
不同生长型园林植物叶片功能性状差异分析
2022-09-06 11:40姬腾岳文亚博何宝辉路兴慧
现代园艺 2022年16期
姬腾岳,文亚博,陈 琛,何宝辉,路兴慧
(聊城大学农学与农业工程学院,山东聊城 252000)
研究植物与其环境之间的相互关系与相互作用是目前农学研究的热点问题之一[1]。“功能性状”一词最早出现在20 世纪末的植物期刊上,被定义为“植物的一个重要特征,反映了植被对环境变化的反应,并能强烈影响生态系统的功能。”生态学家随后的研究工作给予了对“植物功能特性”更精确的定义,即指在植物的生命历程中,能对其定植、生长、存活和死亡有较大影响的植物性状,这些性状表明生态系统对周围生境变化的反应[2],并对生态系统产生重大影响[3]。
生态策略意味着在一定的生态压力下,任何生物都可以做出对物种生存和进化有益的对策。在生态对策上,生物物种的整体生态适应对策必然从各个方面体现出来。当前,Pierce 等[3]提出的CSR 策略理论发展而来的全球植物竞争-忍耐-杂草型策略分类法,是最成熟完善的植物生态策略研究法。该方法以植物比叶面积、叶面积、叶干物质含量3 个核心功能性状之间的关系为依据,利用StrateFy 工具计算该植物的生态策略[4]。目前,CSR 理论及其模型已经被广泛应用于物种适应性、群落过程和生态系统特性等相关的理论和实践研究中,同时这些研究也为CSR 理论提供了实证支持和扩展补充。CSR 理论及其模型不仅可用于不同生长型园林植物功能性分类,也可以作为一个概念性框架进行比较植物生态学研究,从而实现在农学以及生态学方面研究环境对生物多样性、生态系统稳定性等的影响。CSR 理论建立的植物对策类型谱共包含3 个基本生境类型。这3 个生境类型分别占据三角模型的3 个角,分别表示对3 种环境的适应性。三角模型的3 个角分别代表了竞争型植物C(Competitor)、耐胁迫型植物S(Stress-tolerator)和杂草型植物R(Ruderal)。随着植物的进化发展,形成了三角模型内部的次级生境类型,这是植物与环境相互适应的结果。
植物的功能特性是研究植物与环境之间关系的一个非常重要的方法,对功能性状的研究使得量化植物在生长过程中与环境的相互作用和探索植物生长策略成为可能[2]。园林植物是指包括花、草、树三大类的具有一定观赏价值的植物[5]。在园林建设中,园林植物作为园林绿化的植物材料之一,不仅具有美化环境的作用,也是园林生态系统的重要组成成分[5]。在园林应用中,由于不了解园林植物的生长特征而导致错误的植物配置,将会严重影响植物的景观效果。目前,关于园林植物功能性状及生态对策的研究并不多见,关于园林植物生态适应性的研究主要集中在园林植物的抗寒性、耐荫性、抗旱性、耐盐碱性、抗污染性和抗风性等方面。基于不同生长型园林植物幼苗功能性状探讨园林植物对环境的适应性以及生态策略,充分理解并掌握不同生长型园林植物的功能性状及其生态策略,能够为园林植物生态分析提供科学依据。基于此,以27 种常见的园林植物为对象,通过对27 种常见园林植物的功能性状测定、生态策略分析、园林植物学调查来了解不同生长型园林植物物种组成和功能性状差异等重要内容,运用SPSS 方差分析与多重比较和SigmaPlot 图表分析的方法,研究不同生长型园林植物功能性状以及生态策略的差异,试图从比较植物功能生态学角度探讨常见园林植物对生态功能维持的新认识,为园林植物的应用提供科学依据。
1 材料和方法
1.1 试验材料
试验地位于聊城大学(36.43°N,116.01°E),海拔为34。试验对象是27 种常见的园林植物,其中乔木、灌木、草本植物各9 种,包括酸模(Rumex acetosa)、夏至草(Lagopsis supina)、萱草(Hemerocallis fulva)、鸢尾(Iris tectorum)、玉簪(Hosta plantaginea)、地黄(Rehmannia glutinosa)、婆婆纳(Veronica didyma)、费菜(Sedum aizoon)、委陵草(Herba Potentillae chinensis)、石楠(Photinia serratifolia)、冬青(Ilex chinensis)、玫瑰(Rosarugosa)、金叶女贞(Ligustrum×vicaryi)、芍药(Paeonia lactiflora)、牡丹(Paeonia suffruticosa)、月季(Rosa chinensis)、蔷薇(Rosa multiflora)、山茶(Camellia japonica)、紫叶李(Prunus cerasifera)、银杏(Ginkgo biloba)、朴树(Celtis sinensis)、垂丝海棠(Malus halliana)、碧桃(Amygdalus persica)、苹果(Malus pumila)、黄栌(Cotinus coggygria)、梨(Cotinus coggygria)、大叶女贞(Ligustrum compactum)(详见表1)。

表1 试验材料的基本信息列表
1.2 数据采集
2022 年4 月初,在聊城大学内每个物种选择5 株生长健康的植株。对于每一株植物,采集5~10 片完好的阳性成熟叶片,迅速放入自封袋中密封,防止叶片失水。用记号笔在自封袋上记录叶片物种及编号,带回试验室进行植株功能性状测定等一系列工作。
1.3 植物功能性状测定
擦除叶片表面的灰尘,然后利用叶面积仪(WinRhizo,Regent Instruments Inc.,Quebec,Canada)测定叶片面积;称量重量并记录数值;叶绿素含量使用叶绿素计(SPAD 502Plus meter,Konica Minolta,Japan)。将测量完叶面积的叶片放入试管保存,编上序号。然后对叶片进行加热处理,在105℃下杀青处理30min,然后在80℃条件下烘干至重量,冷却后立即称量烘干重[6]。根据所测结果,分别去利用数学公式得到叶干物质含量、比叶面积的数据,计算公式如下:
比叶面积计算:比叶面积=叶片面积/叶片干重。
叶干物质含量计算:叶干物质含量=植物叶片干重/植物叶片鲜重。
1.4 数据处理
用CSR 分析工具‘StrateFy’对27 种园林植物的生态策略进行数据分析,利用方差分析比较不同物种、不同生长型叶功能性状差异。然后利用多重比较分析不同生长型叶片功能性状之间的差异。所有分析均在SPSS(SPSS Statistics 25)中进行,并在sigmaplot 中完成作图。
2 结果与分析
2.1 叶功能性状分析
由图1 可知,草本植物和灌木植物叶绿素的差异不显著(P>0.05),草本植物和乔木植物叶绿素的差异显著(P<0.05),乔木植物和灌木植物叶绿素的差异显著(P<0.05)。

图1 不同生长型园林植物叶绿素差异性分析
由图2 可知,草本植物和灌木植物比叶面积的差异显著(P<0.05),草本植物和乔木植物比叶面积的差异显著(P<0.05),灌木植物和乔木植物比叶面积的差异不显著(P>0.05)。

图2 不同生长型园林植物比叶面积差异性分析
由图3 可知,草本植物和灌木植物干物质含量的差异显著(P<0.05),草本植物和乔木植物干物质含量的差异显著(P<0.05),灌木植物和乔木植物干物质含量的差异不显著(P>0.05)。
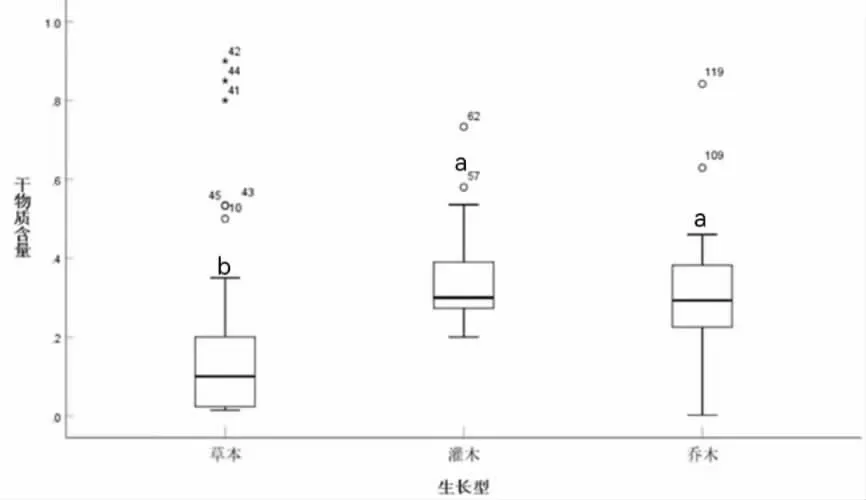
图3 不同生长型园林植物干物质含量差异性分析
2.2 生态策略分析
生长型是在一定的环境下,植物为了适应环境而表现出来的性状特征。不同生长型的园林植物以不同的生态策略适应生长环境。通过对27 种不同生长型植物CSR 权衡策略分类归纳[3],对于不同生长型园林植物,共分为12 类生态策略,有杂草/竞争-杂草型(R/CR,24%)、竞争-忍耐-杂草型(CSR,19%)、杂草/竞争-忍耐-杂草型(R/CSR,13%)、忍耐型/竞争-忍耐-杂草型(S/CSR,9%)、竞争-杂草型/竞争-忍耐-杂草型(SR/CSR,9%)、竞争/竞争-忍耐-杂草型(C/CSR,7%)、竞争-杂草型(CR,6%)、竞争-忍耐型((CS,6%)、忍耐型/忍耐-杂草型(S/SR,3%)、竞争-忍耐型/竞争-忍耐-杂草型(CS/CSR,2%)、竞争-杂草型/竞争-忍耐-杂草型(CR/CSR,1%)、忍耐型/竞争-忍耐型(SR/CSR,1%)。其中C/CSR-CS/CSR-S/CSR,这3种生长策略形成了一个抗干扰能力(R)较强的C-S 权衡策略轴,代表物种有牡丹(45∶29∶27%)、山茶(45∶41∶14%)、石楠(25∶60∶15%)。同时,C/CSR-CS-S/CS这3 个策略形成了抗干扰(R)较弱的C-S 权衡策略轴,代表物种有苹果(20∶67∶13%)、大叶女贞(42∶50∶8%)。其中CSR 策略植物是所有策略中各部分投资和能力最均衡的策略,代表物种有梨(36∶25∶39%)(见图4)。

图4 不同生长型园林植物生态策略分析
对于不同生长型园林植物生态策略的研究分析表明,草本植物偏向于R 策略,灌木偏向于CS 策略,乔木偏向于CS 策略(表1)。
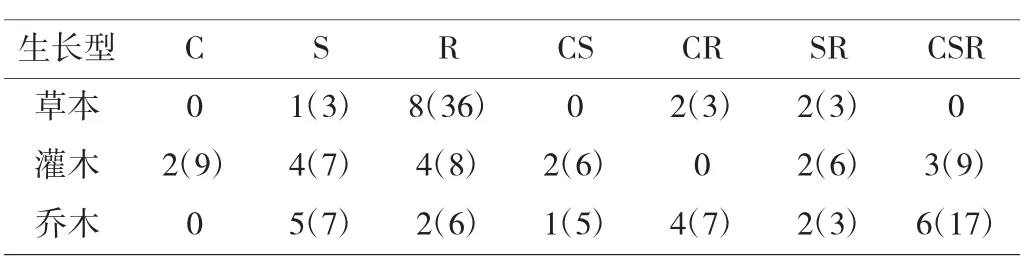
表1 不同生长型园林植物生态策略基本概况
3 讨论与结论
不同生长型植物之间的功能特征不同,表明每种生长型园林植物通过调节自身功能性状来适应生长环境[7]。不同生长型园林植物在生长过程中通过协调自身的功能性状来适应外界多变的环境,以提高自身获取资源的效率,增强自身生存能力。植物叶绿素是研究不同生长型园林植物叶片功能性状的一个关键要素,并在一定程度上决定了群落系统的生态结构和功能[8]。本研究中,草本、灌木、乔木SPAD 值呈下降趋势,表明生长在光资源较弱环境中的草本植物能够通过增加体内的叶绿素含量来获取更多的光资源。本试验中采集的灌木和草本植物均来自光照资源不充足的林下环境,因此光照成为灌木和草本植物生长的限制因子。灌木和草本植物具有较高的叶绿素含量,以提高自身的光合作用,适应低光环境。结果表明:不同生长型园林植物的叶绿素含量都表现出随其生长型变化而变化以适应环境,这与文陇英等[9]的研究结果一致。这可能与植物所处环境资源不同以及光辐射变化有关。植物存在逆境保护机制,随着环境资源变化会改变叶绿素含量以适应叶片对光的吸收来保护其免受光损伤。植物比叶面积反映了植物获取资源的能力:在资源较丰富的环境中生长的物种通常具有较高的SLA[10]。SLA 与物种分布密切相关[11]。本研究中,灌木和乔木植物的幼苗有相对较低的SLA,其叶寿命相对较长,长的叶子寿命增加了叶子面积和对光线的捕捉,这在一定程度上使植物实现了高能量储存,是乔木和灌木在不同环境中生存和发展的重要途径。叶干物质含量体现了植物获取资源的能力,其含量越高,说明叶片抵抗机械损坏的能力越高。本试验中草本与灌木干物质含量差异显著,草本与乔木干物质含量差异显著,灌木与乔木差异不显著。其中草本植物叶干物质含量最低,表明草本植物的抗物理伤害能力最低,灌木和乔木则比较高,说明环境变化对不同生长型园林植物叶片干物质含量有显著差异。本试验研究结果表明草本植物偏向于R 策略,灌木偏向于CS 策略,乔木偏向于CS 策略。这些研究结果表明,不同生长型的植物具有不同的生物量分配和生态适应策略,这些不同生长型的植物以不同方式响应适应其生长环境。
不同生长型园林植物通过调节自身功能性状来适应生长环境,本试验结果证实了不同园林植物由不同的性状组合形成不同的生态策略以实现在园林植物群落中的共存。本试验基于园林植物功能性状,探讨了不同生长型园林植物对环境的生态适应策略,对园林植物的养护管理与园林植物的科学应用具有重要的现实意义。
猜你喜欢
现代农村科技(2022年7期)2022-11-18
农学学报(2022年8期)2022-11-16
农业与技术(2022年16期)2022-09-01
诗潮(2019年10期)2019-11-19
阅读与作文(英语高中版)(2019年9期)2019-11-07
农民致富之友(2017年12期)2018-01-31
诗潮(2017年2期)2017-03-16
中国绿色画报(2016年7期)2016-12-26
江苏农业科学(2016年9期)2016-11-28
