孕酮和骨化三醇通过miR-590-5p靶向RelA调控子宫内膜癌细胞Ishikawa的细胞因子分泌
2022-08-30 10:20:44赵丽洁林丽红安阳职业技术学院安阳455000
中国免疫学杂志 2022年11期
赵丽洁 赵 慧 郭 丹 林丽红 (安阳职业技术学院,安阳 455000)
细胞因子/趋化因子是癌症相关炎症的关键因素,越来越多的证据表明,肿瘤细胞产生的趋化因子是其转移介质[1]。NF-κB 是一种强大的促炎调节剂,在肿瘤中被组成激活,并将炎症和致癌作用联系起来[2]。NF-κB 控制多种功能400 余个基因的表达,这些功能包括炎症、生长、凋亡、侵袭和转移[2]。其通常在细胞中处于静止状态,并且仅在响应压力信号(例如促炎性细胞因子)时被激活[2]。NF-κB 由一系 列亚 基组 成,包括RelA、RelB、c-Rel、p50、p52[2]。子宫内膜癌是常见的妇科恶性肿瘤[3]。炎症过程伴随着月经周期内膜的正常生长,脱落和修复,这由性激素控制,也可能由炎症介质自身控制[4]。此外,慢性炎症可能通过内在(促进肿瘤)及外在(引发肿瘤)癌症途径在子宫内膜癌发展中发挥重要作用[5]。已有研究表明孕酮和骨化三醇抑制子宫内膜癌细胞生长和细胞因子分泌[6-7]。但孕酮和骨化三醇抑制子宫内膜癌细胞因子分泌的分子机制尚不清楚。因此,本研究以子宫内膜癌细胞Ishikawa 为研究对象,旨在探讨孕酮和骨化三醇处理Ishikawa细胞减少细胞因子分泌的分子机制。
1 材料与方法
1.1 材料 子宫内膜癌细胞(Ishikawa)购自广州速研生物科技有限公司(货号:Ishikawa)。RelA、RelB、c-Rel、p50、p52、GAPDH 抗体购自Abcam 公司(货 号:ab16502、ab33907、ab133251、ab133492、ab264236、ab8245)。胎牛血清购自广州市左克生物科技发展有限公司(货号:HQ30071)。孕酮购自上海贤鼎生物科技有限公司(货号:PG427-25g)。第一链cDNA 合成试剂盒购自北京兰博利德商贸有限公司(货号:C1802)。骨化三醇购自广州市左克生物科技发展有限公司(货号:C3078-1MG)。青链霉素混合液(100×)购自上海亿言生物科技有限公司(货号:SL6040-100ml)。hsa-miR-584-3p、hsa-miR-1251-3p、hsa-miR-590-5p、hsa-miR-4455、hsa-miR-3613-3p、hsa-miR-7977、hsa-miR-4468、hsa-miR-5093、hsa-miR-939-3p、hsa-miR-610、hsa-miR-7850-5p、hsa-miR-6134、hsa-miR-4789-5p、hsa-miR-493-3p的mimics 和qPCR 引物均由北京擎科合成。BCA 蛋白定量试剂盒购自北京迈瑞达科技有限公司(货号:M052460-500T)。siRelA 由吉凯基因合成。转染试剂购自广州一科生物科技有限公司(货号:BB-4601-2)。RelA 基因从Ishikawa cDNA 中扩增并插入pCDNA3.1,记为pCDNA3.1-RelA。pEZX-MT01购自BioVector 质粒载体菌种细胞基因保藏中心(货号:pEZX-MT01)。RT2 First Strand 试剂盒购自昆明皇宝商贸有限公司(货号:330404)。荧光素酶标记试剂盒购自广州一科生物科技有限公司(货号:A1001-8)。DMEM/F12培养基购自广州泰勒生物科技有限公司(货号:SH30023.01)。Trizol购自派瑞金(北京)生物科技有限公司(货号:T8361)。CXCL1 ELISA 试剂盒购自上海群己生物科技有限公司(货号:KA3337)。CXCL2 ELISA 试剂盒购自广州徕智生物科技有限公司(货号:BS-E6201H1)。TNF-α ELISA 试剂盒购自广州一科生物科技有限公司(货号:EK0584)。IL-1β ELISA 试剂盒购自北京莱科百奥生物技术有限公司(货号:JL29721)。
1.2 方法
1.2.1 细胞培养与转染 Ishikawa 细胞于含10%FBS 及1%P/S 的DMEM/F12 培 养 基 中,37 ℃、5%CO2培养。分别用25 μmol/L 孕酮、100 μmol/L 骨化三醇处理Ishikawa细胞72 h,进行下游实验。hsamiR-584-3p、hsa-miR-1251-3p、hsa-miR-590-5p、hsamiR-4455、hsa-miR-3613-3p、hsa-miR-7977、hsa-miR-4468、hsa-miR-5093、hsa-miR-939-3p、hsa-miR-610、hsa-miR-7850-5p、hsa-miR-6134、hsa-miR-4789-5p、hsa-miR-493-3p 的mimics、miR-590-5p inhibitor、siRelA及pCDNA3.1-RelA 与转染试剂以1∶3 比例混合,室温静置15 min,缓慢滴入Ishikawa细胞。
1.2.2 RNA 抽取与实时荧光定量PCR RNA 提取试剂盒从对照,分离Ishikawa 细胞总RNA。DNase处理后,通过RT2 First Strand 试剂盒按说明合成cDNA。按说明应用7500RT-PCR 系统(AB Applied Biosciences)定量逆转录酶PCR(RT-PCR)。采用2-ΔΔCt法确定相对基因表达。
1.2.3 荧光素酶报告实验 荧光素酶报告质粒pEZX-MT01 包含RelA 野生型或突变型3'UTR。采用荧光素酶报告基因系统分析荧光素酶活性前,分别将miR-590-5p 及阴性对照(NC)与荧光素酶报告基因共转染至Ishikawa细胞(1×104个)48 h。
1.2.4 酶联免疫吸附实验 25 μmol/L 孕酮及100 μmol/L骨化三醇分别处理Ishikawa细胞72 h,测定CXCL1、CXCL2、TNF-α、IL-1β 分泌水平。收获Ishikawa 细胞上清。通过BCA 蛋白测定上清液总蛋白浓度,应用相应ELISA 试剂盒一式两份定量靶蛋白表达。
1.2.5 免疫印迹 应用针对NF-κB 亚基RelA、RelB、c-Rel、p50、p52 的抗体分析孕酮、骨化三醇处理以及未处理的Ishikawa 细胞提取物。对等量蛋白质进行SDS-PAGE 电泳,分析细胞内GAPDH 为上样对照。按照制造商(Pierce)建议,增强后化学发光系统用于可视化蛋白质条带。采用Image J 软件对免疫印迹条带结果进行定量分析。
1.2.6 RNA-Seq 通过Trizol法抽取孕酮和骨化三醇处理后Ishikawa 细胞总RNA、未处理的Ishikawa细胞总RNA。样品送至北京基因组研究所(中国深圳)进行商业性RNA-Seq 服务和数据分析。应用SOAPaligner/SOAP2 不匹配,将明确读段映射到human 数据库。计算每个基因纯净读数数,将其标准化为每百万分之一读数(TKM),该读数将读数数与基因表达相关。利用log2 比率确定基因调控,选择log2 比率为-1或log2比率为1的差异基因进一步分析。
1.3 统计学分析 所有数据采用SPSS18.0 处理。数据以表示,统计学比较采用双尾学生t检验。P<0.05表示差异有统计学意义。
2 结果
2.1 孕酮和骨化三醇抑制Ishikawa 细胞细胞因子IL-1β、TNF-α、CXCL1、CXCL2 分泌 孕酮和骨化三醇分别处理Ishikawa 细胞后,酶联免疫吸附实验发现细胞因子IL-1β、TNF-α、CXCL1、CXCL2 分泌水平明显下降(P<0.05,图1A~D),实时荧光定量PCR 发现细胞因子IL-1β、TNF-α、CXCL1、CXCL2 的mRNA表达均显著降低(P<0.05,图1E)。
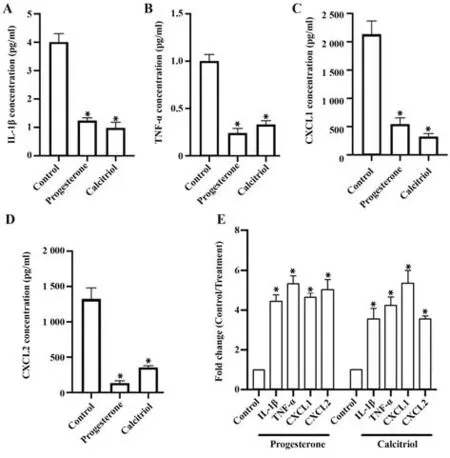
图1 孕酮和骨化三醇抑制Ishikawa 细胞细胞因子IL-1β、TNF-α、CXCL1、CXCL2分泌Fig.1 Progesterone and calcitriol inhibit secretion of cytokines IL-1β,TNF-α,CXCL1 and CXCL2 in Ishikawa cells
2.2 孕酮和骨化三醇在mRNA 水平下调NF-κB 亚基RelA 表达 孕酮和骨化三醇分别处理Ishikawa细胞后,免疫印迹发现NF-κB 亚基RelA 表达下调,NF-κB 亚基RelB、c-Rel、p50、p52 表达无明显改变(图2A、B)。定量结果见图2C。转染RelA promoter-luciferase 质粒后,应用孕酮和骨化三醇分别处理Ishikawa 细胞,发现其荧光素酶报告活性变化差异无统计学意义(P>0.05,图3A),实时荧光定量PCR发现NF-κB 亚基RelA mRNA 水平降低(P<0.05,图3B)。

图2 孕酮和骨化三醇处理后NF-κB亚基表达Fig.2 Expression level of NF-κB subunit after progesterone and calcitriol treatment

图3 孕酮和骨化三醇处理后NF-κB 亚基RelA 转录水平及启动子活性Fig.3 Transcription level and promoter activity of NF-κB subunit RelA after treatment with progesterone and calcitriol
2.3 miR-590-5p靶向NF-κB亚基RelA的-3'端非编码区 使用孕酮和骨化三醇分别处理Ishikawa 细胞后,通过高通量测序发现孕酮处理后有110 种mi-RNAs表达上调,而骨化三醇处理后有57种miRNAs表达上调(图4)。两组均上调者有14种miRNAs,分别 是hsa-miR-584-3p、hsa-miR-1251-3p、hsa-miR-590-5p、hsa-miR-4455、hsa-miR-3613-3p、hsa-miR-7977、hsa-miR-4468、hsa-miR-5093、hsa-miR-939-3p、hsa-miR-610、hsa-miR-7850-5p、hsa-miR-6134、hsamiR-4789-5p、hsa-miR-493-3p(图5)。通过miRNA mimics 表达上述miRNA 后,发现miR-590-5p 可降低NF-κB 亚基RelA mRNA 水平(P<0.05,图6A)。通过荧光素酶报告实验发现miR-590-5p 靶向NF-κB亚基RelA-3'端非编码区(P<0.05,图6B)。同时,通过miR-590-5p mimic 过表达miR-590-5p 后,NF-κB亚基RelA蛋白水平降低。通过miR-590-5p inhibitor敲低miR-590-5p 后,NF-κB 亚基RelA 蛋白水平升高(图6C、D)。定量结果见图6E。
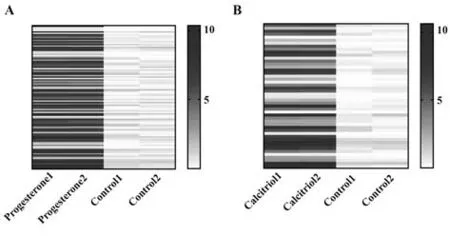
图4 孕酮和骨化三醇处理后Ishikawa细胞的高通量测序Fig.4 High-throughput sequencing of Ishikawa cells treated with progesterone and calcitriol

图5 孕酮和骨化三醇处理Ishikawa细胞的高通量测序结果交集Fig.5 Intersection of high-throughput sequencing results of progesterone and calcitriol treated Ishikawa cells
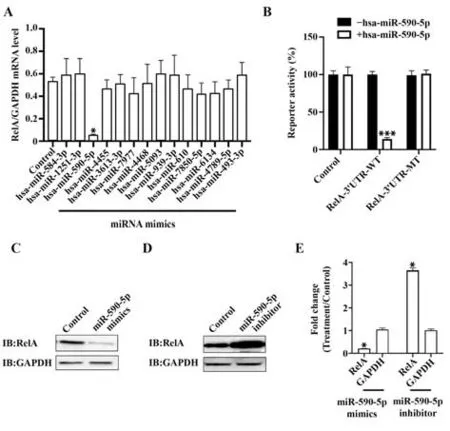
图6 miR-590-5p靶向NF-κB亚基RelA的-3'端非编码区Fig.6 miR-590-5p targets 3' terminal non-coding region of NF-κB subunit RelA
2.4 miR-590-5p靶向NF-κB信号通路调控Ishikawa细胞细胞因子分泌 miR-590-5p mimic 过表达miR-590-5p后,细胞因子IL-1β、TNF-α、CXCL1、CXCL2表达均显著降低(P<0.05)。miR-590-5p inhibitor 敲低miR-590-5p 后,细胞因子IL-1β、TNF-α、CXCL1、CXCL2 表达均显著升高(P<0.05)。但同时过表达或 敲 低miR-590-5p 和RelA 时,细 胞 因 子IL-1β、TNF-α、CXCL1 和CXCL2 表达无明显变化(P>0.05,图7)。
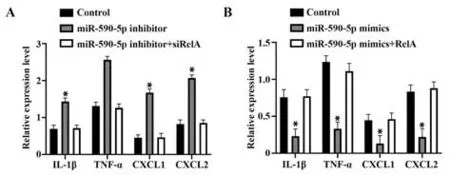
图7 miR-590-5p 靶向NF-κB 信号通路调控Ishikawa 细胞细胞因子分泌Fig.7 miR-590-5p targets NF-κB signaling pathway to regulate secretion of cytokines in Ishikawa cells
3 讨论
本研究发现孕酮和骨化三醇降低Ishikawa 细胞中细胞因子IL-1β、TNF-α、CXCL1、CXCL2 的表达和分泌。越来越多的证据支持炎症能够加速多种癌症发展和进展[8]。子宫内膜癌是由炎症驱动的疾病,涉及复杂细胞因子和趋化因子网络激活[9]。最近的研究表明骨化三醇和孕酮具有抗肿瘤作用[10],可能是得益于其抑制细胞因子表达和分泌。
临床、流行病学和基础科学证据表明,骨化三醇可能是高效的卵巢癌和子宫内膜癌预防剂,可直接调节许多细胞中细胞因子诱导的NF-κB 活性[11-12]。亦可抑制乳腺和前列腺细胞中NF-κB 信号传导,降低血管生成及促炎性细胞因子IL-8 的水平[13]。孕酮是女性生殖道一种强大的抗炎和抗增殖激素[14]。在分化较差的子宫内膜癌细胞Hec50c中,孕酮显示 可 以抑制NF-κB 转录活性[15]。但GEISERT 等[16]发现孕酮可以激活子宫上皮NF-κB转录活性并增加细胞因子表达。提示孕酮与NF-κB转录活性具有较高相关性。基于此,本研究使用骨化三醇和孕酮处理后,检测NF-κB 各个亚基表达,发现NF-κB 亚 基RelA 表达 下 调,而NF-κB 亚 基RelB、c-Rel、p50、p52 表达无明显改变。因此,孕酮和骨化三醇的作用靶点可能是RelA。孕酮和骨化三醇分别处理Ishikawa 细胞,RelA 启动子活性变化差异无统计学意义(P>0.05),但RelA mRNA水平降低(P<0.05)。提示孕酮和骨化三醇在mRNA 水平降低RelA 表达。miRNA 是一个小型非编码RNA 分子,包含约22 个核苷酸,在RNA 沉默和基因表达转录后调控中起作用[17]。凭借这些特性,miRNA 可以调节参与癌症发展的多种信号传导途径,例如细胞增殖、凋亡和迁移[18]。通过高通量测序发现孕酮和骨化三醇分别处理Ishikawa细胞后,有110种miRNAs表达上调,骨化三醇处理后有57 种miRNAs 表达上调,其中均上调的有14种miRNAs。在这些miRNAs中仅有miR-590-5p 可降低NF-κB 亚基RelA mRNA水平(P<0.05)。但其他被孕酮和骨化三醇调节的miRNA 是否在抗肿瘤中发挥其他功能值得进一步探索。虽然本研究和GEISERT 等[16]的研究对象都是子宫细胞,但GEISERT 等人应用的模式生物是猪,人和猪的NF-κB 并非完全同源,两个物种蛋白表达调控机制并不相同,且猪无miR-590-5p 表达,因此孕酮可能导致人和猪产生不同的NF-κB 转录活性。
最近miRNA 被引入,有希望成为癌症治疗靶标。miRNA 有抑制肿瘤的调节潜力,可调节细胞内整个信号网络,这使其成为开发癌症治疗剂的选择之一[19]。miR-590-5p 可消除细胞因子,有望成为治疗子宫内膜癌的新miRNA工具。
综上所述,孕酮和骨化三醇处理Ishikawa 细胞后miR-590-5p 表达上升,miR-590-5p 靶向NF-κB 亚基RelA 的-3'端非编码区,降低RelA 蛋白水平,降低NF-κB 复合体活性,最终下调细胞因子IL-1β、TNFα、CXCL1、CXCL2的表达。
猜你喜欢
中国农业科技导报(2023年10期)2023-11-02 07:48:50
蜜蜂杂志(2022年5期)2022-07-20 09:54:16
皮肤病与性病(2021年3期)2021-07-30 08:07:58
海峡科技与产业(2021年1期)2021-05-22 01:26:20
中国民间疗法(2021年1期)2021-04-20 02:31:08
河北医学(2016年12期)2017-01-05 01:49:26
知识就是力量(2016年8期)2016-11-02 01:00:11
实用中医药杂志(2015年2期)2015-12-01 07:06:28
云南中医学院学报(2015年1期)2015-07-31 18:10:45
中国当代医药(2015年17期)2015-03-01 02:03:51