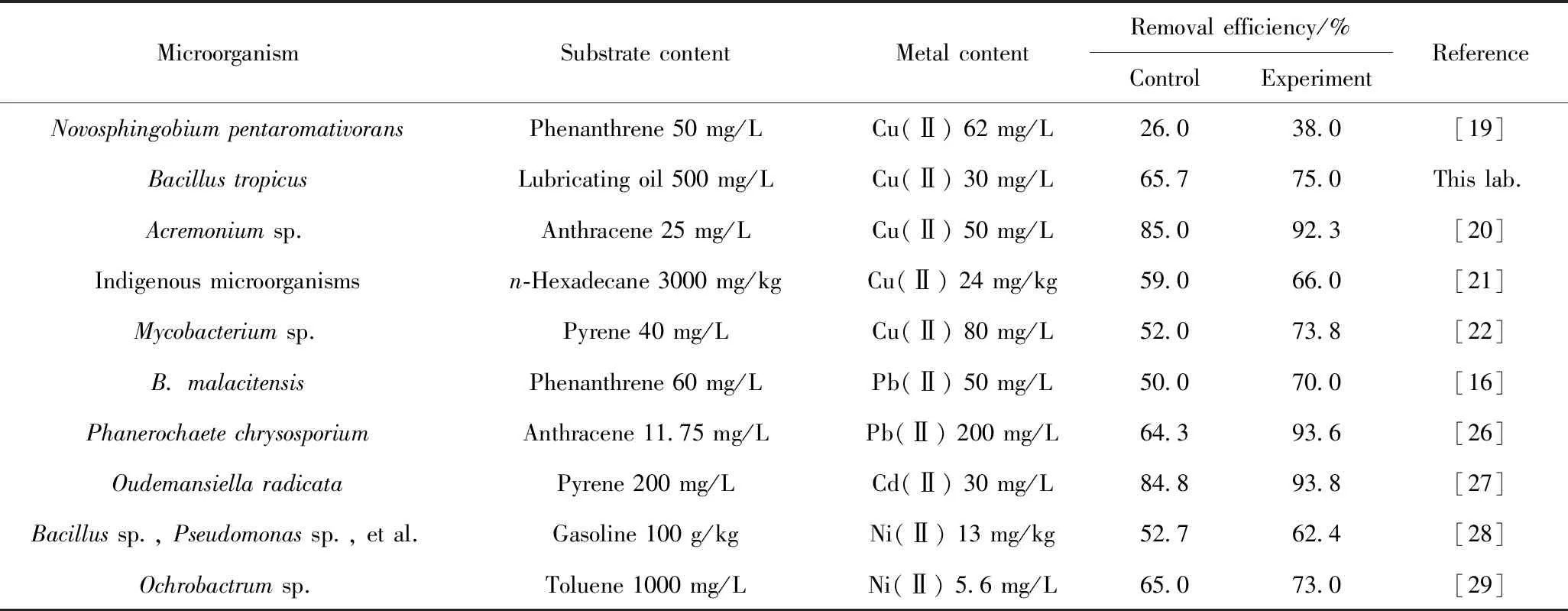
表1 重金属对石油烃降解的促进作用Table 1 The promotion effect of heavy metals on the degradation of petroleum hydrocarbons
3 耐受重金属毒性降解石油烃
油污染场地生物除油研究是当前国内外的研究热点。利用微生物对重金属具有耐受性进行油类污染物的生物降解是诸多报道的研究思路。无论是否考虑重金属的影响,也无论是利用土著微生物还是外源菌,在客观上,只有适应复合污染环境的微生物才能有效代谢石油烃。表2为污染环境中筛选出的耐受重金属石油烃降解菌。在表2中,从油污染场地中选育出的土著微生物普遍具有环境适应性以及石油烃降解能力。这一功能特性的形成被认为与细胞内形成的特有基因有关,即DNA分子中一般都含有抗性基因或降解基因[30-31]。其中,Oyetibo等[32-33]的研究颇具代表性。除Burkholderiacepacia对于原油和蒽都表现出较高的降解能力外,另外2株Pseudomonasaeruginosa和Corynebacteriumkutscheri对难降解的蒽的去除率只有50%左右,其较高的原油去除率则应体现在对其他石油烃组分的降解上。包括该报道在内表2中的各研究均未触及重金属与石油烃的交互作用,但这并不意味着不存在促进或抑制作用,特别是在Oyetibo等的研究体系中,在Ni和Co的存在下,菌株呈现出很高的降解活力原因值得进一步研究。
在实地修复时,可以采用将具有重金属抗性和石油烃降解能力的组合微生物功能菌群应用于复合污染土壤的方法[40]。Alisi等[41-42]利用混合菌群修复了在Mn、Zn、Cd、Cr、Pb多种金属存在下的复合污染土壤,经过15 d的修复,直链碳氢化合物(C12~C20)的降解率为65%~75%,其中,顽固的异戊二烯烷烃类组分(C15~C18)去除率也达到了50%~60%。他们从微生物群落分析入手,发现在重金属和有机毒性胁迫下,外源菌群变得非常活跃,推动整个微生物群落表现出高的代谢活性。在该研究中,外源菌群的功效可能来自于对能量流的特异性捕获,激活整个微生物群落启动生物降解。根据DGGE(变性梯度凝胶电泳)剖面对生态指标的分析,微生物群落的丰度、结构没有明显变化,污染物的胁迫影响主要体现在群落代谢特性上,而非对结构特性产生影响,亦即土壤环境中的微生物多样性并未发生改变。这一报道从污染生态学角度切入,利用Biolog®ECOPlates研究了生物修复过程中污染场地的微生物群落变化,极具特色。但是,研究结论值得推敲。一般认为,环境在遭受污染后,微生物群落的多样性趋于简单化,而微生物的数量在石油烃进入环境后会呈现出逐渐增加的趋势,其原因很简单,有机污染物既提供了碳源等生长因子,也产生了有机毒性,在长期作用下后者导致种群种类减少,前者则刺激了耐受性微生物的生长繁殖。该研究中所得结论可能存在一定局限性,问题出现在实验用的土壤样品。实验用土取自于含有少量Cr、Ni、Cu、Mn等金属的废弃农田,之后复配了柴油和高浓度的Zn、Pb,稳定3 d后开展了外源菌的修复研究。除了前述长期选择作用将导致微生物群落简单化以外,在外源菌的使用上也存在对土著微生物形成竞争性优势的可能性,继而抑制部分耐受微生物的生长和代谢,若这种选择性竞争存在于污染环境中,也会造成微生物种类的减少。
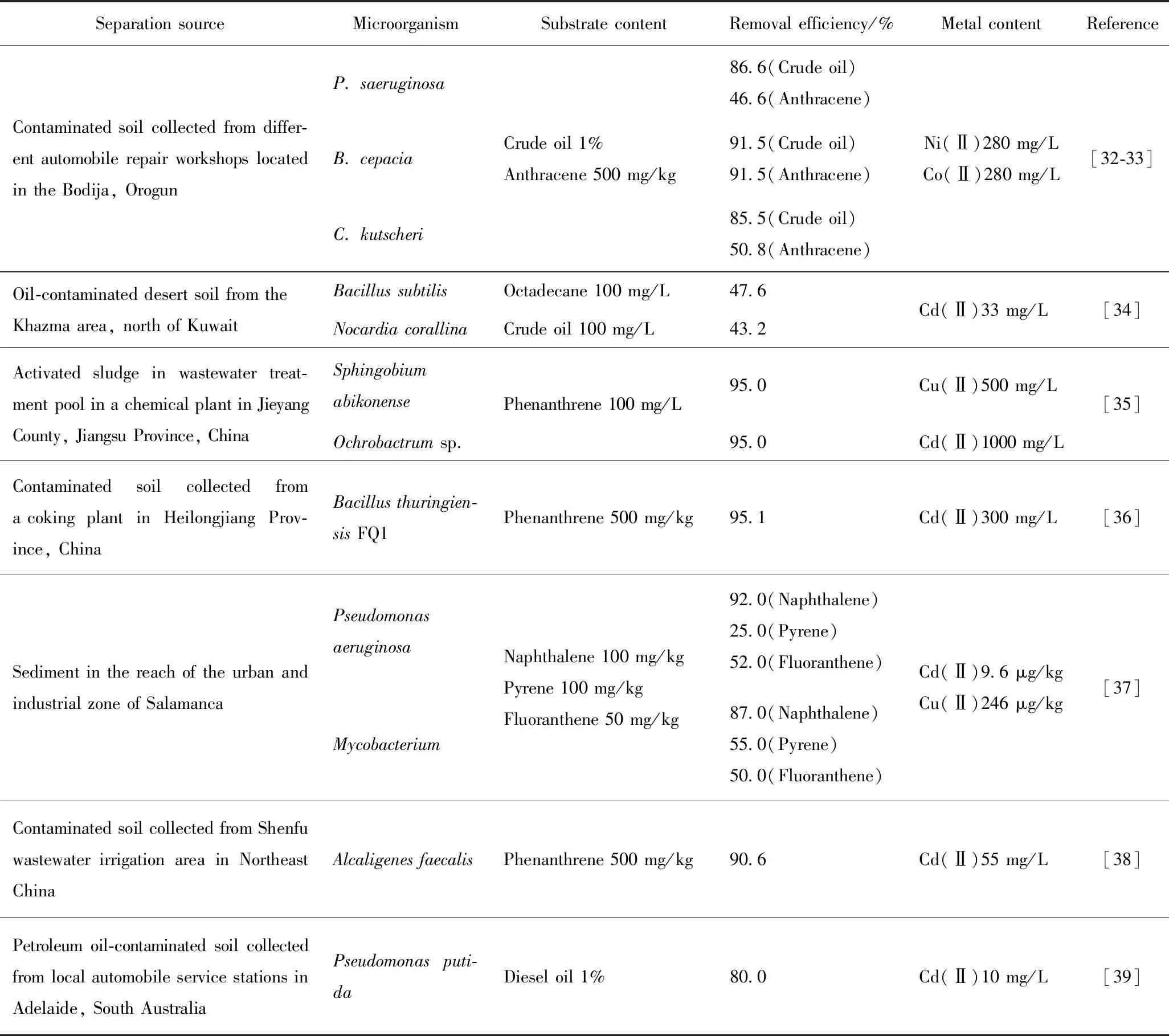
表2 污染环境中筛选出的耐受重金属石油烃降解菌Table 2 Heavy metal-tolerant petroleum hydrocarbon degrading bacteria screened out in polluted environment
无论如何,依靠具有耐受重金属特性的微生物降解石油烃、修复油污染场地是以往的主流研究,并且在大多数报道中均屏蔽了重金属的影响。在笔者看来,利用复合菌群技术处置复合污染场地不失为一种有效手段,其实用性强,符合当前人们的认知水平。
4 石油烃-重金属的共去除
微生物对石油烃与重金属共去除的报道很少,但也逐渐发现Pseudarthrobacterphenanthrenivorans[43]、Sphingomonaspaucimobilis[44]、CitrobacterFreundii[45]、StenotrophomonasMaltophilia[46]、Pseudomonasgessardii[47]等菌株在对重金属的应激反应中进化出相应的防御系统,它们不仅能够耐受重金属毒性,还能在代谢石油烃过程中还原转化和吸附固定重金属,实现了对有机、无机毒性的共去除,如图1所示。其中Cr在环境中多以Cr(Ⅵ)和Cr(Ⅲ)形态存在,依靠石油烃降解菌的代谢活动可以将Cr(Ⅵ)还原为低价态,从而极大削弱铬酸盐生物毒性;与固定作用相比,还原转化作用直接削弱了重金属毒性,这也是抗性生物最主要的解毒机制。对于其他金属元素而言,多以二价等稳定态存在,难以实现电子转移的氧化还原反应,决定了其生物去除只能依靠吸附、吸收、络合等固定作用。当然,固定作用也可以削弱环境污染,但重金属的积累会对细胞生理活性产生消极影响。与此同时,可给态金属离子的减少又缓解了对细胞的胁迫,正是一系列综合作用的结果促成了生物降解石油烃协同去除重金属。
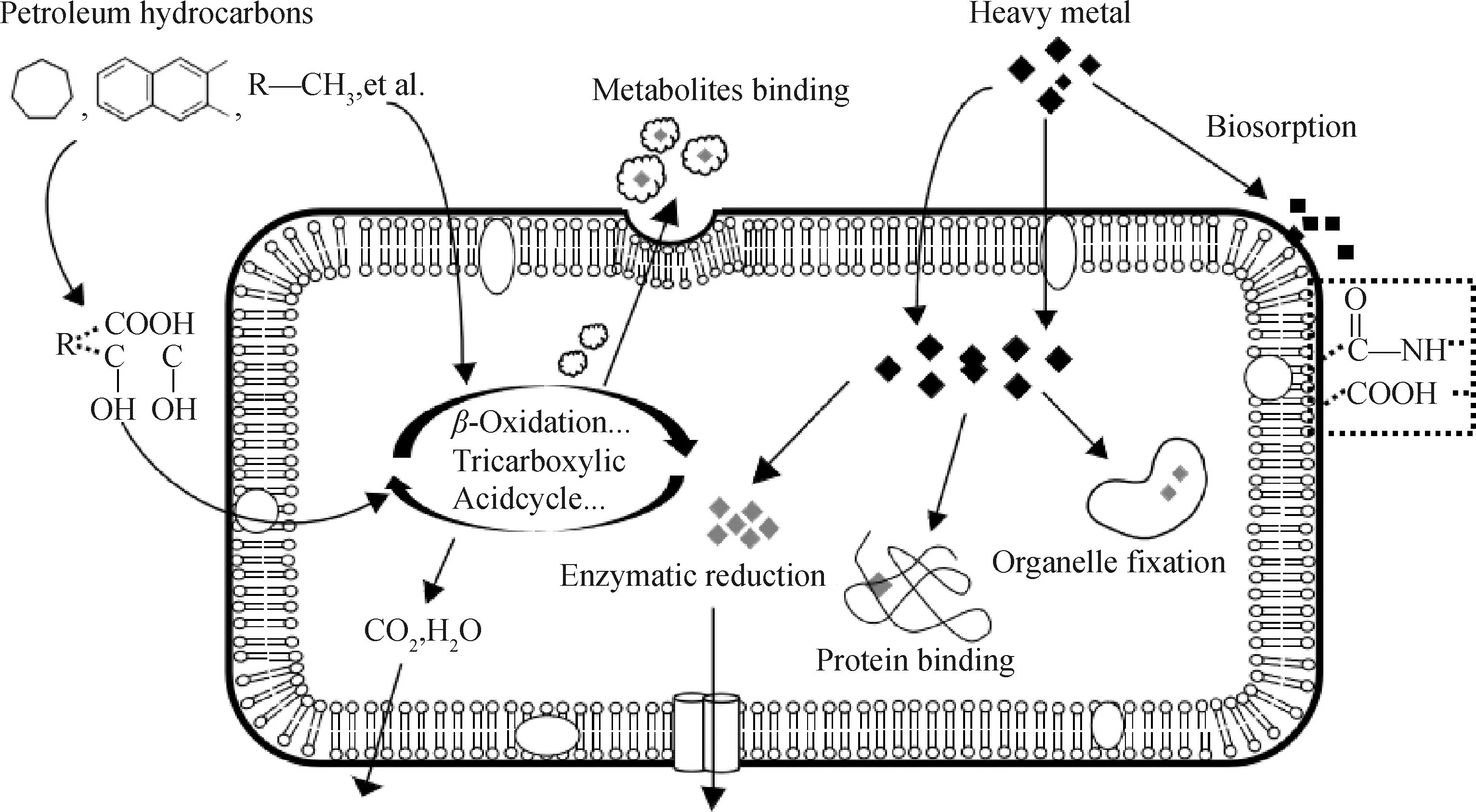
R—Alkly chain图1 微生物对石油烃-重金属共去除作用Fig.1 Microbial co-removal effect on petroleum hydrocarbons and heavy metals
4.1 降解石油烃协同转化重金属
在复合污染环境中,微生物的代谢产物或者石油烃本身可以作为电子供体[48]以实现重金属的还原反应,进而对2种污染物进行解毒。在Huang等[49]对萘和Cr(Ⅵ)的共去除研究中,首次报道了参与萘降解的酶蛋白NahG(水杨酸羟化酶)能够还原Cr(Ⅵ),使PseudomonasbrassicacearumLZ-4可以同时降解6 mmol/L的萘和500 μmol/L的Cr(Ⅵ)。NahG原本是利用FAD(黄素腺嘌呤二核苷酸)作为辅因子将水杨酸转化为儿茶酚,如图2中A、B、C 3条路径所示,这一过程伴随着电子从NADH(还原型辅酶Ⅰ)转移到氧分子上。但当Cr(Ⅵ)存在时,电子转移则遵循了D途径,直接转移到了Cr(Ⅵ)上,从而实现了对铬酸盐的还原。此外,王衍丽[43]发现,rhd是编码PAHs降解酶蛋白的关键基因,而chrA基因则与耐受和去除重金属有关,只有这两个基因同时存在于细胞内才可能发生共去除效应;继而利用IMG/M(The integrated microbial genomes with microbiome samples)的全基因组和宏基因组数据库(Metagenome database)挖掘出13个菌门、20个菌纲、58个菌目、123个菌科、401个菌属和1164个菌种同时具有这两种基因。基于基因挖掘信息,利用一株P.phenanthrenivorans菌证实了在对质量浓度为100 mg/L菲的降解过程中伴随着Cr(Ⅵ)的还原。另外一种研究方法采用了变性梯度凝胶电泳技术,也收到了很好的研究结果。Ibarrolaza等[44]发现,S.paucimobilis对Cr(Ⅵ)形成了明显的聚驱应激,在S.paucimobilis质量浓度低于3.2 mg/L时,Cr(Ⅵ)和质量浓度为200 mg/L的菲可以被完全去除。这些发现对于生物降解石油烃协同转化重金属的研究具有重要意义,毕竟,共去除的核心问题在于污染环境中存在特异性功能型的微生物,这是从事相关研究乃至解决实际问题的物质基础。
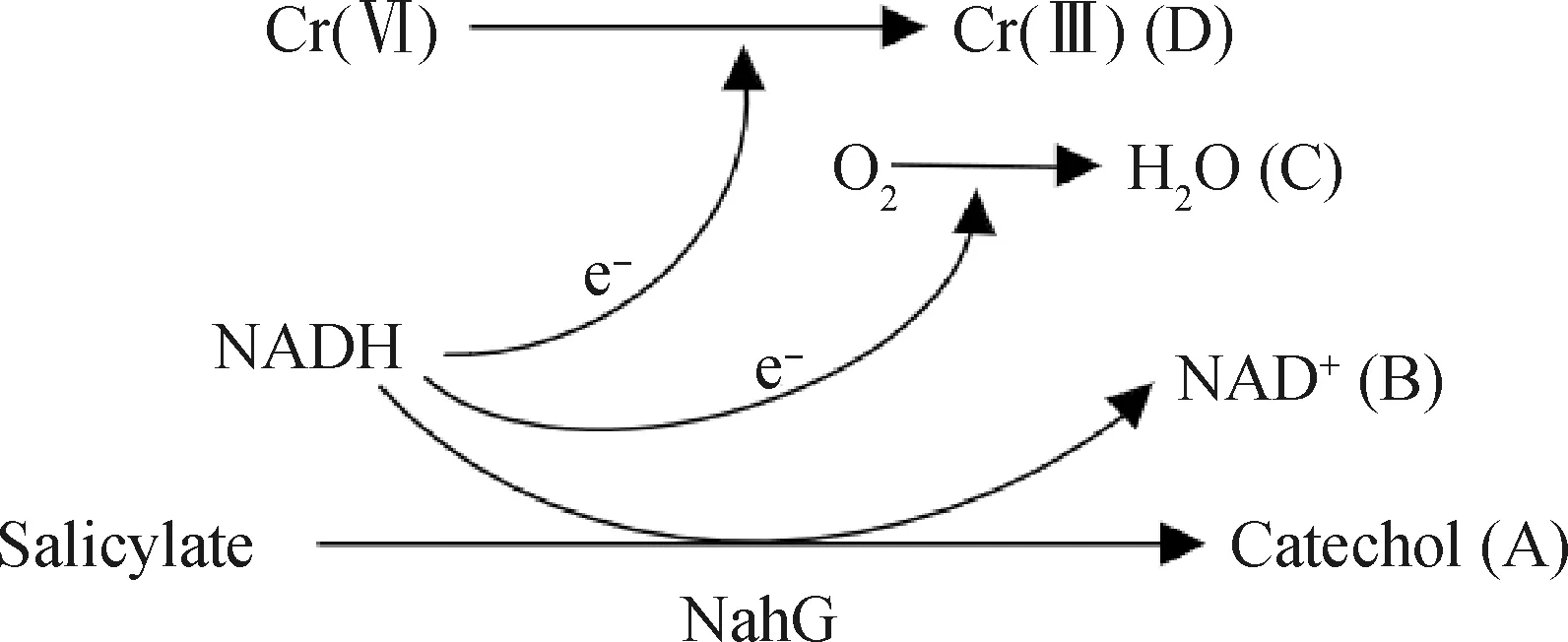
NADH—Reduced form coenzyme;NAD+—Deelectron-reducing coenzyme图2 水杨酸羟化酶(NahG)还原Cr(Ⅵ)过程[49]Fig.2 Salicylate hydroxylase (NahG) reductionCr(Ⅵ) process[49]
4.2 降解石油烃协同固定重金属
石油烃降解菌除了可以通过生长代谢活动改变金属离子的价态,将其转化为低毒或者无毒的状态,还可以通过吸附、络合等作用改变重金属的形态和移动性,从而缓解重金属毒性对生物降解石油烃的胁迫,实现生物降解石油烃同时去除重金属等一系列连锁反应。固定作用可以直接利用细胞表面带负电荷的官能团来吸附游离的正电荷金属离子。孙业政[26]发现在一定浓度范围内石油烃种类不会对P.chrysosporium吸附重金属产生影响,相反在质量浓度为64 mg/L萘存在下,真菌在降解萘的过程中对质量浓度为50 mg/L的Pb(Ⅱ)吸附率提高到75.4%。当前,对于真菌的吸附固定能力高于细菌已经得到越来越多的实验支持。Maddela等[50]就发现与细菌Bacillusthuringiensis和Bacilluscereus对Cu(Ⅱ)的吸附率分别为7.9%和36.3%相比,真菌Geomycessp.对铜的吸附率可以达到97.3%,同时还表现出了对原油的高效降解率。真菌比表面积较大更容易与周围环境发生物理接触,尤其是细胞壁多糖的甘露糖醛酸和古糖醛酸中的高羧基含量提高了对重金属的吸附。这种通过细胞表面官能团吸附固定重金属的报道越来越多,这是细胞固有的特性,当然不局限在复合污染环境中。细胞在吸附重金属后,还可以通过膜表面酶或者借助硫酸盐等离子通道实现跨膜转运至细胞内,被固定在细胞器内,或与细胞内的蛋白质、络合素、多肽结合形成沉淀而被固定。近年来,金属硫蛋白(Metallothionein,MT)在络合重金属方面受到越来越多的关注。这是一种普遍存在于生物体内的金属结合蛋白,其中富含半胱氨酸的短肽,对多种重金属有高度亲和性。如半胱氨酸残基的硫醇基团可以与Cd(Ⅱ)结合形成金属-硫代盐配合物,从而削弱其毒性[51]。这种由重金属诱导产生的具有运输和降低有毒金属作用的蛋白质,对于隔离固定金属离子起到重要作用。
除了上述表面吸附和胞内积累机制以外,微生物在代谢石油烃过程中分泌的铁载体和表面活性剂等代谢产物也可以改变重金属形态[52]。如Fusariumsolani在代谢芘的同时可以产生铁载体类化合物固定Cu、Zn、Cd、Ni、Pb等重金属[53];Pseudomonassp.和Burkholderiasp.等代谢石油烃产生的鼠李糖脂表面活性剂可以螯合Cd(Ⅱ)等[54]。总之,对于石油烃与重金属的共去除作用的认识还非常有限,但这种作用是对污染场地进行生物修复最佳的解决方案,特别在重金属类污染物负荷较高时尤为重要。不过,当前的研究水平尚无法形成稳定的共去除技术,主要存在两个瓶颈问题:其一,缺乏高效的物质基础;其二,成本控制问题。当然,在生物修复领域,环境条件始终是无法回避的现实问题,一定程度地限制了该技术的适用性。笔者所在课题组[55]在石油烃和重金属共去除方面,针对润滑油和Cr(Ⅵ)开发出Lysinibacillusfusiformis功能型菌种资源,可以实现对二者的共去除,但就目前进展来看,要想提高修复效率就要增加助剂成本,而为了控制成本又会牺牲去除率,因此,共去除技术还有很长的路要走,但也的确是最优的解决方案。
5 结 语
建立油污染环境伴随重金属污染的意识具有重要意义。在油污染环境中,重金属也以各种途径进入到污染场地中,形成了石油烃与重金属的复合污染,在实地修复或油污染修复研究中需要统筹考虑。重金属与石油烃间会形成复杂的交互作用,往往并存着诱导促进作用与毒性抑制作用。在多数情况下,重金属的抑制作用占据主导,影响了细胞对石油烃的降解;不过,在部分研究体系中发现,Cu、Cd和Pb等金属也可以通过调节细胞膜的吸附作用或分泌抗氧化酶等机制而对石油烃降解产生积极的促进作用。
以往对于复合污染认知的不足限制了生物修复技术的发展,由于缺乏共去除技术而形成了使用单一污染物处置技术来应对复合污染的局面,而为实现修复目标只能增加修复成本,这种以牺牲处置成本为代价的修复研究只会把生物修复技术带入到没有前途的发展道路。针对复合污染问题,利用特异性石油烃降解菌和重金属抗性菌解决复合污染问题不失为一种切实可行的手段,当然,在生物降解石油烃过程中,能够协同转化、吸附、固定重金属以实现二者的共去除,是最理想的修复策略之一,有助于推动生物修复技术的稳定应用,成为该领域的发展出路。
猜你喜欢
睿士(2023年10期)2023-11-06 14:12:16
小学科学(学生版)(2021年3期)2021-04-13 08:26:18
小哥白尼(趣味科学)(2020年9期)2021-01-18 06:12:42
矿产综合利用(2020年1期)2020-07-24 08:51:42
小哥白尼(野生动物)(2019年5期)2019-08-27 00:53:38
中成药(2018年8期)2018-08-29 01:28:16
Coco薇(2015年5期)2016-03-29 23:14:09
当代化工研究(2016年6期)2016-03-20 16:21:46
中国资源综合利用(2016年3期)2016-01-22 07:28:21
吉林大学学报(医学版)(2015年4期)2015-12-17 07:48:10
