
高原鼠兔刈割对青海湖流域高寒草甸植物种间关联的影响
2022-08-24 05:50齐昊昊庞晓攀周俗郭正刚
草业学报 2022年8期
齐昊昊,庞晓攀,周俗,郭正刚*
(1. 兰州大学草地农业生态系统国家重点实验室,兰州大学草地农业科技学院,甘肃 兰州 730020;2. 四川省林业科学研究院,四川 成都 611731)
高原鼠兔(Ochotona curzoniae)是青藏高原高寒草地生态系统的重要组分[1],主要营家族式生活,一个家族往往集体生活在其巢域范围内[2]。高原鼠兔通过采食、掘洞、覆土、排泄、刈割[3-4]等多种行为干扰高寒草甸,因此,高原鼠兔干扰区和非干扰区植物群落特征的差异,成为阐明高原鼠兔干扰对高寒草甸整体影响的基本途径[5-6],但无法解释单一行为对高寒草地的影响,而高原鼠兔单一行为对高寒草甸的影响,则是分析整体影响的基础单元。刈割是高原鼠兔特有的生存策略[7],主要指高原鼠兔经常会咬断洞口附近和迁移通道的高大植物,以便从茂密草丛中创建开阔的视野,快速发现天敌而迅速逃离危险区[8],从而实现规避捕食风险[7,9],然而这种刈割行为必然会抑制刈割区内被刈割植物的生长,释放刈割区内低矮植物的生长潜势[10-11]。因此,高原鼠兔刈割行为改变了原有植物群落内高大植物和低矮植物间的关系,这不仅会影响高寒草甸植物群落内的种间关系,而且会改变刈割区内原有的植物群落结构。虽然高原鼠兔刈割行为仅能够改变小区域内(刈割区内)的植物群落结构,但高原鼠兔分布面积大,范围广[12],其刈割的点面交织后,成为青藏高原高寒草地植物群落结构自我调节的重要途径。因此,研究高原鼠兔刈割行为对高寒草地植物群落结构的影响,是全面解读高原鼠兔干扰影响高寒草甸植物群落结构的重要内容,也是揭示高原鼠兔驱动植物群落结构变化的途径之一。
合理的研究方法是分析高原鼠兔刈割行为影响高寒草地植物群落结构的基础。高原鼠兔被证实在整个巢域内进行选择性采食[13]。高原鼠兔造就的裸斑和建立的洞道系统因无植被覆盖或植被稀疏[8],一般不会发生刈割现象;高原鼠兔粪尿主要排泄于有效洞口和废弃洞口附近[2-3],而废弃洞口因没有高原鼠兔生活,亦不会发生刈割行为。高原鼠兔巢域内有效洞口和废弃洞口往往呈现镶嵌分布[14],两种洞口附近的土壤类型和气象因子基本一致,因此,通过分析有效洞口附近和废弃洞口附近植物群落的差异,成为量化研究高原鼠兔刈割行为与植物群落结构关系的主要途径[4]。Zhang 等[4]采用此方法证实,高原鼠兔刈割对群落生物量无明显影响,但降低了植物群落高度和禾草类的生物量,增加了植物群落物种丰富度和非禾草类生物量[4],这说明高原鼠兔刈割行为虽然不会影响刈割区内的植物生产力,但高原鼠兔刈割对禾草类植物和非禾草类植物有明显影响,这可能会影响禾草类植物之间以及与其他非禾草类植物之间的种间关系,从而影响植物群落的结构。
种间关联被大量研究用于测度植物的种间关系,常被用来分析植物群落的稳定性[15-16],其主要包括种间联结性和种间相关性。种间联结性包括总体联结性和种对间联结性[17],其中植物群落总体联结性可以反映植物群落的稳定性[15,18],当其处于正向时,说明植物群落内物种间互惠互生、协同共存,植物群落稳定性相对较高,反之,则表明植物群落相互竞争,相对不稳定,处于变化状态[19];种对间联结性则表征两物种之间的相互作用,种对间正关联表示对一方或者双方是有利的,种对负关联可能表明不利于一方或对双方都不利[16]。然而,无论是种间总体联结性,还是种对间的联结性,其均以物种存在与否为基础界定物种间是否存在关联[20],虽然可测度种对联结的性质和显著程度[21],但没有考虑植物种群的生物学信息[22],而种间相关性虽然考虑物种存在与否[20]和物种的生物学信息(多度、盖度和重要值等)[23],但无法判别种间联结的性质。因此,采用种间联结性和相关性相结合的方法,成为分析草食动物干扰对草地植物群落种间关联性影响的途径。目前已有研究采用种间联结性和相关性相结合的方法,解读了大型放牧动物对科尔沁沙地植物群落物种联结特征的影响[16],然而大型放牧动物和小型草食动物干扰草地的方式存在分异,那么种间联结性和相关性相结合的方法,能否测度小型草食动物对草地植物种间关联特征的影响,尚需要科学试验提供证据。
青海湖流域是青藏高原西部荒漠风沙向东扩展的主要生态屏障,其中草地占整个流域面积的比例高达72%,是流域涵养水源、防风固沙和防止水土流失的重要载体[24]。高山嵩草(Kobresia pygmaea)草甸是青海湖流域高寒草地的重要类型之一[5],其草丛低矮,是高原鼠兔的主要生活场所[8]。因此,本研究以青海湖流域内高山嵩草草甸为载体,采用种间联结性和相关性相结合的方法,研究高原鼠兔刈割行为对高寒草甸植物种间关联特征的影响,完善草食动物干扰(大型和小型)与植物种间关系的研究内容,为全面解读高原鼠兔干扰驱动植物群落结构变化提供基础信息。
1 材料与方法
1.1 研究区概况
研究区位于青海湖流域内刚察县的哈尔盖镇(100°30′30″E,37°23′17″N),属典型高原大陆性气候,具有寒冷潮湿的特点[5]。刚察县平均海拔3480 m,年平均气温-0.6 ℃,年平均降水量300~500 mm,主要集中于植物生长季的6-9 月,年平均蒸发量1500~1847 mm。植被类型以高寒草甸为主,主要包括高山嵩草草甸、矮生嵩草(K. humilis)草甸、线叶嵩草(K. capillifolia)草甸等类型,这些高寒草甸被牧户承包,一般划分为冷季放牧草地和暖季放牧草地。每户的冷季放牧草地一般采用围栏封育,既是边界,又是植物生长季休牧的主要手段,而牧户有的暖季放牧地有些有围栏,有些没有围栏,属于多户共享状态[4]。高山嵩草草甸因草丛相对低矮而具有开阔的视野,有利于高原鼠兔发现天敌,因此,高山嵩草草甸是高原鼠兔栖息的主要草地类型之一,反之也是高原鼠兔最易致灾的草地类型[6]。高山嵩草草甸的优势种为高山嵩草,主要伴生种有长毛风毛菊(Saussurea hieracioides)、高原早熟禾(Poa alpigena)和高山豆(Tibetia himalaica)等。
1.2 试验设计
选择冷季放牧草地作为样地布设地,可以利用牧户围栏,排除植物生长季内大型放牧动物的影响。1)2019年4 月,赴青海刚察县选择调查区域,根据目测有无高原鼠兔出没现象,选定刚察县哈尔盖镇公贡麻村一户牧户的冷季放牧草地为调查区域,调查区域的面积约为105 m×350 m,以便合理布设样地;2)2019 年8 月赴已经选好的调查区域,将调查区域划分为3 条样地带,每条样地带10 个样地,鉴于高原鼠兔巢域平均面积1262.5 m2[25],确定样地面积为35 m×35 m,30 个样地呈连续分布状态;3)随机选择一个样地,测定该样地内所有洞口的直径,求其平均值,作为遴选有效目标洞口和废弃目标洞口的依据,洞口的平均直径为0.125 m;4)利用连续3 d 干草堵洞法判别所有样地内每个洞口属于有效洞口或废弃洞口,逐一标记[26-27];5)以0.125 m 为标准,每个样地内采用配对法确定有效目标洞口和废弃目标洞口,随机选择一个符合洞口直径要求的有效洞口,然后在该洞口附近选择一个符合洞口直径要求的废弃洞口,形成配对。整体试验包括60 个洞口,30 个有效洞口和30 个废弃洞口。
1.3 样方调查
高原鼠兔刈割高大植物的范围基本呈现为一个直径为0.5 m 的圆形区域[4]。高原鼠兔每个洞口前方均有一个无植被或植被稀疏的裸斑(图1),将裸斑所在方向定位为洞口的前方,然后确定该洞口的后方、左右方。针对每个洞口,放置一个0.5 m×0.5 m 的大样方,再在洞口位置设置一个0.15 m×0.15 m 的小样方,大、小样方中心和洞口中心重合,且与洞口裸斑中心在同一条直线上,这样在洞口的后方、左右方,小样方和大样方间会形成3 个梯形区域,将这3 个梯形区域作为一个植物群落调查区,其面积约为0.1706 m2[1/2×(0.15+0.5)×0.175×3]。首先记录每个植被调查区域内的所有维管束植物种类,然后测定每个种群的密度、盖度和高度。不同植物种群密度采取不同测定方法,单子叶植物以植被调查区内出现的一级分枝数为基准测定,双子叶植物以植被调查区内出现的总株数计算[28]。盖度采用针刺法测定,从样方右侧边缘开始针刺,横竖方向每次隔2.5 cm 刺一下,一共刺273 下。植物种群高度测定,若某种植物在植被调查区出现的株数超过30 株,则随机选择30 株,以其平均值为该植物的株高;若某种植物在植被调查区内的株数不够30 株时,则全部测量其株高,以其平均值作为该植物的株高。
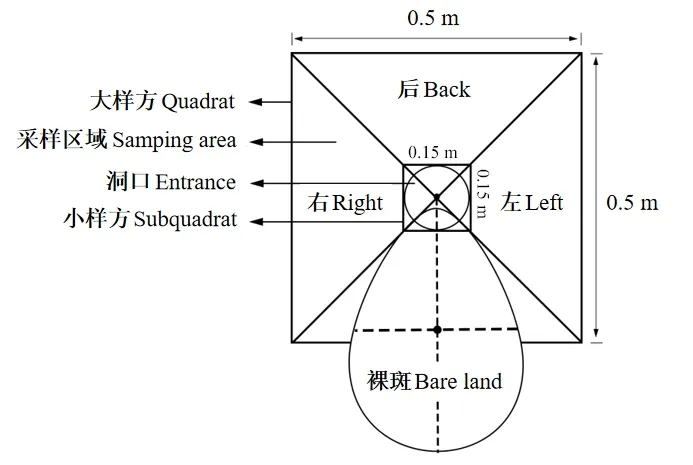
图1 高原鼠兔洞口取样示意图[4]Fig.1 Sampling of a plateau pika burrow entrance
1.4 指标计算
1.4.1目标植物选择 研究地区高寒草甸的物种数较多,但某个物种出现于植被调查区的概率不同。为了便于计算植物种间的关联性和相关性,需要从调查的植物种类中选择适宜分析种间关联性和相关性的物种。首先,遴选30 个有效洞口和30 个废弃洞口植被调查区内共同出现的物种作为初选目标植物;然后计算每一个初选目标植物的平均重要值,根据平均重要值大小,再次遴选物种。植物重要值(importance value,IV)=(HR+CR+DR)/3,其中IV为某一物种的重要值,HR为某一物种的相对高度,CR为某一物种的相对盖度,DR为某一物种的相对密度。当某一初选目标植物的平均重要值>1%时,该物种纳入种间关联分析,若某一初选目标植物的平均重要值≤1%,则被认为该物种对植物群落结构影响不大,不被纳入种间关联分析,最终选择了25 个物种(表1),作为种间关联分析的物种,这些物种基本涵盖了优势种和主要伴生种。

表1 高山嵩草草甸种群25 个物种的平均重要值Table 1 The mean importance value of 25 main species in a K.pygmaea meadow populations(%)
1.4.2多植物种间的总体联结性检验 群落内植物种间总体联结性采用Schluter 方差比率法(VR)检验[29],计算公式如下:

式中:VR值为群落内种间的总体联结指数为所有样方物种数的方差为所有物种出现频度的方差;N为总样方数;S为总物种数;Tj为第j个样方内的物种总数;t为样方内物种的平均值;nk为物种k出现的样方数;Pk为物种k出现的样方数占总样方数的比值。当VR=1 时,表示种间总体无联结;当VR>1 时,表示种间总体联结性为正关联;当VR<1 时,表示种间总体联结性为负关联。测定的总体联结指数利用统计量W(W=VR×N)检验VR值偏离1 的程度是否显著,若W落入χ2界限内则种间总体联结性不显著(P>0.05),反之,若,则种间总体联结性显著(P<0.05)。
1.4.3植物种对间关联性 采用χ2检验评价植物种对间的关联性和显著程度。χ2统计量基于抽样的非连续性,χ2值使用Yates 连续校正方法计算[20],其公式为:

式中:a是两个物种都出现的样方数;b、c分别是仅出现物种1 或物种2 的样方数;d是两个物种都不出现的样方数。N为总的样方数。若χ2>6.635(P<0.01),可认为种对间具有极显著联结;若3.841≤χ2≤6.635(0.01≤P≤0.05),可认为种对间具有显著联结;若χ2<3.841(P>0.05),可认为种对联结不显著。当ad>bc时种对间有正联结,当ad<bc时种对间有负联结,当ad=bc时种对间无关联。若某一物种的频度为100%时,b、d值均为0,无法进行χ2检验,可以令式中的b、d值均为1,以便客观地检验2 个种的关联[17]。
1.4.4种间联结程度 种间联结程度用联结系数(association coefficient,AC)和Ochiai index(OI指数)度量,其中AC值的计算分以下3 种情形:
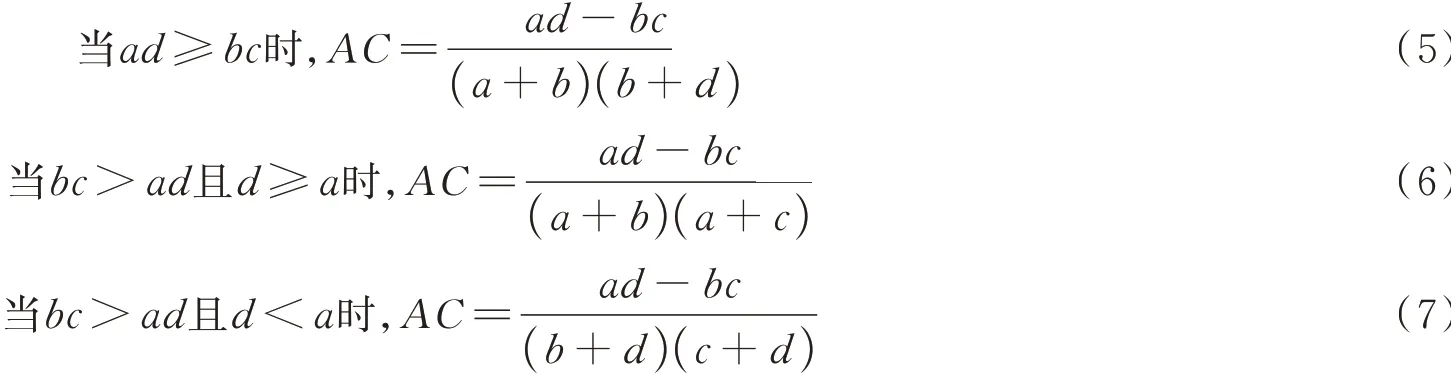
式中:种间联结系数AC的值域为[-1,1]。AC值越趋近于1,表明物种间的正联结性越强;而AC值越趋近于-1,表明物种间的负联结性越强;AC值为0,则物种间完全独立。

式中:OI表示种对同时出现的几率和联结性程度,可以克服AC值受d值影响较大造成的偏差。该指数在无关联时为0,在最大关联时为1[30]。
1.4.5植物种间相关性 应用定量数据(重要值)对种间关系进行Spearman 秩相关分析,首先将重要值数据秩化,将重要值变换为秩化向量,然后将秩化向量代入相关系数公式,得到的值称为Spearman 秩相关系数[r(i,k)][20],计算公式如下:

式中:dj=(xij-xkj);N为样方数;xij和xkj分别为物种i和k在样方j中的秩。
2 结果与分析
2.1 高原鼠兔刈割行为对植物群落总体联结性的影响
对照区植物种间总体联结性的方差比率(VR)为1.09,大于1(表2),检验统计量W为32.70,落入界限,其种间总体联结性呈不显著正关联。高原鼠兔刈割后,植物种间总体联结性的VR为0.61,小于1,检验统计量W为18.30,没有落入界限,其植物种间总体联结性呈显著负关联(P<0.05),说明刈割行为使得高寒草甸植物群落结构变得不稳定。

表2 群落总体联结性变化Table 2 Dynamics of the overall interspecific association in K.pygmaea meadow
2.2 高原鼠兔刈割行为对植物群落种对间联结性的影响
χ2检验结果表明,高原鼠兔刈割行为对植物群落种对的联结性有明显影响(图2),共有的300 个种对在高原鼠兔刈割后有146 个植物种对间的联结性或显著性发生了改变,正关联种对比例由对照区的46.33%变为刈割区的40.33%,负关联种对比例由对照区的50.00%变为刈割区的54.00%,无关联种对比例由对照区的3.67%变为刈割区的5.67%。其中,种对7-24(紫花针茅-三脉梅花草)和种对3-17(高原早熟禾-达乌里秦艽)的联结性分别由对照区的极显著正关联、显著正关联转变为刈割后的不显著正关联,种对8-14(直梗高山唐松草-二裂委陵菜)和种对10-13(乳白香青-小花草玉梅)的联结性均由对照区的不显著负关联转变为刈割后的显著负关联。

图2 χ2检验半矩阵Fig.2 Semi-matrix of interspecific association χ2 correction test
2.3 高原鼠兔刈割行为对植物群落种对间联结程度的影响
AC值检验结果发现,高原鼠兔刈割后群落内正关联种对数降低,负关联和无关联种对数增加(图3),这与χ2检验结果一致。废弃洞口附近和有效洞口附近的植物种对的AC值在(-0.6,0.6)范围内的种对比例分别为81.00%、85.00%,反映大部分种对联结程度中等偏弱。然而,部分种对间AC值在高原鼠兔刈割后发生了变化,例如高山嵩草、长毛风毛菊和高山豆与高原早熟禾的AC值均由废弃洞口附近的0 变为有效洞口附近的(-0.3,0),说明其联结程度由无关联变为较弱的负关联。OI指数可以克服联结系数AC值受d值影响误差偏大的问题,结果表明,废弃洞口附近和有效洞口附近的种对的OI值在(0,0.8)范围内的种对比例分别为85.67%、83.33%(图4),说明大部分的植物种对在废弃洞口附近和有效洞口附近的关联程度中等偏弱,这与AC值检验结果一致。然而,高山嵩草、长毛风毛菊和高山豆与高原早熟禾的OI指数在废弃洞口附近和有效洞口附近没有明显变化,均在[0.6,0.8)范围内,说明其关联程度为中等,该结果与AC值检验结果相异。

图3 联结系数(AC)值半矩阵Fig.3 The semi-matrix of association coefficients

图4 Ochiai 指数(OI)半矩阵Fig.4 The semi-matrix of Ochiai index(OI)
2.4 高原鼠兔刈割行为对植物群落种间相关性的影响
Spearman 秩相关分析结果发现,正相关种对比例由废弃洞口附近的83.66%变为有效洞口附近的74.66%(图5),负相关种对比例由废弃洞口附近的16.33%变为有效洞口附近的25.33%,且种对间相关性显著水平明显高于χ2检验。其中,2-20(长毛风毛菊-矮火绒草)和22-23(西伯利亚蓼-蒲公英)种对间相关性由极显著正相关变为不显著负相关,10-22(乳白香青-西伯利亚蓼)、13-21(小花草玉梅-珠芽蓼)、15-21(雪地黄耆-珠芽蓼)、21-23(珠芽蓼-蒲公英)种对间相关性由显著正相关变为不显著负相关,1-9(高山嵩草-黄花棘豆)和1-11(高山嵩草-莓叶委陵菜)种对间相关性由不显著负相关变为极显著正相关,3-9(高原早熟禾-黄花棘豆)种对间相关性由不显著正相关变为显著负相关。

图5 Spearman 秩相关系数半矩阵Fig.5 Semi-matrix of Spearman’s rank correlation coefficients
3 讨论
种间联结是指不同物种在空间分布上的相互关联性,反映了植物群落内各个物种在适应生境过程中利用种间吸引或排斥形成的有机联系[15-21],而群落稳定性是种间联结性的表达方式之一[18],刈割作为高原鼠兔适应生境和保持生存的特殊策略,具有提高高原鼠兔生存能力的功效,本研究结果表明,高原鼠兔刈割行为迫使高寒草甸植物种间总体联结性从废弃洞口附近的不显著正关联,转变为有效洞口附近的显著负关联,说明高原鼠兔刈割行为减弱了高寒草甸植物群落的稳定性,主要机理是高原鼠兔刈割改变了植物群落内不同植物利用光照资源的能力[9-10],给原有的低矮植物提供了更多光资源,从而使原有高大植物主导利用光照资源的格局被打破,植物群落中原有的高大植物变得相对低矮,与原有低矮植物间竞争光资源的能力减弱,此时原有群落内优势种高山嵩草重要性相对降低,其抑制伴生种的作用相对减弱,这为原有劣势竞争潜力的植物提供了更好的生长环境[31-32]。因此,高原鼠兔刈割行为改变了原有植物群落内的物种竞争光资源的关系,迫使植物群落处于变化状态,不稳定性增加,这对全面认知高原鼠兔在高寒草甸中的作用提供了重要信息。
植物群落内正关联种对比例越高,群落结构及种类组成将越趋于完善和稳定[16],反之,则说明植物群落逐渐退化。本研究共有25 个物种参与配对,形成300 个种对,结果表明高原鼠兔刈割会降低正关联种对的数量,而负关联种对数量和无关联种对数的数量则具有增加趋势,这与高原鼠兔刈割对植物群落总体联结性的影响基本一致,这有力地支持了植物群落总体联结性是所有物种种对间的联结性累积而实现的观点[33],说明植物群落的总体联结性是群落内所有物种种对间关联性的综合体现。AC值和OI指数表明,大部分种对间的关联性属于中等偏弱,表明植物群落内大部分种对间的联结性和联结程度不强,关系相对松散,独立性强[15],譬如优势种高山嵩草与主要伴生种长毛风毛菊、披针叶黄华和高山豆之间的联结性和联结程度,在有效洞口附近和废弃洞口附近没有明显变化。而高原鼠兔刈割行为会改变少数种对的联结性和联结程度,譬如优势种高山嵩草与高原早熟禾、二裂委陵菜和西伯利亚蓼的联结性和联结程度从废弃洞口附近到有效洞口附近,发生了明显变化,分别从无关联、不显著正关联、不显著负关联,转变为不显著负关联、不显著负关联、不显著正关联,主要机理是高原鼠兔刈割区内虽然高山嵩草占据优势的主导地位没有发生改变,但其主导植物群落的优势度有所降低,为植物群落内不同物种占据其分化出的生态位提供了更多的资源空间[34],然而不同物种生物学习性存在差异,有些物种与高山嵩草生态位相对接近,而有些物种与高山嵩草的生态位相对分散[33],从而表现为不同物种与高山嵩草的种间联结性和联结程度对高原鼠兔刈割行为的响应存在差异,这说明分析高寒草甸生态系统食物网中高原鼠兔承上启下的作用时[32],还需要关注其对植物群落稳定性的影响,而植物群落处于分化状态时可能更有利于多种植物种共存[15],因此高原鼠兔在生态系统中的作用,主要取决于人们对草地功能的需求。
Spearman 秩相关分析结果表明,高原鼠兔刈割减少了植物群落内正相关种对的数量,增加了植物群落内负相关种对的数量,譬如种对1-3(高山嵩草-高原早熟禾)和3-4(高原早熟禾-高山豆)由废弃洞口附近的不显著负相关,转变为有效洞口附近的显著负相关,种对2-3(长毛风毛菊-高原早熟禾)则从废弃洞口附近的不显著正相关,转变为有效洞口附近的不显著负相关。因此,Spearman 秩相关分析结果与χ2检验结果基本一致,高原鼠兔一般喜欢刈割有效洞口附近的高大植物,由于禾草类植物通常比非禾草类植物高[8],因此禾草类植物更容易成为高原鼠兔刈割的对象[7],这会明显减少禾草类植物获取和利用光资源的能力,反而增加非禾草低矮植物获取和利用光照资源的能力,较废弃洞口附近植物群落而言,高原鼠兔刈割会提高有效洞口附近植物群落内非禾草类植物的竞争潜力,此时迫使高原早熟禾与其他多数伴生种竞争生境中资源的强度增加,结果导致两个物种相互排斥,出现生态位分离,以适应高原鼠兔刈割后的生境变化。然而,采用χ2检验高山嵩草-高原早熟禾和高原早熟禾-高山豆种对的关联性时,其联结性和联结程度并没有发生明显变化,这说明Spearman 秩相关分析和χ2检验对种间关联性检验时,有时结果会不一致,主要原因是χ2检验物种间的联结性时,以植被调查区内是否存在物种的二元数据为标准,损失了相关多度信息[17],而Spearman 秩相关分析基于物种的生物学数据,可以明确植物空间分布的基本现状,检验结果更灵敏[22],是对χ2检验结果的有效补充和完善[15],这说明利用χ2检验和Spearman 秩相关分析的互补特征,能够相对准确、全面地评价高原鼠兔刈割行为对草地植物种间关联的影响。已有研究已经证实,利用χ2检验和Spearman 秩相关分析相结合的方法,能够评价大型放牧动物对植物群落物种关联特征的影响[16],本研究又证明χ2检验和Spearman 秩相关分析相结合,能够评价小型草食动物对植物种间关联的影响,表明利用χ2检验和Spearman 秩相关分析,不仅能够分析大型草食放牧动物对草地植物群落种间关联性的影响,而且能够分析小型草食动物干扰对草地植物群落种间关联性的影响。因此,本研究结果不仅完善了草食动物(大型和小型)干扰与植物种间关联的研究范围,而且确证了种间联结性和相关性相结合能够用于分析小型草食动物干扰对植物群落种间关联的影响。
高原鼠兔刈割行为是高寒鼠兔干扰高寒草甸的重要方式之一[19],也是高原鼠兔维持生存的基本策略[7]。本研究利用方差比率法、χ2检验和Spearman 秩相关分析相结合的方法,研究了高原鼠兔刈割行为对植物群落总体联结性、种对间关联性和种对间相关性的影响,发现高原鼠兔刈割行为改变了植物群落总体联结性,降低了正关联种对的数量,增加了负关联和无关联种对的数量,说明高原鼠兔刈割行为会促进植物群落分化,证实了χ2检验和Spearman 秩相关分析能够评价小型草食动物对植物群落种间关联的影响,这完善了目前草食动物干扰与植物群落种间关系的研究内容。
猜你喜欢
海外文摘·文学版(2021年3期)2021-05-19
当代陕西(2019年15期)2019-09-02
西部资源(2018年3期)2018-11-06
学苑创造·A版(2018年11期)2018-02-01
读者(2017年5期)2017-02-15
延河·绿色文学(2016年8期)2016-05-14
棋艺(2014年7期)2014-09-09
城市建设理论研究(2014年5期)2014-02-18
小小说月刊·下半月(2011年8期)2011-05-14
