
模拟干旱和磷添加对热带低地雨林叶凋落物分解的影响
2022-08-31 06:08:26王文秀栾军伟李丝雨梁昌强孔祥河刘世荣
生态学报 2022年15期
王文秀,栾军伟,王 一,杨 怀,赵 阳,李丝雨,梁昌强,孔祥河,刘世荣
1 国际竹藤中心, 竹藤科学与技术重点实验室, 北京 100102 2 中国林业科学研究院森林生态环境与自然保护研究所, 国家林业和草原局森林生态环境重点实验室, 北京 100091 3 海南甘什岭自然保护区, 三亚 572014
森林凋落物是森林植物在其生长发育过程中新陈代谢的产物,在森林生态系统养分循环中起着重要作用[1]。通过凋落物的分解作用,有机物质从地上植被转移进入森林土壤,逐步把养分归还给土壤[1—2],向大气释放CO2,为植物和微生物提供养分[3—4]。凋落物分解过程受自身基质质量[5—6]、环境气候条件[7]、土壤理化性质[8]、分解者群落调控[9]。并且,在已有关于凋落物分解研究中,不同功能型(常绿落叶树种、固氮非固氮树种、叶质地分解难易程度)植物的叶片基质质量存在差别[10—11],落叶树种、固氮树种叶凋落物相比于常绿树种、非固氮树种叶凋落物分解较快[11—12]。凋落物分解是生态系统中碳(C)、氮(N)、磷(P)等营养元素循环的重要环节,是生物圈与大气圈之间C、N流通的重要环节[13—15]。
温室气体排放的增加所导致的全球范围内温度升高、水文循环改变成为科学界的共识[16],影响地区包括热带地区[17—18],并且有研究学者指出未来气候变化可能导致极端干旱事件发生的频度、强度增加[16, 19]。热带雨林在水分胁迫条件下,会变得非常脆弱,地上生物量减少[20—21]、树木死亡率升高[22—23]。有研究学者在亚马逊地区开展减雨试验,连续4a时间减少了60%的雨水,得出树木死亡率增加[24]、冠层含水量减少[25]、地上净初级生产量减少[26]的结论。
气候变化从许多方面对凋落物分解产生影响,其中水分条件的变化是影响凋落物分解的重要方面之一。因为降水量减少,会削弱雨水对凋落物的破碎、淋溶作用[27];其次,土壤生物的丰富度与活性受土壤含水量调控,间接使凋落物分解与营养元素释放受影响[28];此外,降水量的减少在一定范围内会使植被生产效率降低,进而导致凋落物生产量降低[29]。干旱胁迫降低凋落物的分解速率这一观点被许多研究支持,比如Willcock等人[30]研究表明,降雨量减少会显著抑制土壤生物活性,从而抑制凋落物分解;有大量研究表明凋落物分解速率和降雨量正相关,水分不足会降低凋落物分解速率[30—31];干旱处理减慢了凋落物分解速率,并且改变了混合凋落物间的相互作用[32]。所以,气候变化可能通过降雨量减少、干旱胁迫影响凋落物分解[33]。甚至,气候变化导致的干旱事件会改变森林凋落物的分解过程,增加对森林生态系统C收支预测的不确定性[34]。
热带、亚热带地区,土壤显酸性,土壤中的P元素易与铁铝氧化物发生结合反应,并且P主要来源于矿石的淋溶,所以土壤中P元素有限;同时土壤中的P元素极易随着雨水的淋溶、冲刷作用流失,因此普遍认为热带地区受P限制[35]。并且有研究证实,P的可获得性确实限制了热带地区的净初级生产力[35]、凋落物分解[36—39]。有研究学者指出,P添加可提高植物地下生物量的累积[40];也有研究学者指出,在土壤中添加P,对于凋落物的分解速率没有显著影响,但是抑制了N损失[41];在土壤N含量高、P含量低的热带森林生态系统中,P添加可促进叶凋落物分解[37—38],但也有研究得出相反结论,在Costa Rich、Brazil的热带生态系统实验中,认为P添加对叶凋落物分解过程中的质量损失没有较大作用[42—43]。目前,土壤中P元素可用性是否调控干旱胁迫对热带森林凋落物分解的影响并不清楚。
基于以上研究背景,本文提出如下研究假设:模拟干旱处理使试验样地的土壤含水量降低,减慢了叶凋落物分解过程中生物、化学反应速率,抑制了参与凋落物分解的分解者的活性,抑制了凋落物分解;P添加处理增加了土壤中P元素的可获得性,可能改变土壤微生物群落结构,尤其是真菌群落,进而可能影响凋落物分解速率;基于上述原因,本文推测土壤中水分含量降低与P可利用性提高可能存在交互作用,土壤中P可利用性增加可在一定程度上缓解土壤水分含量下降对叶凋落物分解的抑制作用。
1 材料与方法
1.1 研究区概况
本实验研究地位于海南省三亚市甘什岭省级自然保护区,国家林草局海南三亚竹藤伴生林生态系统定位观测研究站。该区属于热带低地雨林,地理坐标是109°34′E—109°42′E,18°20′N—18°21′N。地貌是低山丘陵,年平均日照时数大约为2563.0h,年均温25.4℃,每年有明显干湿两季,旱季时间为当年10月至明年5月,除此之外均为雨季时间,平均年降水量约为1800mm,属热带海洋性季风气候。土壤母质为花岗岩,属粗砂粘红壤,砂砾含量大,部分区域土层贫瘠,雨水冲刷导致岩层裸露率高,部分区域达到70%以上,平均林冠高度为9.7—12.1m,部分区域至15m以上,差异较大,郁闭度0.6—0.9,区域内群落结构复杂,植被种类丰富[44]。乔木层主要有铁凌(Hopeareticulate)、青皮(Vaticamangachapoi)、阿芳(Alphonseamonogyna)、琼楠柿(Diospyroshowii)、崖柿(Diospyroschunii)等;草本植物主要有高秆珍珠茅(Scleriaelata)和益智(Alpiniaoxyphylla);藤本植物主要有多叶花椒(Zanthoxylummultijugum)、清香藤(Jasminumlanceolarium)、锡叶藤(Tetraceraasiatica)等[45]。
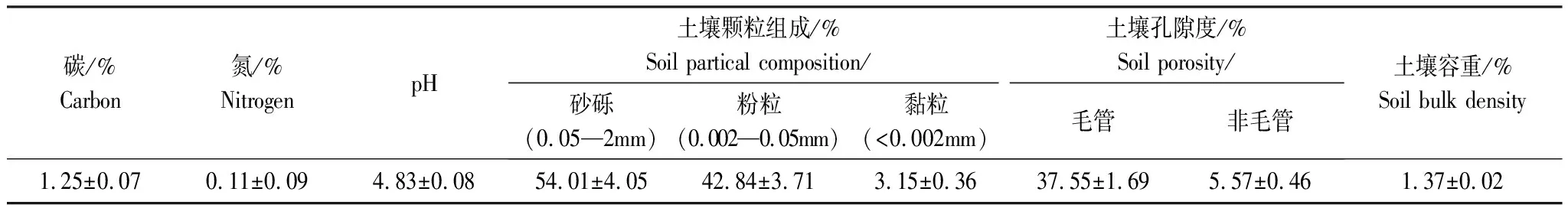
表1 实验样地土壤理化性质
数据为平均值±标准误
1.2 实验设计
依托海南三亚竹藤伴生林生态系统定位观测研究站的长期穿透雨减少与P添加双因素交互控制实验平台,共3个区组,每个区组由4个40m×40m的样地组成,并采取4种不同的处理,分别为穿透雨减少(模拟干旱)(D,50%穿透雨减少)处理、P添加(P,50Kg P hm-2a-1)处理、模拟干旱×P添加(DP,50%穿透雨减少×50Kg P hm-2a-1)处理、对照(CK)样地。每块样地四周铺设PVC软板,阻止样地内外的水分交换。穿透雨减少采用不锈钢管结合PEP透光膜在林冠下离地表约2m处建立拦截架,拦截50%的林内穿透雨、导入排水槽排出样地;P添加采取人工撒施Ca(H2PO4)2·H2O(TSP)于地表,施P强度为50Kg P hm-2a-1,从2019年7月开始,起初施P频率为1次/2月,2020年后稳定在1次/4月(施P频率降低,总量不变)。本实验根据样地内各树种多度、碳固持类型、叶质地,选取4个相对密度较高树种的叶凋落物为研究对象,分别为铁凌(Hopeaexalata)、白茶树(Koilodepasbainanense)、黑叶谷木(Memecylonnigrescens)、山油柑(Acronychiapedunculata),2019年10月收集铁凌、白茶树、黑叶谷木、山油柑叶凋落物,将收集的叶凋落物于70℃下烘干24h,用于测定4种叶凋落物初始化学成分含量与野外分解试验布设。本研究采用尼龙网分解袋,大小为20cm×30cm,网孔1mm×1mm,每个分解袋中装8g叶凋落物(4个树种×4个处理×6个重复×4次回收=384袋)。于2019年11月,将全部叶凋落物分解袋随机布置于样地地表,分解袋回收时间分别为2020年1月、5月、6月、8月(取样时间受疫情影响),对应分解时间分别为57d、178d、230d、273d。每次回收,每个树种叶凋落物4个处理下分别取6袋(每个样地分别取2袋),取样后将分解袋中的杂物泥沙处理干净,于70℃烘干24h,称量、粉碎、测定C、N含量。

表2 4种凋落物树种性质
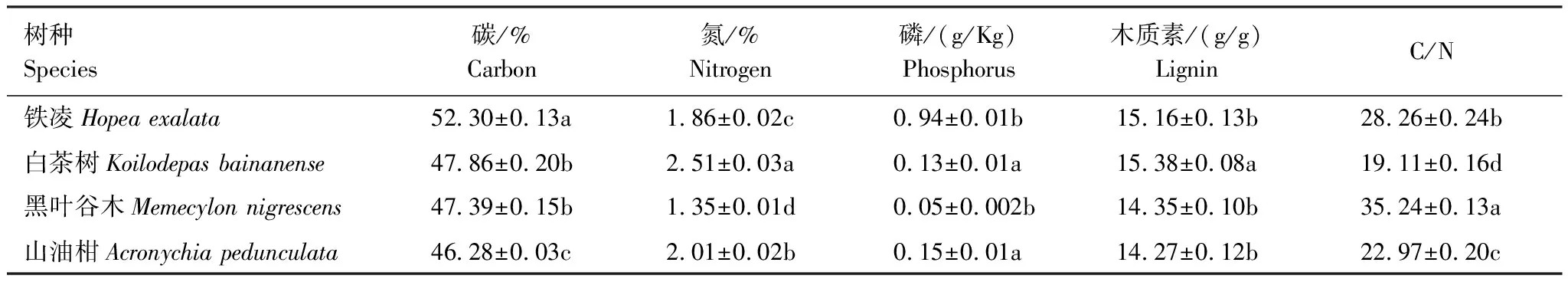
表3 4种叶凋落物化学成分含量
1.3 凋落物化学成分测定
凋落物总C含量的测定:元素分析仪(ECS 4010 CHNSO, Costech, Italy)。
凋落物总N含量的测定:元素分析仪(ECS 4010 CHNSO, Costech, Italy)。
凋落物初始全P含量的测定:硫酸-高氯酸法消煮(300℃,3h),采用钼锑钪比色法通过化学元素分析仪(Smartchem 300, AMS-Alliance, Italy)。
凋落物初始木质素含量的测定:重量法(GB/T 35818/2018)。
1.4 叶凋落物质量残存率、分解系数以及养分损失率计算


C、N损失率CL、NL(%):CL=[(M0×C0)-MT×CT]/(M0×C0)×100%;
NL=[(M0×N0)-MT×NT]/(M0×N0)×100%
式中,Mt表示分解t时间后凋落物残余质量占初始质量的比例(%);MT表示分解t时间后凋落物残余质量(g);M0表示凋落物初始质量(g);t表示分解时间(a);k表示分解系数(a-1);C0、CT表示凋落物初始C含量、回收时剩余C含量(%);N0、NT表示凋落物初始N含量、回收时剩余N含量(%)。
1.5 数据分析
使用SPSS 22.0软件进行统计分析,用单因素方差分析(One-way ANOVA),比较4种叶凋落物初始化学性质之间的差异是否显著;用一般线性模型(General Linear model),分析凋落物基质质量、分解时间、模拟干旱、磷添加对凋落物分解系数、C、N损失的影响;用线性回归分析(Linear regression analysis),分析叶凋落物分解系数、C、N损失与初始化学组分含量的关系;用双因素方差分析(Two-way ANOVA),检验模拟干旱、P添加处理分别对4个树种叶凋落物分解系数、C、N损失的影响。作图软件使用Origin 2018。
2 结果与分析
2.1 基质质量对叶凋落物分解的影响
凋落物基质质量对叶凋落物分解系数、C、N损失率均有显著影响(P<0.001;P<0.001;P<0.001)(表4),表现为白茶树叶凋落物的分解系数大于其它3个树种叶凋落物的分解系数,白茶树叶凋落物的C损失率大于
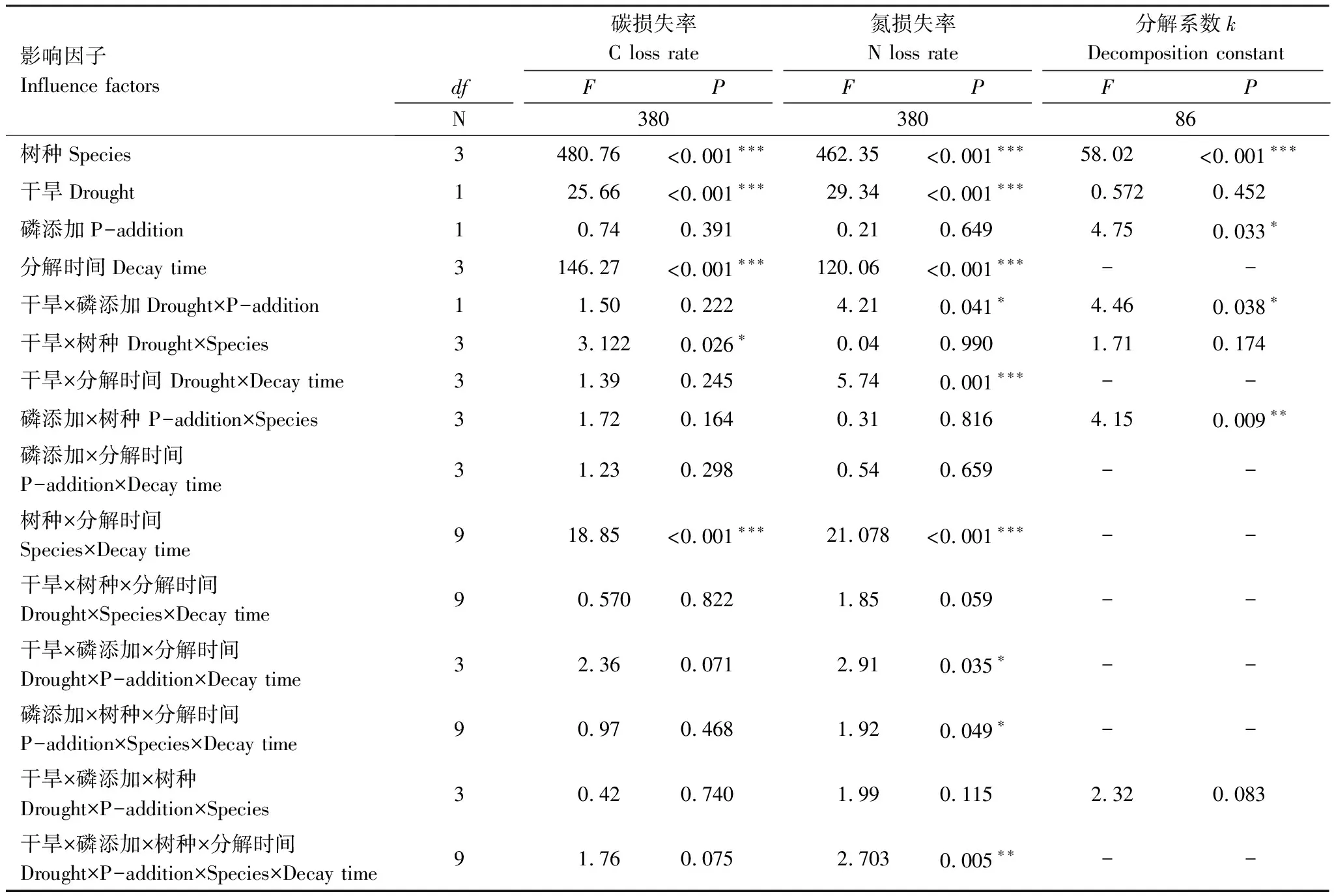
表4 树种、模拟干旱、磷添加、分解时间及交互作用对叶凋落物分解的影响
其它3个树种叶凋落物的C损失率,N损失率由大到小依次为白茶树、山油柑、铁凌、黑叶谷木(图1)。并且,叶凋落物分解系数、C、N损失率与凋落物初始N含量、C/N比存在显著正线性相关、负线性相关关系,比如,初始N含量较高、C/N比较低的白茶树叶凋落物分解较快,C、N损失率较高(图2)。

图1 4个树种叶凋落物碳、氮损失率、分解系数Fig.1 Decomposition constants, C, N loss rates of four-tree species leaf litter
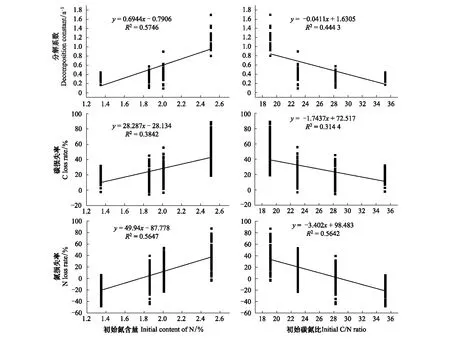
图2 叶凋落物初始氮含量、碳氮比与碳、氮损失率、分解系数的关系Fig.2 The relationship between initial N content, C/N ratio and C, N loss rate, decomposition constant
2.2 模拟干旱处理对叶凋落物分解的影响
4个树种叶凋落物分解过程中的分解系数,模拟干旱处理使铁凌叶凋落物分解系数显著降低了11%(P=0.012),但对其它树种叶凋落物无显著影响(图3)。
4个树种叶凋落物的C含量,分解57d后,模拟干旱处理使白茶树叶凋落物的C损失率下降了14.46%(P=0.042)(图4);分解178d后,模拟干旱处理使白茶树叶凋落物C损失率显著下降了12.24%(P=0.044)(图5);分解230d后,模拟干旱处理使白茶树叶凋落物C损失率显著上升了0.6%(P=0.033)(图6);分解273d后,模拟干旱处理对铁凌、白茶树叶凋落物C损失率有显著影响,分别使铁凌叶凋落物C损失率显著下降了21.65%(P=0.050),使白茶树叶凋落物C损失率显著下降了13.72%(P=0.010)(图7)。并且模拟干旱处理对叶凋落物C损失率的影响同时还与叶凋落物的树种有关(P=0.026)(表4)。
模拟干旱处理对叶凋落物N损失率均产生显著影响(P<0.001)(表4)。4个树种叶凋落物的N含量,分解57d后,模拟干旱处理对4个树种叶凋落物N损失均产生显著影响,分别使铁凌叶凋落物N损失率下降了806.62%(P=0.003);使白茶树叶凋落物N损失率下降了64.93%(P=0.022);使黑叶谷木叶凋落物N损失率上升了15.13%(P=0.046);使山油柑叶凋落物N损失率下降了195.34%(P=0.015)(图4);分解178d后,模拟干旱处理使山油柑叶凋落物N损失率显著下降了108.69%(P=0.024)(图5);分解230d后,模拟干旱处理使黑叶谷木叶凋落物N损失率显著下降245.74%(P=0.001)(图6);分解273d后,模拟干旱处理使白茶树叶凋落物N损失率显著下降了11.95%(P=0.005)(图7)。
2.3 磷添加处理对叶凋落物分解的影响
P添加处理对叶凋落物分解系数存在显著影响,使叶凋落物分解系数升高0.64%(表4;P=0.033),但该影响与叶凋落物树种无关(表4;图3)。
4个树种的叶凋落物C含量,分解57d后,P添加处理使铁凌叶凋落物C损失率增加183.42%(P=0.004)(图4);分解178d后,P添加处理对4个树种叶凋落物C损失率均不产生显著影响(图5);分解230d后,P添加处理对4个树种叶凋落物C损失率均不产生显著影响(图6);分解273d后,P添加处理对4个树种叶凋落物C损失率均无显著影响(图7)。
4个树种的叶凋落物N含量,分解57d后,P添加处理对4个树种叶凋落物N损失率均无显著影响(图4);分解178d后,P添加处理使黑叶谷木叶凋落物N损失率显著下降了58.62%(P=0.039)(图5);分解230d后,P添加处理对4个树种叶凋落物N损失率均不产生显著影响(图6);分解273d后,P添加处理对4个树种叶凋落物N损失率均无显著影响(图7)。
2.4 模拟干旱与磷添加处理对叶凋落物分解的交互效应
模拟干旱与P添加处理叶凋落物分解系数产生显著交互效应,在仅模拟干旱处理下未对叶凋落物分解系数产生显著影响,但当模拟干旱、P添加处理同时存在时,使叶凋落物分解系数降低了18.74%(表4,P=0.038),且交互效应与叶凋落物树种无关(表4;图3)。
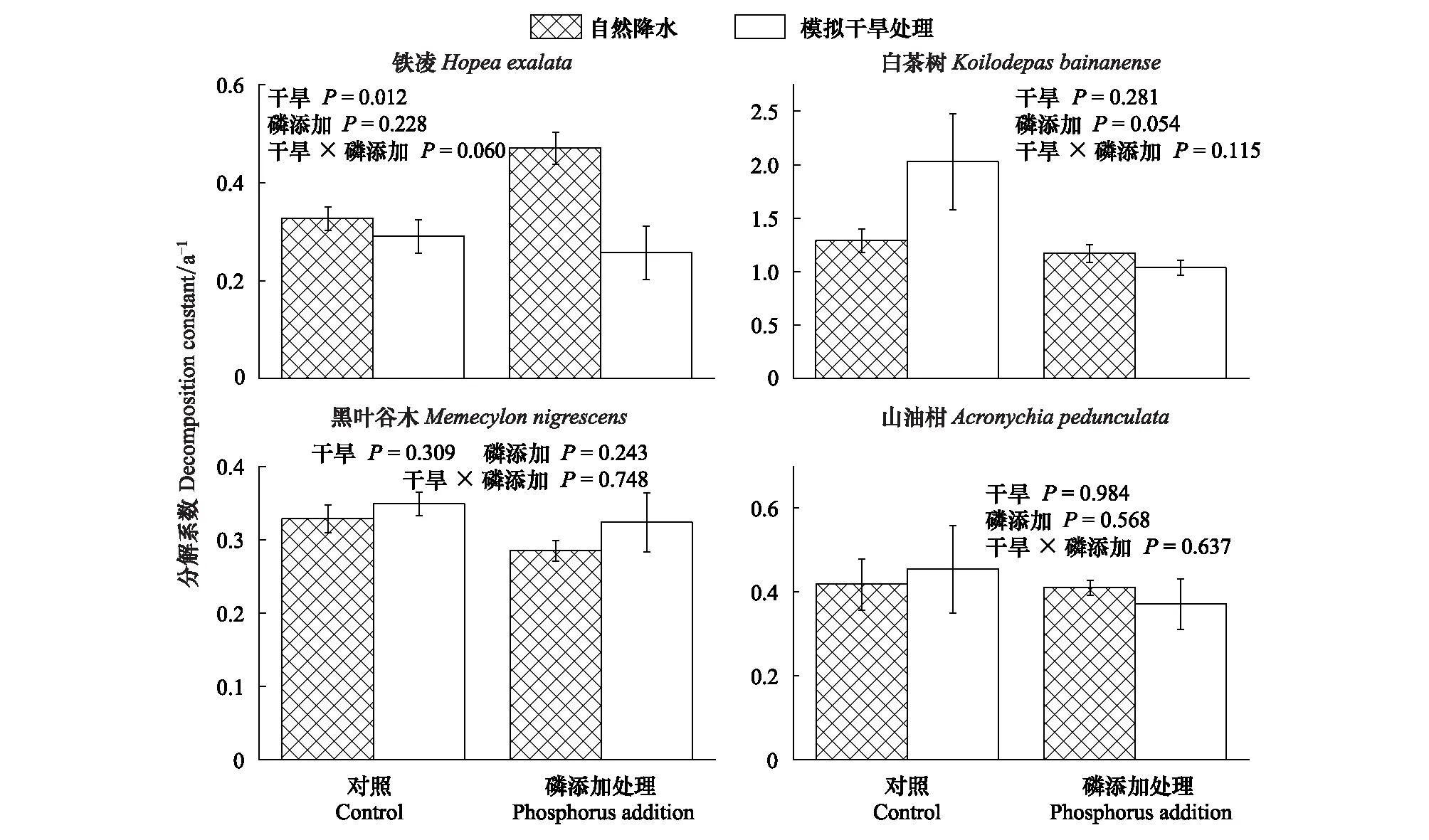
图3 4种叶凋落物的分解系数Fig.3 Decomposition constants of four-tree species leaf litter
4个树种的叶凋落物C含量,分解57d后,模拟干旱与P添加处理对4个树种均未产生显著交互效应(图4);分解178d后,模拟干旱与P添加处理对4个树种叶凋落物C损失率均不产生显著交互效应(图5);分解230d后,模拟干旱与P添加处理对白茶树叶凋落物C损失率产生显著交互效应,在仅模拟干旱处理下,白茶树叶凋落物C损失率增加0.06%(P=0.033),但当模拟干旱与P添加同时作用时,C损失率显著下降了31.95%(P=0.028)(图6);分解273d后,模拟干旱与P添加处理对4个树种叶凋落物C损失率均无显著交互效应(图7)。

图4 分解57d后,4种叶凋落物的碳、氮损失率Fig.4 C, and N loss of four-tree species leaf litter, after decaying 57 days

图5 分解178d后,4种叶凋落物的碳、氮损失率Fig.5 C, and N loss of four-tree species leaf litter, after decaying 178 days
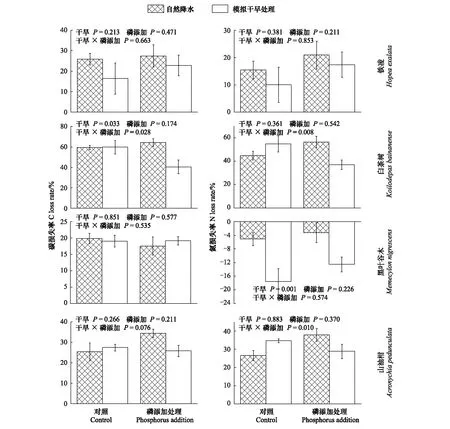
图6 分解230d后,4种叶凋落物的碳、氮损失率Fig.6 C, and N loss of four-tree species leaf litter, after decaying 230 days
4个树种的叶凋落物N含量,分解57d后,模拟干旱与P添加处理对黑叶谷木叶凋落物N损失率有显著交互效应,在仅模拟干旱处理下,黑叶谷木叶凋落物N损失率增加15.13%(P=0.046),但当模拟干旱与P添加同时作用时,N损失率下降了138.6%(P=0.023)(图4);分解178d后,模拟干旱与P添加处理对4个树种叶凋落物均不产生显著交互效应(图5);分解230d后,模拟干旱与P添加处理对白茶树、山油柑叶凋落物N损失率存在显著交互效应,在仅模拟干旱处理下,未对白茶树、山油柑叶凋落物产生显著影响,但当模拟干旱与P添加处理同时作用时,白茶树叶凋落物N损失率显著下降了17.44%(P=0.008),山油柑叶凋落物N损失率显著上升了8.35%(P=0.010)(图6);分解273d后,模拟干旱与P添加处理对4个树种叶凋落物均无显著交互效应(图7)。

图7 分解273d后,4种叶凋落物的碳、氮损失率Fig.7 C, and N loss of four-tree species leaf litter, after decaying 273 days
3 讨论
凋落物分解系数、C、N损失与凋落物基质质量紧密相关[41]。在本试验中,分解结束时,白茶树叶凋落物较其它三种叶凋落物分解剩余质量最少、C、N损失率最高,这可能因为白茶树叶凋落物初始N含量最高、CN比最低。这与一些研究学者的结论相似,凋落物分解速率与初始N含量呈极显著正相关,是控制分解速率最主要的因素[47];N含量高、CN比低的凋落物满足土壤微生物养分需要,更易被微生物分解[48]。
本研究中,模拟干旱处理对叶凋落物分解的影响表现为抑制作用,使铁凌叶凋落物分解系数下降11%,使铁凌、白茶树叶凋落物C损失率下降,使铁凌、白茶树、黑叶谷木、山油柑叶凋落物N损失率下降。与前人研究结果一致,Salamanca等人在温带森林开展有关降雨量变化对凋落物分解影响的研究中发现,降雨量对凋落物分解的控制主要是通过雨水的淋溶作用与微生物、动物群落活动,并且根据土壤呼吸、微生物生物量数据可知,降雨量减少降低微生物、动物群落丰富度与活性,间接抑制凋落物分解[49]。同时,在本试验中,模拟干旱处理对4个树种叶凋落物的抑制作用存在差异,可能因为不同树种叶凋落物的基质质量不同。这与前人的研究结果一致,Sanaullah等人表明不同的凋落物分解中基质质量会调控干旱胁迫对分解的影响[50],原因可能是在分解过程中,微生物分解者更倾向于分解质量较高(初始N含量较高)的凋落物,所以当包括干旱在内的非生物条件限制微生物分解者的活性时,该种凋落物分解受抑制程度最大[50]。
本研究中,P添加处理对叶凋落物分解产生影响,但对C、N损失产生影响不同,促进C损失但抑制N损失,比如使铁凌叶凋落物C损失率升高,使黑叶谷木N损失率下降。原因可能是土壤中P含量的升高,促进微生物分解高C物质的能力;N和P均是植物生长过程中重要的限制因素,相互独立又相互影响[51—52],所以土壤中P含量的升高可能会影响凋落物分解中的N元素变化。首先,在本研究中,P添加处理对铁凌叶凋落物的C损失有促进作用,这与一些研究结果相似。有研究表明提高土壤中的P含量对凋落物分解有促进作用,土壤中较高的P含量可提高微生物群落与胞外酶活性[53],进而提高微生物分解高C物质的能力,比如木质素、纤维素[53—56],所以促进凋落物中C释放。其次,在本研究中,P添加处理对黑叶谷木叶凋落物N损失有抑制作用,曾有研究学者表明,比如P添加对凋落物分解产生抑制作用或无明显作用[43, 54, 57—59],原因可能是P添加抑制了微生物的P矿化作用[60],当P添加时微生物就有了足够的P,不必要通过分解凋落物等有机物来获取P[58],所以抑制了微生物分解者对凋落物的分解,但是抑制微生物P矿化这一说法很少被承认[61];还有研究学者认为P添加对凋落物分解的抑制作用因为土壤中P含量的增加提高了微生物利用C的能力,但是微生物利用C完成自身生长代谢需要,而不是用于合成凋落物分解所需的酶[62—63]。同时也有研究学者认为P添加对凋落物分解N损失存在促进作用,土壤中P的可获得性影响着土壤微生物的丰富度与活性,比如氨氧化古菌,进而增加了硝化反应速率,促进了微生物分解者从凋落物中获取N[64]。不同研究关于P添加对凋落物分解的影响结论不同,原因可能是P添加处理时的药品剂量不够大、对样地的处理时间不够长或者P限制不是研究地区凋落物分解的主要影响因子[38],也有研究学者认为高浓度的P添加处理可能对P限制地区的凋落物分解有抑制作用[58]。试验过程中,施P频率发生变化,但施P总量没有变化,并且P元素在土壤中属于缓释,所以本试验不将施P频率改变作为影响分解结果的主要因子。
本试验中,模拟干旱×P添加处理对叶凋落物分解产生了显著的交互效应。P添加缓解甚至反转了干旱处理对叶凋落物分解过程中C、N损失率的作用,说明了降水量与土壤P含量均是该地区的影响因子,以及提高土壤中P的可用性,可改变干旱胁迫的影响。前人的研究中也得到相似观点,比如有研究学者关于水分条件、P限制哪个是温带农作物生产量的主要限制因子实验中得出,在水分胁迫条件下,P添加使植物对干旱胁迫的耐受能力更强,产生更多的生物量[65]。有研究表明P添加与水分胁迫的交互作用与土地类型有关,在中度干旱的条件下,P添加有助于在黑土中生长的植物产生更多的生物量,但在砂土中不存在促进作用[66]。也有研究得到相反结论,比如有研究学者表明,干旱胁迫与P添加同时存在情况下,并未对土壤呼吸产生显著交互效应[67]。原因可能在于研究地区环境因素不同,土壤中P的可用性对不同生态系统类型的限制程度不同,或者P添加处理的浓度与时间不同,所以处理产生的效果不同。有研究指出,干旱处理与营养元素添加均是影响土壤与微生物呼吸等活动、物种丰富度的影响因子,但是他们之间的交互作用机制仍不清晰;干旱对土壤及微生物的影响作用强于P添加的作用,干旱抑制了土壤的呼吸作用,使土壤微生物群落中的多样性降低;但是,干旱与磷添加同时作用时,土壤呼吸未被抑制,土壤微生物群落中丰富度仍发生变化[67]。在本试验中,干旱与P添加对凋落物分解的影响机制,本研究猜测可能是,干旱抑制了土壤中参加分解的微生物的活性与丰富度,但是土壤中P可利用性的提高,促进了一些微生物的活性与丰富度,所以二者存在交互作用。但是,干旱与P添加哪个影响因子的作用更强,在本试验中,无法得出。
4 结论
本研究验证了不同树种叶凋落物因质量不同,分解存在差异;干旱胁迫抑制叶凋落物分解,但对不同树种叶凋落物的影响存在差异;土壤中P含量的升高会促进叶凋落物的C损失、抑制叶凋落物的N损失;提高土壤中P的可获得性在一定程度上可缓解干旱胁迫对凋落物分解的抑制。综合本试验与以往研究,模拟干旱、P添加处理对叶凋落物分解的影响结果,可能与研究地土壤理化性质、凋落物基质质量、施P浓度、时间相关。在本试验中,也有一些不足希望在以后的试验中有所改进,比如在实验设计中,应对减雨处理与P添加处理的土壤水分含量与P含量进行监控,检验处理是否在试验中产生效果;并且减雨处理与P添加处理可以设定一些梯度,更加细致的讨论土壤水分与不同程度的P可利用性对叶凋落物分解等生态过程的影响程度;微生物分解是凋落物分解过程中的一个重要途径,所以关于凋落物分解的研究离不开与微生物相关的实验,在未来除关注凋落物分解过程中的养分元素变化外,还需要关注微生物的变化。
猜你喜欢
现代园艺(2021年23期)2021-12-01 07:47:44
今日农业(2021年1期)2021-11-26 07:00:56
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
应用数学(2020年2期)2020-06-24 06:02:38
学苑创造·A版(2020年3期)2020-04-24 09:21:39
海外星云(2016年19期)2016-10-24 11:53:42
红领巾·萌芽(2016年2期)2016-09-10 07:22:44
中国蜂业(2016年3期)2016-09-06 09:03:17
河池学院学报(2014年5期)2014-02-27 13:37:18
