
陆稻的起源与适应性进化研究进展
2022-09-06 10:05周衡陵李朝阳罗利军
上海农业学报 2022年4期
罗 志,周衡陵,李 静,李朝阳,王 磊,罗利军,夏 辉∗
(1上海市农业生物基因中心,上海 201106;2华中农业大学,武汉 430070)
亚洲栽培稻(Oryza sativaL.)是最重要的粮食作物之一,为全世界一半以上的人口提供主食。亚洲栽培稻具有丰富的遗传多样性,在适应不同农业生境的过程中逐渐演化出不同的生态型。根据土壤-水分条件及耕作方式的不同,可以大致将栽培稻分为灌溉稻(Irrigated rice)、低地稻(Rainfed lowland rice)、陆稻(Rainfed upland rice)、深水稻(Deep-water rice)4种生态型[1]。其中,灌溉稻种植于具备灌溉条件,土壤相对肥沃,并且具有成型田埂和犁底层的水田中;雨养低地稻虽然依赖降雨,但同样利用成型田埂与犁底层来保持田间水分,在生育期的大部分时期都与灌溉稻有相似的土壤-水分条件。因此,通常将灌溉稻与低地稻归类为水稻。目前,水稻在全球的种植面积占栽培稻种植面积的80%以上,贡献了超过90%的稻米产量[2]。陆稻适应旱作农业生境,水分来源主要依赖于降雨,通常采取直播旱作的耕种模式,与小麦、玉米等旱作作物相似。陆稻广泛分布于亚洲、非洲和拉丁美洲,在主要耕作地区亚洲,栽培稻仍以水稻为主,虽然在拉美和非洲地区以陆稻种植为主,但相对于全球的稻作规模,陆稻占比相当小。根据国际水稻研究所的最新调查,全球范围内,陆稻的种植面积仅占种植面积的约8%,贡献约4%的稻米总产量[2]。水稻与陆稻的农业生境具有较大的差异,在长期适应各自农业生境的过程中逐步分化(表1)。水稻在较为适宜的水、肥条件与人类精耕细作下,获得了较高的产量与较好的品质;而陆稻所处的农业生境通常干旱、贫瘠、劳动力缺乏,在长期适应的过程中积累了大量节水抗旱、耐贫瘠、适应轻简栽培的绿色性状。不过,陆稻的产量较低,仅为1 000 kg∕hm2左右[2],在产量为导向的栽培稻生产中长期得不到重视。虽然我国是陆稻的发源地与多样性中心,但传统陆稻种植区域的陆稻面积呈快速缩减的趋势[3]。

表1 水稻和陆稻的农业生境差异及陆稻适应性表型Table 1 Differences in agricultural habitats between lowland rice and upland rice and adaptive phenotype of upland rice
过去几十年,随着水稻理论研究进展、育种技术进步及大量水、肥、农药的投入,水稻在实验田中的产量潜力已高达18 000 kg∕hm2以上。但水稻这种高投入、高产出的生产模式,也导致了诸多的问题,使其在未来的发展遇到极大的挑战。这些问题包括:(1)水稻的高产潜力依赖于大量水、肥与劳动力的投入,在条件欠缺的中低产田水稻的平均产量与高产田可相差30%以上;我国有大约70%的稻田为中低产田[4],提高中低产田的产量,是提高我国粮食总产量,解决粮食安全问题的关键。(2)水稻生产消耗大量的水资源,占农业用水总量的一半以上[5],而我国是世界上水资源最匮乏的国家之一,淡水资源仅占世界的6%,但我国灌溉用水却需要用于生产世界25%的粮食,预估2015—2030年淡水的增量难以满足目前粮食增产的需求[6]。在2012年印发的《国务院关于实行最严格水资源管理制度的意见》(国发〔2012〕3号)中指出,截至2030年,力争将国内年用水量控制在7 000亿m3以内,因此,节水农业是保障未来粮食稳产、增产的发展方向;同时,水稻对干旱十分敏感,我国每年有超过60%的水田会受到不同程度的干旱危害,造成减产甚至绝收[7]。提升水稻节水抗旱能力,使其能够应对全球气候变化带来的干旱缺水问题,是现在水稻育种的重要目标之一。(3)水稻淹水种植的方式会产生许多生态问题,包括稻田温室气体排放,传统稻田的甲烷排放占农业系统总排放量的11%[8],灌溉在大量水分流失的同时也带来了严重的面源污染[5]。只有改变水稻传统淹水种植的模式,才能在较大程度上解决上述农业生态风险问题。
为了解决上述水稻生产中存在的问题,实现环境友好、资源节约的水稻生产可持续发展模式,上海市农业生物基因中心率先提出发展节水抗旱稻(Water-saving and drought-resistance rice,WDR)这一理念[7],并培育出了适应轻简栽培、具备较高节水抗旱能力的节水抗旱稻品种[9]。“节水抗旱稻”的核心思想就是利用常规育种将陆稻生态型中普遍存在的节水抗旱、耐直播、耐贫瘠等绿色遗传资源通过高强度的“抗旱-高产”交替选择的育种模式导入到高产优质的水稻中,培育出适应轻简栽培、在常规条件下高产优质且能在轻中度干旱胁迫下保持较高产量与品质的新型栽培稻品种,在国内外受到广泛的认可与关注[9]。发展节水抗旱稻的重要基础工作就是科学研究和利用陆稻节水抗旱遗传资源,而陆稻节水抗旱遗传资源的起源与进化是其中最根本的科学问题,揭示陆稻抗旱性起源与进化的遗传机制,对于挖掘、研究和利用陆稻抗旱基因资源,发展节水抗旱稻育种技术有着重要的理论指导意义。本文就水稻抗旱性起源与进化的研究进展进行综述。
1 陆稻起源的研究
陆稻作为一种重要的栽培稻生态型存在已有几千年的历史,却很少有人探讨其起源问题。长期以来,水稻从野生稻进化,然后迁移到旱地环境形成陆稻生态型的观念深入人心。这种观点最有力的依据在于水稻的考古证据(如水田遗址和相关农具)可以追溯到至少8 000年前的长江中下游地区[10-12],而陆稻最早的稻作遗存距今仅4 000年左右[10,13]。
然而,这种水稻先驯化的观点仍然值得商榷,理由如下:(1)在栽培稻驯化的早期,农耕条件极为落后,稻田不具备灌溉和蓄水条件,早期的栽培稻需要面对季节性干旱;(2)虽然一般认为野生稻的生境是沼泽环境,仍不可避免会遭受季节性干旱,野生稻也有适应季节性干旱的遗传多样性,陆稻有直接从野生稻驯化而来的遗传基础[14];(3)最原始的栽培方式更可能是直播,而陆稻具有直播特性;(4)近年来,特别是基于高通量测序的群体遗传学研究更多地关注亚洲栽培稻的起源和驯化历史,提出我国珠江流域才是粳稻的起源地[15]。而华南地区正是我国陆稻分布的多样性中心,且史籍记载表明华南地区的原始稻作始于陆稻。云南地区栽培稻遗传多样性极其丰富,且该地区的野生稻在遗传与生理上均与现代栽培稻最为接近,长久以来被一批学者认为是栽培稻可能的起源中心[16-17]。
本课题组在近期的工作中,通过对大规模具有代表性的水稻、陆稻地方品种进行群体遗传学研究,发现在栽培稻驯化历史中,野生稻-陆稻-水稻的假说在遗传上是可能的。(1)陆稻的遗传多样性高于水稻,这与水稻是基本型的假设相悖:因为如果陆稻是从水稻中分化出来,必然会经历一个遗传瓶颈,导致遗传多样性较大程度下降。(2)陆稻与野生稻的遗传关系更近,陆稻与野生稻共享更多的私有等位基因。在陆稻与野生稻之间并不存在显著基因流的情况下,只有陆稻直接从野生稻中驯化,才能解释这一现象。(3)促使陆稻与水稻发生适应性分化的驱动力以陆稻中对抗旱性和产量的平衡选择为主,而非定向选择,这与陆稻由水稻分化去适应旱作农业生境的假设相悖[18]。在后续的研究中,本课题组通过基于溯祖理论的SMC++及基于频谱统计的fastsimcoal2方案进行了种群发生历史的动态推演,得到陆稻更早地从野生稻驯化,然后水稻从陆稻分化出来的结果。另外,通过对控制根系、氮素吸收利用等关键适应性性状基因的单倍型分析,也发现其中有很大比例的基因,其陆稻优势单倍型直接或间接来源于野生稻,而非来源于水稻;相反,大多数基因的水稻优势单倍型都能在陆稻中找到。这也支持了陆稻直接从野生稻分化出来的假设,并且说明了陆稻遗传资源直接来源于野生稻。
与此同时,来自纽约大学的Gutaker等[19]研究人员基于群体遗传学、考古学及气象学模型提出了粳亚种生态型分化模式:粳稻最早在大约9 000年前以热带粳稻的形式起源于中国东部长江和黄河流域之间;大约在4 200年前的一次全球性降温事件导致热带粳稻向更温暖的西南方迁移;而在粳稻起源地(长江-黄河流域之间)则演化出了更适应低温的温带粳稻。在Gutaker等[19]的研究体系中,粳稻亚种内的生态型分化主要是热带粳稻与温带粳稻的分化。但从现有的资源来看,(亚)热带粳稻主要是典型陆稻,而温带粳稻多为水稻,且两者在节水抗旱[18,20]、耐直播[21]、耐贫瘠与养分高效[22]等方面与水稻、陆稻农业生境适应相关的性状上有显著差异。因此,Gutaker等[19]所说的(亚)热带粳稻就是陆稻,之后分化的温带粳稻就是水稻。Gutaker等[19]对于粳稻驯化历史的解释与本课题组关于陆稻先起源的假说是吻合的。当然,要证实这个猜想,还得依赖于进一步的考古遗存及古DNA等证据。
2 陆稻与水稻的适应性分化
2.1 陆稻与水稻的表型差异
旱作适应性是陆稻与水稻首要的区别。水稻的种植环境具有良好的蓄水能力或者灌溉条件,整个生育期遭遇干旱胁迫的概率较低。而陆稻一般种植于排水通气良好的土壤中,采取旱作模式,在没有田埂和犁底层保水的条件下,自然降水的不稳定导致陆稻在全生育期均有遭受干旱的风险。为了抵御干旱胁迫,陆稻通常具有较好的节水抗旱性。陆稻的节水抗旱能力表现在:
(1)陆稻具有更深、更发达的根系。根系是植株吸收水分的主要器官,旱地作物需要更深的根系来吸收深层土壤的水分。典型陆稻相较于水稻而言,具有更粗壮且向下发展的根系以及更高的根冠比,这使陆稻在遭遇干旱时能从土壤中吸收更多的水分[18,20,23]。例如,Lou等[20]研究发现,陆稻具有较高的深根性,即陆稻的根系主要是向下发展;Zhao等[23]研究发现,陆稻具有较粗、较为发达的根系,并鉴定出多个与根系发育相关的基因。
(2)陆稻叶片保持水分的能力更好。在增加水分吸收的同时,陆稻也发展出一系列适应性机制以保持较低的水分散失和消耗。叶片是植物散失水分的主要部位。陆稻叶片具有较低的气孔密度和较厚的蜡质层,从而减少了水分散失[24-25]。此外,叶片形态在植物适应环境的过程中会发生有规律的演化,陆稻相较于水稻有较长、较宽的叶片[18]。有研究显示,水稻叶片的长宽与干旱下的叶片卷曲有一定的相关性[26]。
(3)陆稻的分蘖较少。更少的分蘖和叶片同样能够降低陆稻由蒸腾作用导致的水分散失,被认为是陆稻适应旱作的一种机制。有意思的是,作为补偿机制,陆稻往往拥有较大的穗和较大的籽粒从而弥补部分由分蘖减少导致的产量损失[27]。
除了节水抗旱性以外,陆稻还在耐直播特性(中胚轴长度)[18,21]、耐贫瘠与养分高效[28]等方面与水稻产生了适应性分化。因此,陆稻种质资源拥有丰富的绿色基因资源,在目前逐步展开的倡导资源节约、环境友好、效率优先的水稻生产变革中,是重要的绿色种质资源。
2.2 陆稻与水稻的遗传分化
Ono[29]在1973年追溯了日本陆稻的起源,认为日本陆稻是水稻适应旱作以后演化出的新生态型,但遗传上与水稻基本没有太大差异。之后有学者利用同工酶标记发现了日本水稻、陆稻群体特异性的位点[30-31]。利用这些分子标记可以较为准确地鉴定和区别水稻或陆稻种质资源[31]。可惜的是,上述研究所用到的水稻、陆稻品种较少,开发的分子标记数量不足,也没有进一步从进化生物学的角度解释两者的遗传分化。随着分子标记技术的发展,Yu等[32]利用42对RAPD(Randomly amplified polymorphic DNA)标记研究水稻、陆稻的遗传分化,但聚类结果仅仅是将14份试验材料分成籼稻与粳稻两个组。我国学者也在2009年利用36对SSR标记研究包含超过2 100份水稻和800份陆稻地方种的遗传多样性与遗传结构,发现仅有7.58%的遗传变异存在于不同生态型中,却有36.65%的遗传变异是由籼、粳分化造成的,因此认为水稻、陆稻的整体遗传分化水平较低[33]。2014年,Lyu等[34]基于基因组SNP标记对水稻与陆稻的遗传分化进行了研究,同样认为水、陆稻的遗传分化仅出现在粳亚种中;而籼稻遗传背景则较为复杂,可能存在多种起源。上述研究表明,水稻、陆稻在基因组水平的遗传分化程度并不高。
分子生态学的观点认为,环境适应导致的同一物种不同生态型间的遗传分化往往不会覆盖全基因组,而是在与环境适应相关的位点及与之连锁的区域因差异化的选择产生分化,从而在基因组上形成不连续分布的“分化岛”[35]。因此,后续对水稻、陆稻的遗传分化的研究开始重视在水-陆稻间的高分化位点。Xia等[24]利用干旱响应的EST-SSR(可表达序列标签-微卫星)标记对水稻、陆稻的适应性分化进行了研究,发现水稻与陆稻确实仅在少量EST-SSR标记上呈现出显著分化,并且相关EST-SSR标记所关联的基因也与栽培稻抗旱性有一定的联系。利用这些受选择的分子标记,能够准确区分水稻与陆稻生态型,并且可以辅助栽培稻抗旱性筛选,提高栽培稻抗旱性育种的效率[36]。Xia等[18]考察水稻、陆稻的农艺性状与抗旱表型,发现栽培稻的产量性状与抗旱性性状间普遍存在负相关关系,其潜在的遗传机制是产量基因和抗旱基因间的紧密连锁或者一因多效,解释了陆稻抗旱但产量差、水稻产量潜力高却干旱敏感的表型;他们首次提出陆稻驯化的“双向选择”机制:即陆稻在雨水充沛的年份选择高产性,在干旱缺水的年份选择抗旱性。正是在这种选择模式下,陆稻与水稻在抗旱性与产量上均产生了显著分化(图1);同时,陆稻保留了大量与抗旱相关的遗传变异与抗旱性-高产重组基因型,使其成为栽培稻抗旱性育种的重要种质资源。与此同时,节水抗旱稻的育种模式,实际上是借鉴了陆稻“双向选择”的驯化模式,采取高强度的抗旱性-优质高产交替选择,将陆稻中有利于节水抗旱的遗传资源导入水稻中[9]。所以,节水抗旱稻在水稻遗传背景下获得陆稻的节水抗旱特性,从本质上讲与陆稻在适应旱作驯化过程中逐渐进化出比水稻更好的节水抗旱性,在一定程度上是相通的[9]。
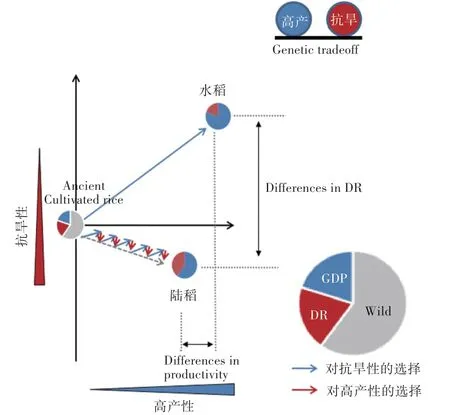
图1 水稻和陆稻适应性分化的模式图Fig.1 Pattern of adaptive differentiation between lowland rice and upland rice
水稻受到的选择主要是高产性选择,陆稻受到的是“双向选择”。因栽培稻抗旱性(DR)与高产性(GDP)之间存在tradeoff,陆稻“双向选择”的结果使陆稻的抗旱性较好而生产力较差,与水稻形成适应性分化。
虽然籼亚种内水稻、陆稻间的遗传分化程度较低,但Wang等[37]依然鉴定出籼型水-陆稻间的高分化位点,并将其与粳型水-陆稻高分化位点进行比较,发现两者仅共享1.8%的分化位点与1.6%的表达差异位点,由此认为籼型水-陆稻分化与粳型水-陆稻分化是相互独立的,且具有完全不同的遗传基础。
2.3 水稻与陆稻基因表达的分化
植物的基因表达也会受到自然选择,因此植物的适应性进化也会体现在基因表达层面,尤其是复杂性状的进化。研究水稻与陆稻在基因表达层面的分化,对于正确理解陆稻抗旱性的适应性进化,挖掘有利基因资源有着重要意义。本课题组利用转录组技术研究了典型水稻(50份)、陆稻(40份)地方种在正常水田与干旱胁迫下基因表达层面上的适应性分化,共鉴定出2 358个水-陆稻表达分化基因,但其中只有约15%的基因在DNA序列上形成遗传分化。这意味着少量关键的水-陆稻遗传分化基因就能驱动陆稻转录组水平的旱作适应。后续的基因共表达网络分析也表明:在每一个共表达网络子模块(Module)中,都有少量水-陆稻遗传分化基因是该模块的节点基因(Hub gene),这些节点基因通过基因间的共表达关系,驱动了转录组水平的旱作适应[38]。在另一项以节水抗旱稻沪旱2B为对象的研究中,同样发现来源于典型陆稻‘IRAT109’的几个关键转录因子驱动了节水抗旱稻中的抗旱基因表达调控网络,是沪旱2B抗旱性的潜在原因。由此看来,发现和应用陆稻旱作适应与抗旱性进化中的关键基因,是未来节水抗旱稻研发中的重点之一。
2.4 水稻与陆稻的表观遗传分化
表观遗传是不依赖于DNA序列变化的可遗传的表型变化,主要包括DNA甲基化、组蛋白修饰、RNA干扰与染色质构型变化,是近年来植物学领域研究的热点之一。基因组甲基化是表观遗传的重要组成部分之一,植物通过基因组的甲基化调节基因表达,在植物(包括水稻)的抗生物或非生物胁迫中起着重要作用[39-40],并且这种甲基化还会以一定的频率传递到子代中,使后代抗逆性提升[41]。
Xia等[42]利用甲基化敏感扩增多态性(Methylation sensitive amplification polymorphism,MSAP)分子标记技术研究了水稻、陆稻的表观遗传分化。结果表明:总体而言,DNA甲基化在两种生态型中的差异并不大,但在少量甲基化位点,水稻与陆稻存在显著差异,并且陆稻的甲基化水平较低。低甲基化意味着在该位点基因的表达能够更快地响应干旱,从而实现干旱适应(图2)。这可能是陆稻适应旱作的一种重要机制。
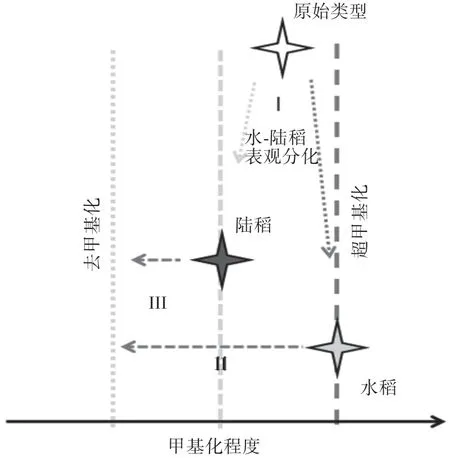
图2 基于MSAP的水-陆稻表观遗传分化模型(修改自黄卫霞[43])Fig.2 Lowland rice-upland rice epigenetic differentiation model based on MSAP
目前,水稻、陆稻表观遗传分化仍处于起步阶段,但一项基于表观遗传机制的水稻抗旱性驯化的研究则给水稻、陆稻表观遗传分化提供了一些佐证。Zheng等[44-45]利用MSAP与全基因组甲基化测序(Whole genome bisulfite Sequencing,WGBS)技术,对一个典型水稻和典型陆稻在旱地环境下连续栽种10代并复水栽培1代后的全基因组甲基化状态进行了研究,结果显示,典型陆稻与水稻之间确实存在大量的差异甲基化位点(DMC)与差异甲基化区域(DMR)。其次,发现有7%—9%受干旱诱导的DNA甲基化位点,在恢复水田种植1代后仍能保持受诱导的状态;经过连续10代旱地栽种以后,抗旱性有了轻微提高,推测可能与两个材料经过连续10代旱作后DNA甲基化状态的定向(向有利抗旱的方向)改变相关。该研究从一定程度提示了水-陆稻基因组甲基化分化的成因,以及水-陆稻的甲基化分化位点与陆稻干旱适应相关。
3 陆稻绿色遗传资源研究与利用
3.1 陆稻在栽培稻抗旱性研究中的应用
如上所述,陆稻具备较为丰富的抗旱遗传资源,因此陆稻作为一种抗旱种质资源在抗旱性的研究与育种中被广泛使用。在禾本科比较基因组网站Gramene(https:∕∕archive.gramene.org∕)上收录的栽培稻抗旱性QTL数据库中,有一半以上的抗旱相关QTL是利用陆稻种质资源构建群体进行遗传定位获得的(表2)。

表2 Gramene数据库中部分抗旱性状QTL及其所用到的陆稻种质信息Table 2 QTLs of some drought resistance traits in Gramene database and the upland rice germplasm information used
根系介导避旱性是栽培稻抵御干旱胁迫的首道防线,在栽培稻早、中期抗旱中发挥着重要作用。同时,根系构形相关的表型通常遗传力高、由主效基因调控。随着近年来各种针对根系构形相关表型(包括根长、根粗、根深等)评价方法的发展与改进,相应的遗传研究也越来越多。典型陆稻与典型水稻的根系存在十分显著的差异,这种差异有利于陆稻干旱适应,因此,陆稻发达的根系是栽培稻抗旱性遗传资源挖掘与研究的热点,陆稻是进行根系相关表型遗传定位与基因资源挖掘中最重要的种质资源。其中,来自非洲的典型陆稻‘IRAT109’根系发达、粗壮、深长,具有良好的避旱性,在栽培稻抗旱性育种中广泛应用,因此也常被用于根系相关表型的遗传定位,并取得了不错的效果(表3)。

表3 近年来利用‘IRAT109’进行抗旱相关表型遗传定位的研究Table 3 Studies on genetic mapping of drought resistance related phenotypes using‘IRAT109’in recent years
随着高通量测序技术与全基因组关联分析(Genome-wide association analysis)的发展,对自然群体根系表型的GWAS分析,结合关联位点等位基因单倍型及频率在水稻、陆稻生态型中的差异,成为挖掘根系发育与构形相关候选基因的一种重要方式。例如,Lou等[20]通过GWAS在一个由水稻、陆稻杂交构建的重组自交系群体中定位到多个与深根比相关的QTL,其中部分QTL与图位克隆的结果一致。Li等[60]在一个栽培稻自然群体中(含部分陆稻种质)针对21个根系表型通过GWAS挖掘到143个关联位点。
另外,通过比较转录组研究基因表达响应干旱胁迫的差异,也是一种常见的研究栽培稻抗旱性分子机制与挖掘抗旱候选基因的方案。其中,陆稻种质也常常作为抗旱性材料得到应用(表4)。值得一提的是,在Ma等[61]的研究中,典型陆稻‘IRAT109’是被作为敏感性材料使用的,因其抗旱性主要得益于发达的根系(避旱性),当在该研究中根系的作用被屏蔽以后,‘IRAT109’就表现出干旱敏感。因此,有一种假设认为,因为不存在犁底层,陆稻更偏好避旱性机制;水稻则由于水田犁底层的存在限制了其根系向下发展的空间,因此不得不在耐旱性机制上获得更好的进化,如两个水稻品种‘Dular’和‘IR5’在IRRI的相关研究中就表现出不逊于陆稻的抗旱性[62]。但该研究在另一陆稻品种‘IAC1246’中发现了2个阿魏酸代谢相关的耐旱性候选基因,通过干旱胁迫下的光合保护机制起到耐旱作用[61]。由此可见,陆稻在旱作适应中也能演化出一些耐旱机制。不过,在栽培稻自然群体中耐旱性与避旱性之间也可能存在一定的tradeoff,使得大多数种质不能同时获得耐旱性与避旱性[18]。这就要求在保护、研究和利用栽培稻抗旱种质资源时,需要科学、全面地对水稻、陆稻进行抗旱性评价,了解两者在不同抗旱机制上的优劣,从而利用现代育种技术去综合不同的抗旱机制,培育出更好的抗旱性栽培稻品种。

表4 近年来在转录组研究中得到应用的陆稻种质Table 4 Upland rice germplasm applied in transcriptome research in recent years
3.2 陆稻基因资源研究进展
如前所述,陆稻在长期适应旱作农业生境的过程中,产生或积累了许多抗旱基因的有利遗传变异。近年来,随着许多避旱性、耐旱性主效QTL相继被定位[20,72-74],许多抗旱基因资源也逐渐从陆稻中被鉴定出来(表5)。例如,Lyu等[72]通过研究ETAS(Elite variety tag SNP alleles)在典型水稻品种(‘IR64’‘桂朝二号’等)与陆稻品种(‘IRAT104’)间的差异,结合相关ETAS在陆稻群体中的等位基因频率与受到的选择作用,挖掘到一个控制冠根形成的候选基因Nced,在陆稻中存在一个C-T的突变,受定向选择,导致陆稻侧根表型发生改变。同样的,Li等[74]从qRT9中鉴定出一个控制根粗的基因OsbHLH120。该基因在陆稻中存在SNP与InDel,通过评价渗入系的表型,证明陆稻等位变异能使根系更为粗壮。另外,在OsJAZ1与Nal1上同样鉴定出有利抗旱的陆稻等位变异[60]。
水稻种质资源的育种应用本质上就是对有利自然变异的应用。然而,在过去的很长一段时间里,水稻重要自然变异的基础研究往往是滞后于育种实践的。有许多重要农艺性状相关基因在被鉴定之前,就被育种家们广泛地应用于育种实践中了。从表5可以看出,利用高通量手段找到的陆稻基因资源往往淹没于大量的基因组数据当中,正如Wei等[75]指出,育种家很难从大量的基因组数据和种质资源中得到需要的信息。RiceNavi是近年来第一个专门针对育种开发的基因资源整合平台,该平台对已有功能基因进行了人工筛选。其整合的功能基因均来自于种质资源,并有相应的功能验证。该平台还提供了一套包含以上绝大多数功能基因自然变异、尤其是稀有等位基因型的群体。通过多个群体的GWAS,作者对这些功能基因自然变异的表型效应进行了预估,从而辅助目标基因的选择[75]。陆稻基因资源的充分挖掘与利用,一方面是源于基因资源挖掘手段的进步,另一方面针对抗旱基因资源,也可以尝试搭建类似的平台,让近年来飞速发展的分子生物学和基因组学更好地助力育种实践。

表5 陆稻中发掘的抗旱相关基因Table 5 Drought resistance related genes discovered in upland rice

基因名OsTb2 LAX1 OsRL3.3 OsSIZ2 OsRL7.1 OsRL8.2 OsRL11.1 Nal1 OsJAZ1 OsbHLH120 Nced基因号Os09g0410500 LOC_Os01g61480 LOC_Os03g50350 LOC_Os03g50980 LOC_Os07g03160 LOC_Os08g43040 LOC_Os11g43320 LOC_Os04g52479 LOC_Os04g55920 LOC_Os09g28210 Os12g0435200基本描述在陆稻中发生一个三碱基突变,该突变在陆稻中受定向选择,导致陆稻分蘖减少,推测其有利于旱作适应利用典型水稻、陆稻构建的自然群体通过GWAS鉴定出一个已知产量LAX1,进一步发现它同时影响抗旱性,是抗旱基因“一因多效”的典型案例基于GWAS鉴定出多个与根系表型相关的QTL,并从中挖掘到5个与根长表型相关的基因,它们在陆稻中受到特定选择在陆稻该基因的CDS区域有一个转座子插入导致根系变得粗壮基因启动子区域突变导致水稻、陆稻基因表达差异在陆稻中存在SNP与InDel,该变异导致渗入系根系更为粗壮在陆稻中存在一个Cimages/BZ_21_958_1176_988_1207.pngT的突变,受定向选择,导致陆稻侧根表型发生改变等位基因来源‘IRAT104’等水稻、陆稻自然群体水稻、陆稻自然群体栽培稻自然群体、‘IRAT109’栽培稻自然群体‘IRAT109’‘IRAT104’参考文献[27][18][23][60][60][74][72]
4 陆稻抗旱性起源进化与节水抗旱稻育种理论
目前,在栽培稻抗旱性的理论研究与育种应用之间仍存在较大的差距:一方面,植物(特别是作物)的抗旱性是一个微效多基因调控的复杂性状,存在较强的基因-基因、基因-环境间的互作。截至2020年底,已经发掘和研究的栽培稻抗旱基因的数量已经超过300个,但其中真正能够在田间表现出显著抗旱效果、具有育种应用价值的却寥寥无几。另一方面,以“绿色超级稻”与“节水抗旱稻”为代表的栽培稻抗旱性育种,将陆稻的节水抗旱特性引入到高产优质的水稻背景中,实现了栽培稻的高产、稳产、节水、抗旱,取得了极大的成功。上述理论研究与育种实践间的差距,恰恰表明了栽培稻的抗旱性是一个系统性的问题,需要用系统生物学的视角和方法进行研究。这种系统性,就是陆稻抗旱性进化与节水抗旱稻育种存在的相通性,具体表现为:
(1)陆稻旱作适应性的进化,是一个系统的过程。研究表明,陆稻与水稻形成适应性分化,其中涉及的基因数量(包含遗传水平与表达水平的分化)多达上千个[18,38]。虽然数量多,但它们可以通过基因间的互作,形成复杂的调控网络,这无论是在功能基因的水平,还是在组学水平[38,76]都得到过证实。前期的研究表明,少量遗传上高度分化的基因可以通过共表达关系(潜在的调控关系)形成一个涵盖全基因组2 000余个基因的表达调控网络,使陆稻旱作适应[38,77]。其中某些基因在整个网络中发挥了重大效果,可能是促进陆稻抗旱性进化与旱作适应的关键点。因此,陆稻的适应性进化可以表述为关键基因在驯化中受到选择,并通过基因表达调控网络驱动陆稻旱作的适应[38]。
(2)节水抗旱稻的选育过程,是借鉴了陆稻驯化兼顾产量与抗旱性的双向选择,同样是一个系统性的过程。在高选择强度的抗旱性-产量交替选择下,只有能提高栽培稻抗旱性的关键基因才会在选择过程中被保留下来。有意思的是,在前期的研究中,本课题组已经发现,相对于水-陆稻的适应性分化,节水抗旱稻中来源于陆稻的遗传成分并不多,但这些成分往往居于基因表达调控网络的节点位置,能够发挥牵一发而动全身的效果[77]。因此,推测节水抗旱稻选育的遗传机制是通过抗旱性-产量的交替选择使促进陆稻旱作适应的少数关键基因进入到水稻背景中,然后通过基因表达调控网络发挥系统作用,提升节水抗旱稻的抗旱性与旱作适应性。当然这一遗传机制仍需要节水抗旱稻育种实践进行充分证明。
综上所述,无论是陆稻的进化,还是节水抗旱稻的育种,都是关键基因(位点)在这个过程中受到选择,并且在陆稻和节水抗旱稻中发挥系统性的作用,使它们在节水抗旱性与旱作适应性上得以提升。通过对这两个过程遗传机制的研究与相互印证,就能鉴定出在抗旱性进化与节水抗旱稻育种中的关键基因,推动陆稻抗旱基因资源的利用与节水抗旱稻研发。
猜你喜欢
江苏安全生产(2022年9期)2022-11-02
今日农业(2022年14期)2022-09-15
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
当代陕西(2019年12期)2019-07-12
江苏农业科学(2017年1期)2017-02-27
广西农学报(2016年3期)2017-02-06
天津农业科学(2016年12期)2017-01-11
农产品市场周刊(2014年20期)2014-09-02
金融周刊(2014年4期)2014-04-16
