
NFATc1 在小鼠毛囊生长周期中的定位和表达
2022-08-31 07:07:36范阔海张敏爱胡燕姣刘艺王慧荣于秀菊
山西农业大学学报(自然科学版) 2022年4期
范阔海,张敏爱,胡燕姣,刘艺,王慧荣,于秀菊*
(1.山西农业大学 实验动物管理中心,山西 晋中 030801;2.太原海关技术中心,山西 太原 030000;3.山西农业大学 动物医学学院,山西 晋中 030801)
活化T 细胞c1 核因子(nuclear factor of activated T-cells,NFATc1)是NFAT 家族成员之一,主要参与调控细胞的增殖、分化、凋亡、炎症反应、血管生成和肿瘤的发生发展[1-2],近年来的研究证实,NFATc1 表达于哺乳动物多种组织中[3],并且发现其在免疫系统和被皮系统中也发挥着重要的调控作用。 相关研究表明:紫外线会降低NFATc1 的磷酸化水平,导致基质金属蛋白酶活性增加、胶原蛋白降解,从而引发皱纹的形成。Hwang 等研究证明,荨麻、羽衣草和消皮素C 等药物可以通过调控NFATc1 的转录水平从而降低紫外线对皮肤造成的伤害[4-6]。Hariri 等在对Axenfeld Rieger 综合征患者进行基因分析和表型变化的研究中发现,Nfatc1基因的突变会导致毛囊细胞的 退 化 和 汗 腺 的 缺 失[7]。 Horsley 等 制 备 的NFATc1 缺失型小鼠表现为毛发的过度生长,并对该现象的分子机制进行研究发现NFATc1 具有维持毛囊干细胞保持静止状态的生物学功能,NFATc1 的缺失导致毛囊干细胞被激活,毛囊始终处于生长时期,从而引发毛发生长过剩[8]。
毛囊作为皮肤中重要的附属结构,对于人和动物都具有重要的意义。其生长呈现周期性的变化过程,大致分为3 个时期:毛囊生长期(Anagen)、退 化 期(Catagen)和 相 对 静 止 期(Telogen)[9]。各种动物的毛囊生长周期的时间长短不同,例如:小鼠的毛囊生长周期约为28 d[10]。毛囊的周期变化受到Hox 基因、角蛋白(Keratins)、βcatenin 等一系列的基因和蛋白的调控。近年来研究发现,毛囊的周期性变化与毛囊干细胞的激活与否有着密切的关系。截至目前,已经证明的毛囊干细胞标记物有多种,主要包括CD34,还有Lhx2、Sox9、Tcf3、Nfatc1等转录因子[11-12]。前面的研究表明NFATc1 与毛囊干细胞具有密切的关系,但在小鼠自然毛囊生长周期性中的作用如何尚不清楚。本研究通过免疫组织化学,荧光定量PCR 和Western blotting 技术对NFATc1 在小鼠毛囊生长周期中的组织定位、mRNA 和蛋白表达量进行分析,探寻NFATc1 在小鼠毛囊周期性生长过程中发挥的作用。
1 材料和方法
1.1 试验材料
20 只2 月龄的ICR 小鼠(属于Swiss 小鼠,由美国癌症研究所(Institute of Cancer Research)的3个首字母命名,是国际通用的封闭群小鼠,具有繁殖力高、适应性强和实验重复性较好的特点),雌雄各半,体重(20±2)g,购自北京华阜康生物科技股份有限公司。饲养条件为:温度(23±1)℃,湿度为45%左右,自由采食和饮水。交配确定受孕后将雌鼠单独饲喂,选择同一天生产的5 窝小鼠,于出生后的第15 天(处于毛囊生长期),第19 天(处于毛囊退化期)和第21 天(处于毛囊相对静止期),每窝取1 只小鼠,麻醉后取其背部皮肤,将其皮肤组织置于Bouins 固定液中固定24 h,70%~100%的梯度酒精脱水,1∶1 酒精与二甲苯处理后,二甲苯透明后浸蜡包埋,制成厚度为5 μm 的石蜡切片,用于免疫组织化学分析;将其部分皮肤组织置于液氮中保存,按照Trizol 法和组织总蛋白抽提试剂盒的方法分别提取小鼠皮肤的总RNA 和总蛋白,用于荧光定量PCR 和Western blotting 检测。兔抗NFATc1 多克隆抗体购自武汉三鹰生物技术有限公司;β-actin 多克隆抗体和HRP 标记的山羊抗兔IgG 购自北京康为世纪生物科技有限公司;反转录试剂盒和荧光定量PCR 试剂盒购自宝生物(大连)工程有限公司,组织蛋白抽提试剂盒购自北京普利莱基因技术有限公司。
1.2 试验方法
1.2.1 免疫组织化学分析
制备好的石蜡组织切片,二甲苯脱蜡,70%~100%的梯度酒精复水至蒸馏水;3%过氧化氢用于去除内源性的过氧化物酶,PBS 洗3 次,每次5 min;5%非免疫性牛血清作为封闭液,37 ℃孵育30 min;去除封闭液后滴加1∶100 兔抗NFATc1 多克隆抗体,室温孵育30 min,4 ℃过夜孵育,室温孵育30 min;PBS 洗3 次,每次5 min,滴加HRP 标记的山羊抗兔IgG,37 ℃孵育30 min,PBS 洗涤3 次,每次5 min;滴加DAB 显色剂显色3 至5 min,PBS洗3 次,每次5 min;苏木精轻度复染,70%~100%的梯度酒精脱水后二甲苯透明,中性树胶封片。非免疫牛血清代替一抗作为阴性对照。
通过Image-Pro Plus 6.0 软件对NFATc1 蛋白在小鼠皮肤组织中的免疫组织化学结果进行光密度的测定,每个时期的3 只小鼠皮肤组织切片中分别取5 个视野,获得阳性细胞的光密度值。所得数据用SPSS 软件进行统计学分析。分析结果用平均值±标准误(Mean±SE)来表示。
1.2.2 荧光定量PCR 分析
电泳检测提取的小鼠皮肤总RNA 的完整性,ND-1000 超微量核酸蛋白测定仪测定其浓度并将各个样本的浓度调至同一浓度。按照反转录试剂盒提供的方法完成cDNA 的合成。
根据NCBI 小鼠Nfatc1的基因序列,借助Primer5.0 软件,设计用于荧光定量PCR 检测的特异性引物,并由上海生工生物工程有限公司进行合成。引物序列为:Forward primer:5’-CCCGTCACATTCTGGTCC-3’,Reverse primer:5’-GCTGCCTTCCGTCTCATAG-3’。以合成cDNA 为模板,以特异性引物进行荧光定量PCR 分析,反应在八连管中进行。反应体系为:TB Green Fast qPCR Mix(2×)12.5 μL、Forward primer 1 μL、Reverse primer 1 μL、cDNA 2 μL、ROX Reference Dye II(50×)0.5 μL、加dH2O 至25 μL。反应条件为:95 ℃预变性30 s;95 ℃5 s,60 ℃10 s,40个循环。每个样本、Nfatc1基因、β-actin基因3 个重复。由熔解曲线判断PCR 的特异性,根据荧光曲线获得CT值,利用2-△△CT分析定量的结果。
1.2.3 Western blotting 检测
BCA 法测定提取的小鼠皮肤总蛋白的浓度,并将各个样品的蛋白浓度归至同一浓度。每个样品总蛋白上样量为200 μg,在8%的SDS-PAGE进行电泳分离后转至PVDF 膜。PVDF 膜于5%脱脂蛋白粉封闭液中孵育1h,孵育一抗(1∶1000 兔抗NFATc1 多克隆抗体;1∶500 兔抗β-actin 多克隆抗体),室温放置30 min,4 ℃孵育过夜,室温放置1 h。TBST 洗膜3 次,每次5 min,孵育二抗(1∶10 000,HRP-山羊抗兔IgG),37 ℃孵育1 h。TBST洗膜4 次,每次10 min。运用ECL 试剂盒进行曝光之后对其信号条带扫描。使用Image J 软件对小鼠毛囊不同生长时期的皮肤组织中NFATc1 和β-actin 蛋白的灰度值进行分析。
2 结果与分析
2.1 NFATc1 在小鼠毛囊生长周期中的定位
NFATc1 的免疫组化阳性反应物主要分布于毛囊的膨大部、根鞘、皮脂腺和表皮,在毛囊生长的3 个时期均有表达,但在不同的毛囊生长时期表达量存在差异。在毛囊的生长期,毛纤维生长茂盛,整个毛囊的长度不断增长达到最长,毛囊乳头直径显著大于根鞘部直径,在毛囊的根鞘部、皮脂腺和表皮可见NFATc1 的阳性反应物(图1A);毛囊进入退化期,毛囊乳头萎缩成杵形的圆状结构,整个毛囊的长度明显变短,这个时期,在毛囊的膨大部、皮脂腺和表皮存在NFATc1 较强的阳性表达物(图1B);在短暂的退化期(小鼠3 d 左右)后毛囊进入相对静止期(图1C):毛芽生长,膨大部的干细胞不断分化成内根鞘和毛干向上推移,接近“杵形毛”,杵形毛最终脱落,从而新毛占据旧毛穴。在毛囊的相对静止期,NFATc1 的阳性物主要是在毛囊的膨大部。对照组中均未见阳性反应(图1D~图1F)。NFATc1 在毛囊生长期、退化期和相对静止期的平均光密度值分别是0.17±0.012 0、0.48±0.014 7 和0.23±0.001 8。
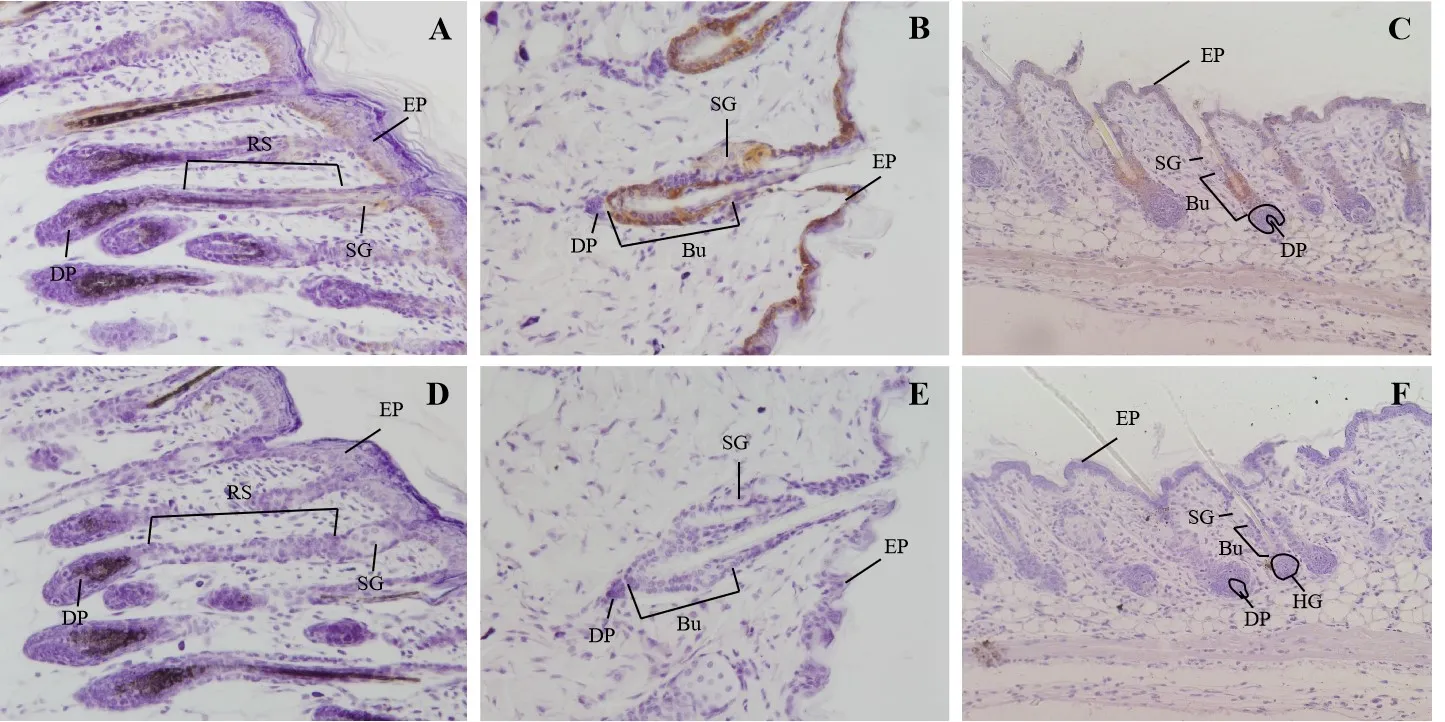
图1 NFATc1 在小鼠生长周期中的免疫组织化学染色Fig.1 Immunohistochemistry of NFATc1 in the hair follicle cycle in mice
2.2 NFATc1 在小鼠毛囊生长周期中的基因表达差异分析
荧光定量PCR 的扩增曲线符合标准的“S”形曲线,Nfatc1和β-actin基因的扩增曲线平行性较好,拐点清楚,基线平而无上扬情况。熔解曲线集中有2 个峰,表明Nfatc1和β-actin没有产生引物二聚体,产物单一。Nfatc1基因在毛囊生长期、退化期和相对静止期的mRNA 表达量变化分析结果显示(图2):Nfatc1在毛囊生长期的mRNA 表达量最低;在毛囊退化期的mRNA 表达量显著高于毛囊生长期和相对静止期,是毛囊生长期的3.34 倍;毛囊相对静止期,Nfatc1的mRNA 表达量有所下降,是生长期的2.3 倍。
与毛囊生长期比较,**P<0.01,下同Compared with Anagen,**P<0.01,The same below
2.3 NFATc1 在小鼠毛囊生长周期中的蛋白量差异分析
在小鼠毛囊的生长期、退化期和相对静止期皮肤组织中各期小鼠皮肤中均存在与NFATc1 多克隆抗体结合的蛋白条带,蛋白条带的大小约为120 kDa,通过Image J 软件对蛋白灰度值进行分析得出,NFATc1 在小鼠毛囊生长的3 个时期的表达量存在差异,其蛋白在毛囊退化期的表达量显著高于毛囊生长期和相对静止期(图3)。

图3 NFATc1 在小鼠生长周期中的蛋白表达水平分析Fig.3 NFATc1 protein expression levels in in the hair follicle cycle in mice.
3 讨论
毛发对于人和动物都具有重要的作用。毛囊是毛发数量和生长所依赖的基本组织结构单元[13]。毛囊作为皮肤中的特有器官,毛囊发育形成后,其上半部分在出生后是恒定不变的,称之为“永久部”;而毛囊的下半部分处于循环变化之中,称之为“循环部”。根据毛囊生长过程中的结构特征将其大致分为:毛囊生长期、毛囊退化期和毛囊相对静止期[14]。毛囊的周期性变化呈现为毛纤维的生长和脱落。本研究结果显示:小鼠处于生长期的毛囊形态为毛乳头增大,干细胞被激活,毛根鞘分化增殖,毛干不断增长;进入退化期的小鼠毛囊,毛乳头萎缩,该时期干细胞处于静止状态,根鞘增殖停止,小鼠毛囊退化期为3 天左右;在短暂的退化期后小鼠毛囊进入相对静止期,在该阶段毛发容易脱落,毛囊干细胞被活化,毛囊乳头不断增大,根鞘生长旺盛,新的毛发不断向上生长将旧的毛发顶至脱落,占据旧毛穴[15-16],毛囊进入了下一个毛囊生长周期。毛囊生长周期包括增殖、分化、迁移以及凋亡等一系列过程,并受多种生长因子、细胞因子、激素、神经肽和药物的影响和调控[17]。近年来研究证实:毛囊的再生依赖于毛囊干细胞在毛发周期的相对静止期向生长期过渡过程中的激活[18],这个过程受到内在和环境因素的严格控制。毛囊干细胞位于毛囊的膨大部(Bulge),它可分化产生毛基质的祖细胞,进而分化向上推移,形成毛囊的内根鞘和毛干。随着基质细胞的分化,膨大部的毛囊干细胞在毛囊生长期处于一个短暂的对称自我更新阶段,最后在退化期再次回到静止状态[19]。Hu 等报道,源于毛乳头的Wnt 信号和BMP 信号与毛囊的生长有关,Wnt/β-catenin 信号通路通过激活毛囊干细胞从而在毛生长过程中发挥重要的促进作用,BMP 信号通路抑制毛囊干细胞的激活,而TGF-β2 信号可以拮抗BMP 促进毛囊的再生。这两个信号通路均通过β-catenin、TCF、LEF1 转录因子进而调控相关靶基因的表达来激活或抑制毛囊干细胞的激活[20]。
Blanpain 等在研究中观察到转录因子Nfatc1在毛囊的基底细胞和膨大部的表达存在显著的差异。Yamamoto 等发现,器官移植过程中使用免疫抑制药物环孢霉素A(CSA)后,机体表现为毛发生长加快,并在进一步研究该现象的机制中证实,CSA 抑制了钙依赖磷酸酶的表达,该酶的减少导致Nfatc1 的表达量降低,从而毛发表现为生长加快的现象[21]。大量研究表明,Nfatc1 表达量的降低或是基因缺失,会导致毛发的过度生长。Gafter-Gvili 根据实验结果提出“NFAT 蛋白在毛囊周期中的退化阶段发挥着重要的作用”[22]。本实验对NFATc1 蛋白在小鼠毛囊生长周期中的定位进行了研究,结果显示NFATc1 蛋白主要定位于毛囊的膨大部,在根鞘、皮脂腺和表皮也有表达,且NFATc1 蛋白在毛囊退化期的膨大部呈强阳性表达,NFATc1 蛋白的阳性反应定位于毛囊干细胞的定殖部位吻合,由此得出:NFATc1 蛋白在毛囊干细胞维持静止状态中发挥着重要的作用。Horsley 等在敲除NFATc1 小鼠模型研究中也发现NFATc1 具有维持毛囊干细胞保持静止状态的生物学功能,NFATc1 的缺失会导致毛囊干细胞被激活,毛囊始终处于生长时期,从而表现毛发生长过剩[8]。进一步对NFATc1 在小鼠毛囊生长周期中的基因和蛋白表达量进行分析,荧光定量PCR和western blotting 的结果显示,NFATc1 在毛囊退化期的mRNA 表达量和蛋白表达量最高,且显著高于在毛囊生长期和相对静止期的表达量。
4 结论
本研究对NFATc1 在小鼠毛囊生长周期中的组织定位和基因蛋白表达量差异进行分析,结果证明,NFATc1 的表达量与毛囊干细胞的活性呈负相关,该因子具有抑制小鼠毛囊干细胞的激活的作用,在小鼠毛囊的周期性生长过程中发挥着重要的作用,为全面揭示毛发周期性生长的分子调控机制提供了理论支持。
猜你喜欢
军事文摘(2023年2期)2023-02-17 09:20:24
感染、炎症、修复(2021年1期)2021-07-28 06:18:08
中国生殖健康(2019年11期)2019-01-07 01:27:32
青少年科技博览(中学版)(2017年5期)2018-02-28 21:23:59
中成药(2017年9期)2017-12-19 13:34:27
CHINESE JOURNAL OF AERONAUTICS(2017年1期)2017-11-21 12:54:14
中成药(2017年5期)2017-06-13 13:01:12
保健与生活(2016年1期)2016-04-12 18:29:44
中国医疗美容(2015年2期)2015-07-19 10:11:59
家畜生态学报(2013年9期)2013-11-30 04:43:30
