
广西地区籼型陆稻、深水稻与水稻的遗传分化
2022-09-06 10:05:56周衡陵
上海农业学报 2022年4期
王 磊,罗 志,周衡陵,李 静,夏 辉
(上海市农业生物基因中心,上海 201106)
亚洲栽培稻(Oryza sativaL.)(以下简称栽培稻)是世界上最重要的粮食作物之一,为世界一半以上的人口提供口粮。根据农业生境的不同,栽培稻可以分为水稻(Lowland rice)、陆稻(Upland rice)和深水稻(Deep-water rice)[1]。水稻农业生境的典型特点是有蓄水作用的水田,通常有灌溉条件,采取插秧的种植模式。水稻的种植面积占栽培稻种植面积的80%左右,贡献超过90%的稻米产量[1]。陆稻适应旱作的农业生境,通常采取直播、旱作的耕种模式,水分来源主要依赖于降雨。陆稻的种植面积不到栽培稻总面积的15%,贡献大约10%的稻米总产量[1]。深水稻是分布于水域(江河湖泊)附近、适应季节性水淹(深度通常超过50 cm)的一种栽培稻生态型,其种植面积仅占栽培稻种植面积的6%,产量贡献则更低。可见,这3种栽培稻生态型适应不同的农业生境,其中水是主要的差异环境因子。这3种栽培稻生态型在现代育种中发挥了不同的作用:水稻是栽培稻高产、优质的本底与基础;陆稻是节水抗旱基因资源的主要供体,利用陆稻种质资源可以培育出节水抗旱稻[2-3];深水稻则是耐涝、耐淹的优良种质资源,提供耐涝、耐淹基因资源[4]。
对植物不同生态型遗传分化的研究,一直是分子生态学研究的热点[5]。前人对栽培稻不同生态型的遗传分化研究主要集中在水稻与陆稻上[6-10]。Zhang等[6]、Lyu等[7]与Xia等[8]基于分子标记发现水稻与陆稻的遗传分化主要发生在粳亚种内。Xia等[9]进一步研究发现,水稻与陆稻的遗传分化与抗旱性相关,是由于陆稻受到了抗旱性-产量的“双向选择”导致的。Wang等[10]基于全基因组SNP的分析认为,籼稻中也有水稻-陆稻的分化,且与粳稻的水稻-陆稻分化共享部分位点,因此水稻-陆稻分化在籼、粳亚种中是一种“平行进化”的模式。在研究水稻-陆稻分化的基础上,科学家们从陆稻中鉴定出许多抗旱基因[7,11]。目前,人们对于深水稻的关注点只在于它的耐淹性,通过构建试验群体定位、发掘和研究耐淹基因,然后从耐淹基因出发去解释深水稻耐淹的分子机制[12-14]。但关于深水稻与其他不同生态型间遗传分化的研究,目前仍然欠缺。水稻-陆稻分化与节水抗旱性相关,深水稻-水稻分化与耐淹性相关,陆稻-深水稻则是适应两个不同方向导致的分化,通过研究3种生态型的遗传多样性,比较它们两两间的异同,可以更深入理解抗旱性与耐淹性的遗传机制,为今后进一步发掘和研究抗旱、耐淹基因资源,并将其用于节水抗旱稻新品种培育奠定基础。
广西地区是我国栽培稻遗传多样性中心之一,具有丰富的稻种资源[15-17]。广西地区同时具有陆稻、深水稻与水稻3种生态型,给科研工作提供一个很好的条件去研究栽培稻是如何适应不同水分环境,以及相关基因资源是如何在不同的生态型中形成与演化的。本试验以广西来源的籼亚种的陆稻、深水稻与水稻为对象,研究其表型性状的差异与基于全基因组SNP的遗传多样性及两两间的遗传分化,以期揭示3种栽培稻生态型适应性分化的遗传机制,为进一步保护、研究和应用广西栽培稻种质资源提供理论依据。
1 材料与方法
1.1 试验材料
本研究选取来自广西地区的20份水稻地方种、22份陆稻地方种与14份深水稻地方种(表1)。上述材料均为籼亚种,由广西壮族自治区农业科学院水稻研究所提供。
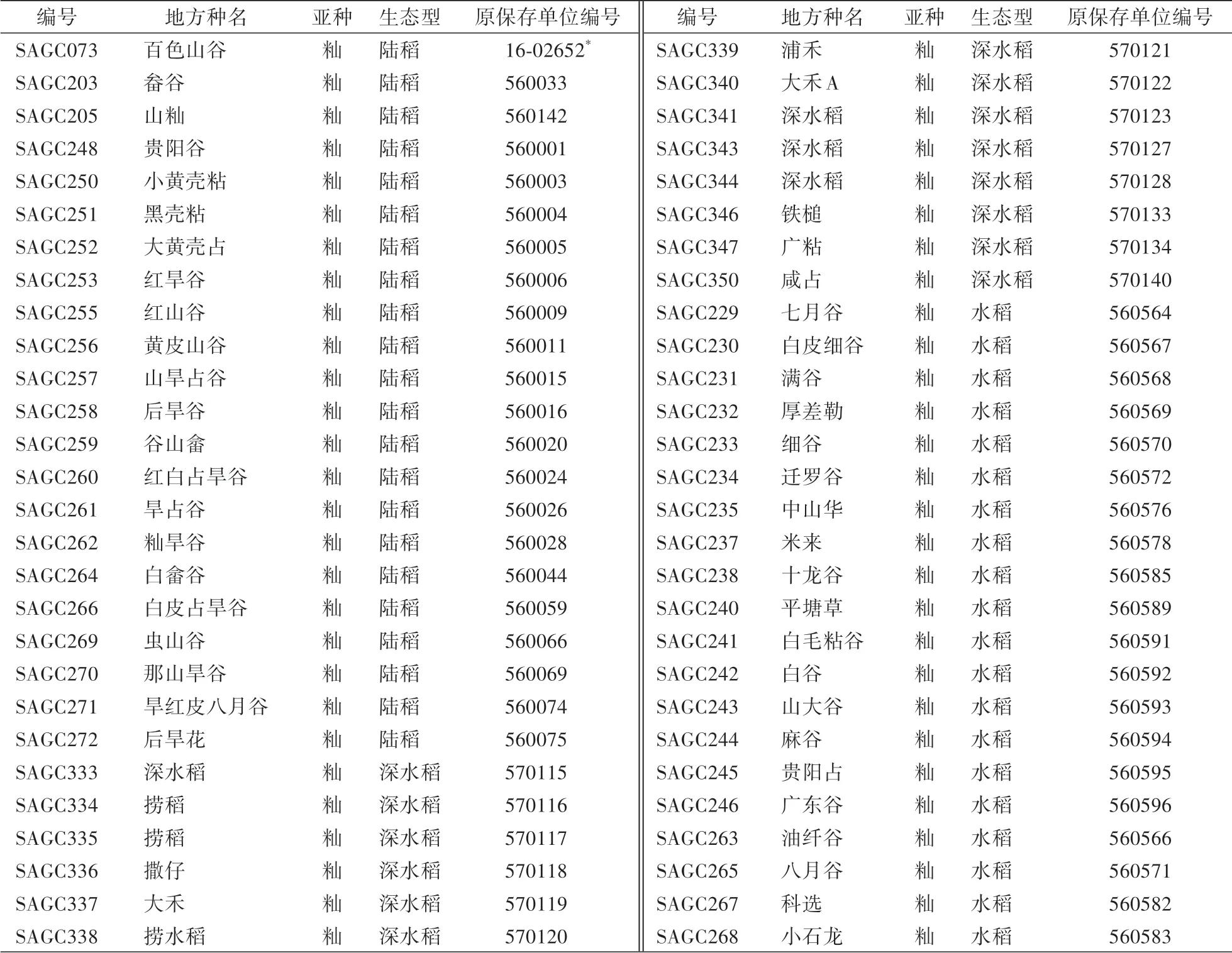
表1 试验所用栽培稻材料及相关信息Table 1 Cultivated rice materials used in the experimen and relevant information
1.2 表型性状测量
首先,测量栽培稻的3个适应性性状,其中中胚轴是一种适应直播的特性,深根比是一种避旱表型,2 h离体叶片失水速率可以反映气孔调节能力。中胚轴的测量参照Wu等[18]的方法,即将浸种2 d的种子置于25℃培养箱中暗培养10 d,然后测量幼苗的中胚轴长度。深根比采用改良的“篮子法”测量[19]。2 h叶片离体失水速率的测量方法:称量刚采下的分蘖期主分蘖顶叶质量W0,将叶片放置在25℃房间内2 h后称取质量W1,然后将叶片置于55℃烘箱烘烤过夜直至恒重时称取质量W2,则2 h离体叶片失水速率=(W0-W1)∕(W0-W2)。测量上述3个表型时,每份材料均设置3个生物学重复。
其次,测量栽培稻的8个重要农艺性状,包括株高、分蘖数、单株产量、生物量、收获指数、谷粒粒长、粒宽与长宽比。田间试验于2014—2015年在上海市农业科学院海南陵水实验基地开展。每份材料种植7行×8列的小区,株间距为20 cm。栽培稻成熟后,每份材料在小区中央选取生长情况较为均匀一致的8个单株测量株高、分蘖数、单株产量、生物量、收获指数,挑选20粒饱满的种子测量粒长、粒宽并计算长宽比。采用多重比较(Student-Newman-Keuls浮标方法)检测各表型参数在不同生态型之间是否存在差异。
1.3 基因组重测序与数据分析
将每份栽培稻材料的种子萌发并培养至3叶期,取3株幼苗混样冻存于-20℃,用于DNA提取与基因组重测序。委托上海欧易生物医学科技有限公司进行测序服务,测序平台为Illumina Hiseq2500。下机的原始数据首先用Fastp v0.6.0(https:∕∕github.com∕OpenGene∕fastp)进行过滤[20];然后利用bwa mem(v0.7.16)将Reads比对到‘日本晴’参考基因组上[21],比对结果经SAMtools(v1.9)转换格式后,利用Picard标记PCR重复;接着,利用GATK version 3.8.1中的HaplotypeCaller得到每个样本的gVCF,通过GenotypeGVCFs得到群体SNP的VCF[22]。这一步骤按照以下指标进行硬过滤:QUAL<30||QD<2.0||MQ<40.0||FS>60.0||SOR>3.0||MQRankSum<-12.5||ReadPosRankSum<-8.0。最后,利用vcftools v0.1.16软件以最小等位基因频率(MAF)>0.05条件对上一步获得的SNP进行二次过滤,获得群体的有效SNP位点。后续所有分析均基于MAF>0.05过滤后的SNP。
首先,比较SNP位点在3种不同生态型间的异同,获得每个生态型特有的SNP位点,并分析每种生态型特有SNP在各自群体内的频率分布。利用plink v1.90软件进行主成分分析[23];利用fastStructure软件以默认参数进行群体结构分析[24];基于Nei的定义[25],计算3种生态型两两间的遗传距离。接着,采用步移滑窗法(窗口100 kb,步移50 kb),通过vcftools软件计算3种生态型两两间的遗传分化指数(FST)及3个群体各自的遗传多样性(π)[26]。将FST值超过95%置信区间的窗口定义为高分化窗口,并根据‘日本晴’IRGSP-1.0参考基因组注释信息(http:∕∕rice.uga.edu∕pub∕data∕Eukaryotic_Projects∕o_sativa∕annotation_dbs∕pseudomolecules∕version_7.0∕all.dir∕)筛选出位于这些窗口内的基因。最后,利用R语言的clusterProfiler包,以粳稻群体(Oryza sativa JaponicaGroup)作为参考(因为只有粳稻的注释数据库)对这些群体间高分化窗口内的基因集进行GO(Gene Ontology)与KEGG(Kyoto Encyclopedia of Genes and Genomes)Pathway的富集分析[27]。
为探索高分化区间与已知抗旱和耐淹相关基因之间的关联,首先筛选出315个已知抗旱和耐淹相关基因,包括302个抗旱相关基因和18个耐淹相关基因(有5个基因与抗旱和耐淹均相关)。按100 kb窗口50 kb步移进行滑窗分割,则每个基因会分布在1—3个窗口中,再对不同基因所分布的相同窗口进行合并统计,可知302个抗旱基因和18个耐淹基因分别分布于590个和37个窗口中。由于以遗传分化指数最高的5%窗口作为高分化区间,如果抗旱基因或耐淹基因与高分化区间没有显著关联,则含有抗旱基因或耐淹基因的高分化区间的预期数量应为590×0.05=29.5个或37×0.05=1.85个。以此预期数量与实际观测到的含有抗旱基因或耐淹基因的高分化区间数量进行比较,即可反映高分化区间与抗旱或耐淹基因的潜在联系。
2 结果与分析
2.1 陆稻、深水稻与水稻的农艺性状差异
由表2可见,在栽培稻3个重要的适应性性状中,陆稻具有最长的中胚轴长度;深水稻具有最大的深根比;在2 h离体叶片失水速率上,3种生态型间没有显著差异。在测得的重要农艺性状中,深水稻具有最多的分蘖,陆稻的分蘖最少;深水稻的株高最小,水稻、陆稻间则没有显著差异;在单株产量、生物量、收获指数与粒长上,3种生态型间均没有显著差异;在粒宽与长宽比上,深水稻的谷粒更宽、长宽比更低。进一步针对11个表型性状进行主成分分析,3种生态型的个体无法通过第一与第二主成分进行区分,提示3种生态型间没有显著的表型分化(图1)。
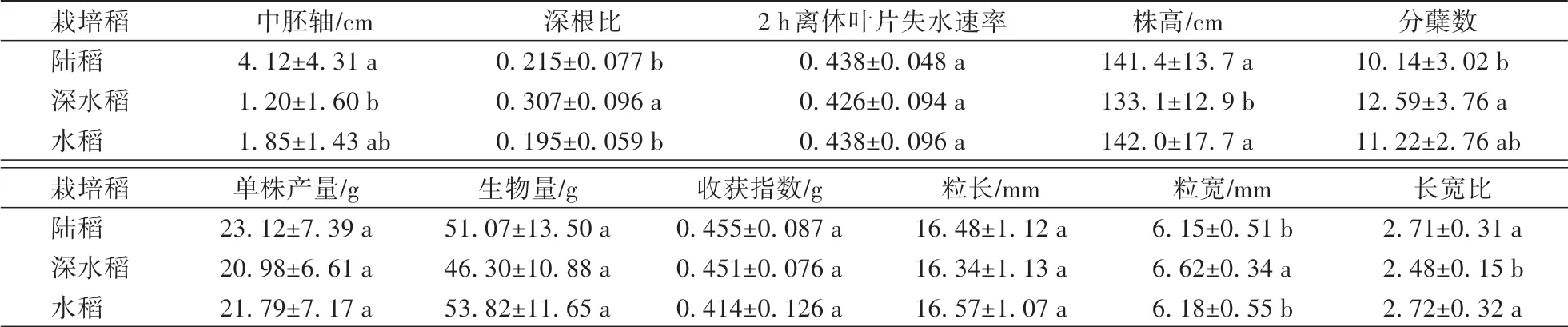
表2 陆稻、深水稻与水稻群体在11个表型性状上的差异Table 2 Differences in 11 phenotypic traits of upland rice,deep-water rice and lowland rice groups
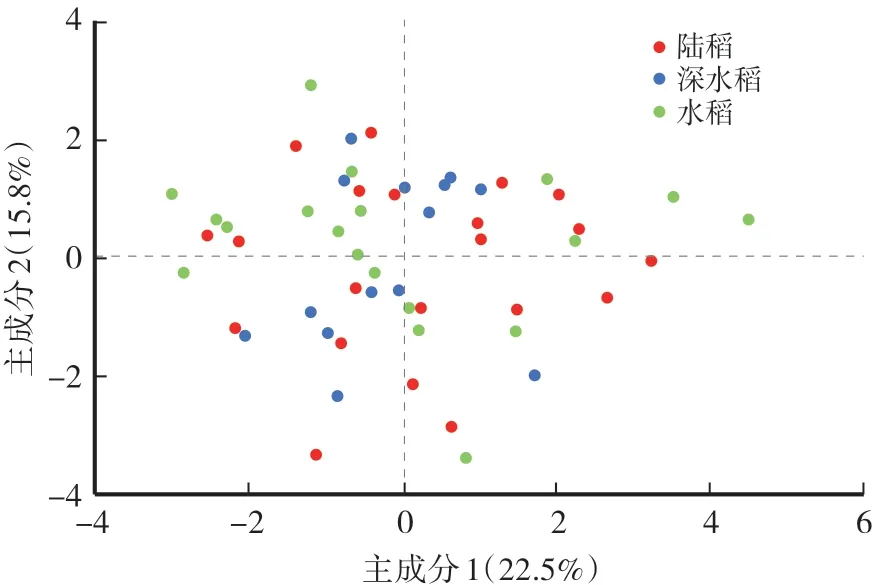
图1 基于11个表型性状的主成分分析Fig.1 Principle component analysis based on 11 phenotypic traits
2.2 陆稻、深水稻与水稻的遗传多样性
基于全基因组重测序,共鉴定出6 007 073个单核苷酸多态性(SNP)位点。经MAF>0.05条件过滤后,共有3 118 328个有效SNP位点。其中,陆稻有11 721个私有SNP位点,深水稻有29 237个私有SNP位点,水稻有35 640个私有SNP位点(图2a)。进一步分析这些私有SNP位点的等位基因频率分布,发现绝大多数陆稻与水稻的私有SNP位点的参考等位基因类型在各自群体内都处于较低的频率(<0.25);而在深水稻中有相当一部分私有SNP位点的参考等位基因类型具有中等水平的频率(0.30<F<0.60)(图2b),提示深水稻的私有SNP位点受到不同于水稻与陆稻生态型的选择。进一步利用滑窗法分析这3个群体的遗传多样性(π),发现三者的遗传多样性总体上相似,其中陆稻的遗传多样性为0.002 17,深水稻的遗传多样性为0.002 35,水稻的遗传多样性为0.002 35。

图2 陆稻、深水稻与水稻群体的SNP位点韦恩图(a)及其私有SNP位点参考等位基因型的频率分布(b)Fig.2 Venn diagram for SNP loci(a)and frequency distribution of reference alleles frequency for private SNP loci(b)of upland rice,deep-water rice and lowland rice groups
2.3 陆稻、深水稻与水稻间的遗传分化
基于有效SNP位点的主成分分析显示,陆稻、深水稻与水稻能够在第一主成分上被区分出来,其中陆稻与深水稻在遗传上更为接近,水稻单独聚集在另一侧(图3a)。这一点也能够在Structure的结果中体现出来:在K=3、4与5的情况下,陆稻与深水稻都具有相似的遗传组分;而水稻的遗传组成比较复杂,几乎具有陆稻与深水稻的全部遗传组分(图3b)。
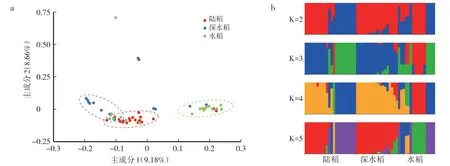
图3 基于SNP的主成分分析(a)与Structure分析(b)的陆稻、深水稻与水稻的群体遗传结构Fig.3 Population genetic structure of upland rice,deep-water rice,and lowland rice groups revealed by principle component analysis(a)and Structure analysis(b)based on SNP
其次,计算了两两生态型间的遗传距离与遗传分化指数。结果显示:3种生态型间在基因组水平上的遗传分化程度均不高,其中深水稻与水稻间的遗传分化程度最高,其遗传分化指数与遗传距离分别为0.049与0.058;而水稻与陆稻间的遗传分化程度最低,其遗传分化指数与遗传距离分别为0.027与0.035(表3)。

表3 陆稻、深水稻与水稻群体间的遗传距离(下部对角数据)与遗传分化指数(FST,上部对角数据)Table 3 Genetic distances(lower triangle data)and genetic differentiation index(FST,upper triangle data)among upland rice,deep-water rice and lowland rice groups
通过滑窗(100 kb窗口,50 kb步移)法,计算了3种生态型两两间的遗传分化程度(FST)与相对遗传多样性(图4a—c),并将具有最高5%FST值的窗口定义为两种生态型间的高分化区间(Highly differentiated region,HDR)。陆稻与深水稻、陆稻与水稻、深水稻与水稻间全基因滑窗相对多态性均值分别为-0.151、-0.137和0.015,但对应的高分化区间滑窗相对多态性均值则分别为-1.027、-0.723和0.257,三组两两比较间均存在显著差异(图4d—f)。这表明高分化区间中群体间遗传多样性差异高于全基因组平均水平。
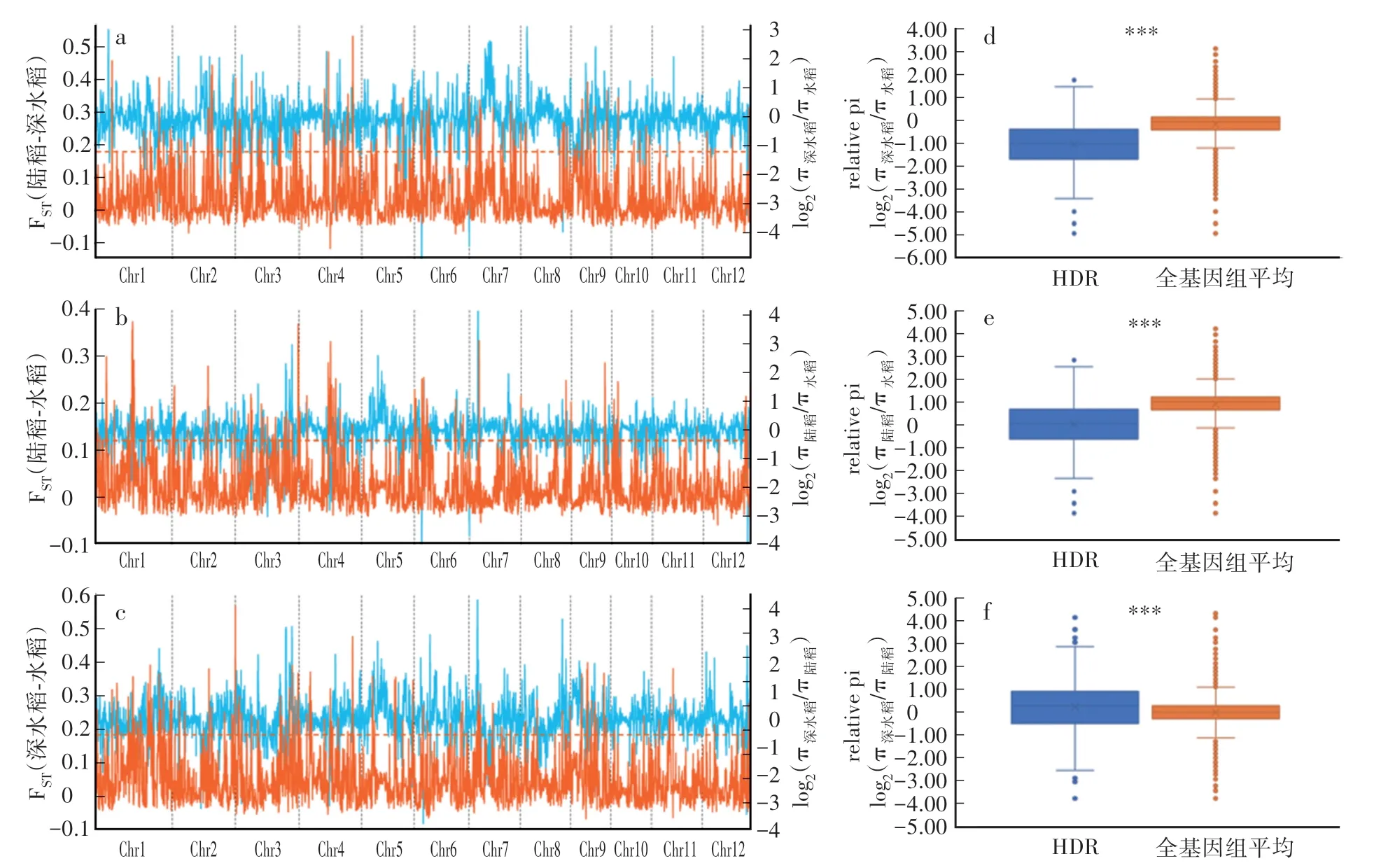
图4 陆稻、深水稻与水稻群体的滑窗遗传多样性及遗传分化指数(FST)Fig.4 Sliding window genetic diversity(π)and genetic differentiation index(FST)for upland rice,deep-water rice and lowland rice groups
进一步比较3个群体间高分化区间的异同(图5a)发现:陆稻-深水稻间分化与深水稻-水稻间分化共享123个高分化区间,占比19.7%;陆稻-深水稻间分化与陆稻-水稻间分化共享41个的高分化区间,占比仅5.80%;陆稻-水稻间分化与深水稻-水稻间分化共享77个高分化区间,占比11.5%。只有3个区间在3种生态型的两两比较中均为高分化区间。
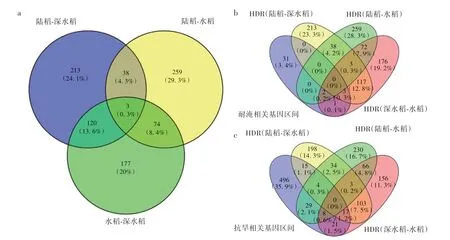
图5 陆稻、深水稻与水稻高分化区间和耐淹抗旱相关基因的韦恩图Fig.5 Venn diagram of highly differentiated regions among upland,deep-water and lowland rice groups and submergence tolerance and drought resistance related genes
接着,对两两生态型间高分化区间的基因进行GO与KEGG pathway的富集分析。结果表明:3种生态型两两间的遗传分化涉及的生物学过程与代谢通路也有较大的差异(图6)。其中,陆稻-深水稻及深水稻-水稻间的遗传分化共享较多相同的生物学过程,如细胞通讯(Cell communication)、信号(Signal)与信号传导(Signal transduction)(图6a,c);但它们也包含了许多特有生物学过程,如陆稻-深水稻分化涉及的定位(Localization)与大分子定位(Macromolecule localization)等(图6a),以及深水稻-水稻分化涉及的阳离子转运(Cation transport)与含氧酸代谢过程(Oxoacid metabolic process)等(图6c)。陆稻-水稻间的遗传分化涉及的生物学过程主要包括初生代谢过程(Primary metabolic process)、有机酸生物合成过程(Organic acid biosynthetic process)、小分子代谢过程(Small molecule biosynthetic process)等与合成、代谢相关的生物学过程(图6b)。KEGG pathway的富集分析表明,陆稻-深水稻间的遗传分化涉及内质网蛋白加工(Protein processing in endoplasmic reticulum)、内吞作用(Endocytosis)等代谢通路(图6d);深水稻-水稻间的遗传分化涉及碳代谢(Carbon metabolism)、精氨酸和脯氨酸代谢(Arginine and proline metabolism)与氧化磷酸化(Oxidative phosphorylation)等代谢通路(图6f);陆稻-水稻间的遗传分化涉及植物MAPK信号途径(MAPK signaling pathway)、氰基氨基酸代谢(Cyano-amino acid metabolism)、果糖和甘露糖代谢(Fructose and mannose metabolism)等代谢通路(图6e)。
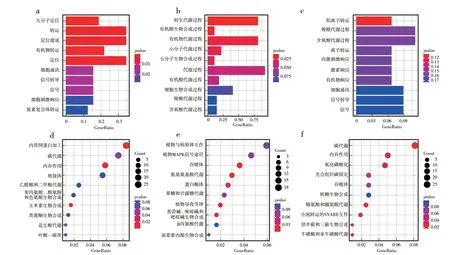
图6 陆稻、深水稻与水稻高分化区间基因的GO和KEGG分析Fig.6 GO and KEGG analysis for genes located in highly differentiated regions among upland rice,deep-water rice and lowland rice groups
2.4 陆稻、深水稻与水稻间的遗传分化与其生境适应性相关
陆稻需要适应依赖降雨的旱作生境,因此具有较好的节水抗旱性;深水稻需要适应季节性水涝,因此具有较好的耐淹性。首先计算了包含耐淹基因或抗旱基因的窗口在两两生态型间的FST,并将其与基因组均值比较,发现耐淹基因所在窗口的深水稻-水稻间FST显著高于基因组均值(图7a),表明耐淹基因在深水稻与水稻间的分化中受到选择。而抗旱基因所在窗口任意两两生态型间FST的均值都与其对应的基因组均值没有显著差异(图7b)。这可能是由于抗旱是一个微效多基因调控的复杂性状,单一基因的效应较低,因此不易受到选择。
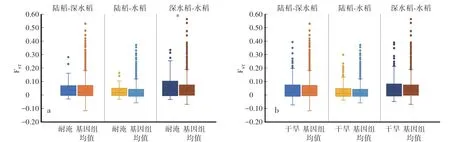
图7 陆稻、深水稻与水稻间包含耐淹和抗旱基因区间FST与全基因组FST的比较Fig.7 Comparisons between genome-wide FST and FST of regions containing submergence tolerance and drought resistance related genes of upland rice,deep-water rice,and lowland rice groups
其次,分析了已知抗旱基因与耐淹基因在陆稻、水稻与深水稻3种生态型两两间高分化区域的分布,用于研究两两生态型间的分化与抗旱性或耐淹性之间的关联。在共18个已知耐淹基因中,有4个基因(包括OsCBL10、sd1、Sub1B、Sub1C)分布在深水稻-水稻间的6个高分化区域(图5b),超过随机预期(随机预期为1.85个高分化区域)。另外,在共302个已知抗旱基因中,有24个基因分布在陆稻-水稻间的41个高分化区间(图5c),超过随机预期(随机预期为29.5个高分化区域)。有意思的是,已知抗旱基因在深水稻-水稻间高分化区域的分布也是超过随机预期的:有32个基因分布在46个高分化区域(图5c)。
3 讨论
栽培稻不同生态型间的适应性分化是一个受到颇多关注的科学问题,因为不同生态型间的适应性分化往往涉及许多重要适应性性状及其相关的遗传基础。广西是我国栽培稻资源多样性中心之一,具有丰富的稻种资源,包含了陆稻、深水稻与水稻3种生态型。这3种生态型分别适应不同的农业生境,各自具备相应的适应性性状,如陆稻适应旱作,具有较好的节水抗旱性;深水稻需要应对水涝浮水生长,具有较好的耐淹性。通过研究广西地区籼亚种内陆稻、深水稻与水稻间的遗传多样性与遗传分化,有助于理解这3种生态型适应各自农业生境的遗传机制,为后续保护、研究和利用这3种生态型种质资源及其包含的遗传资源提供理论指导。
本研究发现,无论是基于表型数据还是基于全基因组SNP,籼亚种内的陆稻、深水稻与水稻间的总体分化程度较低。这3种生态型两两之间的遗传分化指数仅为0.027—0.049,与前人报道的结果相似[6,8,28]。但在基于SNP的主成分分析中,却能在第一主成分上将这3种生态型大致区分开来,提示这3种生态型间存在一定程度的遗传分化。另外,无论是主成分分析还是群体结构分析,结果都表明陆稻与深水稻总体上具有更相似的遗传组成,而水稻群体则具有一定的混杂性。
进一步分析表明,虽然3种生态型间总体的遗传分化程度不高,但依然可以鉴定出许多高分化位点,部分位点的遗传分化指数可以超过0.30。这3种生态型两两间的遗传分化涉及不同的位点,表明栽培稻适应不同农业生境需要不同的遗传机制,这在GO与KEGG富集结果中也有所体现。当然,也应当注意到,不同两两生态型间也会共享比例不同的高分化区间,这也给发掘、研究和应用不同生态型基因资源提供了一些重要启示:(1)陆稻-深水稻间分化与深水稻-水稻间分化共享最高比例的高分化区间,反映出深水稻在适应自身农业生境过程中获得的遗传特性与水稻、陆稻间均存在较大差异。(2)陆稻-深水稻间分化与陆稻-水稻间分化共享最少的高分化区间,表明陆稻-水稻及陆稻-深水稻间的差异不仅仅只涉及旱作与水种的差异,否则两组群体分化应当共享更高比例的位点;反之,共享的41个高分化区间,应是陆稻适应旱作而有别于另外两种生态型的关键区域。(3)陆稻-水稻间分化与深水稻-水稻间分化共享了多达77个高分化位点,提示陆稻相对于水稻的旱作适应与深水稻相对于水稻的耐淹适应,在遗传机制上有一定的共通性。抗旱与耐淹在机制上有一定相通性的典型例子就是水稻sd1与Sub1A基因,它们在水稻抗旱与耐淹中均发挥了重要作用[14,29]。另外,sd1(LOC_Os01g66100)也在本研究中同属于水稻-陆稻分化与水稻-深水稻高分区间。有意思的是,在节水抗旱稻育种实践中,同样观察到类似的现象,如节水抗旱稻品种‘旱优73’既节水抗旱,有很好的旱作适应性[30],又具备良好的耐淹性,在水淹十几天后仍能正常生长并维持一定产量(https:∕∕export.shobserver.com∕baijiahao∕html∕301336.html)。
最后,与预期的一样,在深水稻-水稻高分化区间内找到了一些已知耐淹基因,且耐淹性基因所在区间的遗传分化指数也显著高于基因组均值,因此深水稻-水稻的遗传分化可能与深水稻浮水生长适应季节性水涝相关。与之相似的,在陆稻-水稻高分化区间内也鉴定出超过预期数量的耐旱基因,提示与粳稻的水稻-陆稻分化一样,籼稻的水稻-陆稻分化可能同样与陆稻适应旱作从而使其与水稻形成抗旱性差异相关[8-9]。这与Wang等[10]所揭示的籼稻与粳稻的水稻-陆稻分化属于平行进化的结论相仿。本研究表明:广西籼型陆稻与深水稻资源蕴藏有丰富的抗旱基因与耐淹基因,特别是在相应的高分化区间内,对这些高分化区间的进一步深入研究将有助于节水抗旱稻的改良,以获得具有更强抗逆性状的节水抗旱稻新品种。
猜你喜欢
中学数学研究(广东)(2023年9期)2023-06-03 03:32:40
水利水电快报(2023年2期)2023-02-21 02:34:54
中学生数理化·八年级物理人教版(2022年9期)2022-10-24 07:03:48
现代装饰(2022年5期)2022-10-13 08:47:08
今日农业(2021年14期)2021-11-25 23:57:29
今日农业(2020年13期)2020-12-15 09:08:51
现代装饰(2020年8期)2020-08-24 08:23:52
现代园艺(2018年2期)2018-03-15 08:00:04
河南水利年鉴(2017年0期)2017-05-19 02:29:16
新农业(2016年23期)2016-08-16 03:38:14
