
吉林蛟河针阔混交林群落邻体竞争效应*
2022-08-20 06:19:14粟佳琳范春雨张春雨赵秀海
林业科学 2022年5期
粟佳琳 王 娟 范春雨 张春雨 赵秀海
(1.北京林业大学森林资源和环境管理国家林业和草原局重点实验室 北京 100083; 2.北京林业大学生态与保护学院 北京 100083)
近年来,空间点格局已成为群落生态学研究的热点之一。林木空间分布格局是种群相互作用及多种生态过程综合作用的结果(许爱云等, 2020),林木个体的空间布局方式决定了其功能的发挥(Delvaux, 1981)。林木个体间的相互作用也称为邻体效应,包括促进、抑制和无相关作用(del Ríoetal., 2014; 郝珉辉等, 2017a)。邻域竞争是邻体效应的一个重要特征,反映相邻个体在生态位或资源位置上的相互作用(Zhangetal., 2019)。邻体效应受到林木的种类、特征和相对位置影响(Getzinetal., 2008)。例如,种子的扩散模式(Heetal., 1997; Murrell, 2002)、物种间竞争耐受力差异(Coomesetal., 2007)、不同林层间物种组成及生态特征差异(Graveletal., 2010)都会对邻体效应产生影响。
生境异质性指一个区域内不同位置之间的环境差异性,包括地形和生境类型等(马芳等, 2018)。生境异质性对物种空间分布有重要影响(郝珉辉等, 2017b; 王宇等, 2020; Harmsetal., 2001; Zhangetal., 2012; Itohetal., 2010)。点格局分析方法是通过量化不同尺度下的物种空间分布特征来反映物种分布格局类型与空间尺度的关系(樊登星等, 2016; Wälderetal., 2007; Ripley, 1981),在林分结构研究中被广泛应用。以往研究大多从树木的相对多度入手,研究多尺度物种空间分布的关联性(杨华等, 2014; 姚杰等, 2018; 樊登星等, 2016)。相较于相对多度,林木的大小特征能更好地反映环境条件的影响程度(Russoetal., 2008; Benito-Garznetal., 2013),进而揭示生境异质性对邻体效应的潜在作用。
标记点过程为邻木竞争分析提供了一个理论框架(Wälderetal., 2007),其中“点”表示树木的位置,“标记”可用来表示胸径、树高或特定时段的生长特征等(Diggle, 2013)。标记相关函数描述了被观测的点格局中标记属性的空间自相关性,有助于理解林木生长的相互作用效应(Wälderetal., 2007; Diggle, 2013)。点格局分析中完全空间随机零模型(complete spatial randomness)要求生境背景必须同质(Ripley, 1981),但复层异龄天然林中的生境因子多呈异质性分布,不可避免地会对点格局分析带来影响。本研究尝试通过生境划分的方法,将异质性样地划分为2个亚区(每个亚区内部生境相对同质),然后通过标记点格局分析探讨不同生境亚区中林木个体间的相互作用规律,旨在为森林经营作业提供理论支撑。
1 研究区概况
试验区吉林省蛟河林业实验区国有林保护中心(127°42′789″—127°43′310″E, 43°57′897″—43°58′263″N)位于蛟河市前进乡境内,东与吉林省延边朝鲜族自治州相邻,北与黑龙江省五常县接壤,西南部为蛟河。辖区总经营面积31 823 hm2,植被属长白山植物区系,主要林分类型为天然针阔混交林。土壤类型为山地暗棕色森林土,土层厚20~80 cm。该区属亚温带大陆性季风气候,年均气温3.8 ℃,最热月7月平均气温21.7 ℃,最冷月1月平均气温-18.6 ℃,年均降水量695.9 mm。
2 研究方法
2.1 样地设置与调查 林分类型为原始阔叶红松(Pinuskoraiensis)林经采伐干扰后形成的次生针阔混交林。本研究主要用2009年样地建立时的林木胸径原始数据。样地长660 m,宽320 m,总面积21.12 hm2。调查时将整个样地分为528个20 m×20 m 连续样方,记录每个样方中全部胸径大于1 cm的木本植物的种名,并测量其胸径、树高、冠幅及在样地内的相对坐标。样地共调查到66种木本植物,主要乔木树种的基本数量特征统计结果如表1所示。

表1 样地中主要乔木种的多度、胸高断面积、密度和重要值Tab.1 Abundance, basal area, density, and important value of main arbor species in study plot
2.2 样地异质性生境的划分 在Monte-Carlo模拟过程中,完全空间随机零模型假定研究区域生境均匀同质,要求林木在所有位置出现的概率都相等。当研究区的生境具有明显异质性时,将会极大影响点格局分析。为尽量降低生境异质性的影响,可根据生境属性将样地划分成若干个生境型(Getisetal., 1987; Houetal., 2004)。在计算了本研究样地中每个20 m×20 m样方的海拔、坡度、坡向及凸凹度后(Itohetal., 1995; Harmsetal., 2001),发现样地内树木总胸高断面积与地形因子相关性极高(表2)。因此,在以往研究中(Zhangetal., 2012)利用多元回归树对该样地划分了生境类型。基于此,本研究将整个样地划分成A和B这2个亚区(每个亚区内部生境相对一致)(图1)。

表2 样地中树木总胸高断面积与地形变量之间的相关系数①Tab.2 Correlation coefficients between total basal area and different topographic variables in the forest plot

图1 样地中海拔等高线分布及生境划分示意Fig. 1 Schematic diagram of altitude contour distribution and habitat division in the forest plot
2.3 标记点格局分析 本研究分别在整个样地(不区分生境)、A区和B区3种情境下开展标记点格局分析。标记相关函数Kmm(r)描述了被观测的点格局中标记属性的空间自相关性,已被长期用于研究森林中相邻树木的竞争关系(Penttinenetal., 1992),“标记”可以为胸径、树高或特定时段内的生长特征等指标(Diggle, 2013)。Ripley’sK(r)是一种常见的点格局分析方法,主要依据点的相对位置来分析空间点格局(Ripley, 1981)。标记相关函数Kmm(r)的不同之处在于,其考虑了与Ripley’sK(r)中mi事件相关的定量特征。本研究中采用的定量特征为树木的胸径。标记相关函数Kmm(r)的具体表达公式为(Wälderetal., 1996):
(1)
式中:m为个体的数量;mi、mj表示分别位于i点和j点的2个标记值;wij用于边缘效应的修正;I(i,j)用于指示函数。如果μ是事件mi的平均值,此时观测值与期望值的比值Lmm(r)可计算为:
(2)
当将公式(1)中的mimj替换成(mi-mj)2时,可用来量化树木标记之间的空间关联性。标记点格局的显著性分析基于完全空间随机零模型,通过 Monte-Carlo模拟进行置信度检验,最终计算得出99% 的置信区间。分析过程采用R软件的spatstat包实现。当观测数据计算的标记相关函数Lmm(r)值位于置信区间之上时,表示树木标记间显著正相关; 当标记相关函数Lmm(r)值位于置信区间之下时,表示树木标记间显著负相关; 当观测值位于置信区间之内时表示树木标记间相关性不显著。为了进一步评价不同垂直层树木间的竞争关系,按胸径将林分划分为林下层(5 cm≤DBH<10 cm)、亚林层(10 cm≤DBH<20 cm)和林冠层(DBH≥20 cm)3个垂直层次,并利用标记相关函数分别对3个林层进行分析。
3 结果与分析
3.1 主要树种的标记点格局分析 本研究分析了重要值前14位(表1)的物种在0~15 m尺度上的胸径标记点格局(图2)。结果显示2个亚区中绝大多数树种的个体胸径间呈负相关,尤其是在中小尺度上(A区:r<9 m; B区:r<12 m)未发现个体胸径间呈正相关的树种。在0~15 m尺度上,检验到6个树种个体胸径间呈显著负相关(图3)。色木槭仅在较小尺度上(r<3 m)表现出个体胸径间负相关; 千金榆、裂叶榆和黄檗在r<3 m尺度上个体胸径间相关性不显著,其后随着尺度增大表现出负相关; 白牛槭和胡桃楸则在全部尺度上呈个体胸径间负相关。

图2 不同空间尺度上胸径呈正相关和负相关的树种数对比Fig. 2 Comparison of tree species number with positive and negative correlation in DBH at different spatial scales

图3 胸径呈负相关性的树种标记点格局Fig. 3 Mark point pattern of tree species with negative correlation in DBH
3.2 不同林层个体的标记点格局分析 利用标记相关函数在整个样地上分析林木个体胸径空间关联性。林下层及亚林层的林木胸径间空间相关性不显著。这可能与2个林层中林木个体较小无法通过树冠直接相互作用有关。与之相对应地,林冠层林木在0~6.1 m的尺度上表现出显著的个体胸径间负相关关系(图4)。
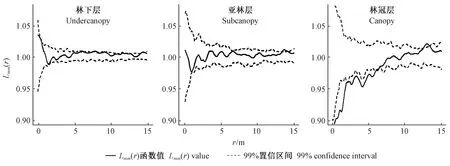
图4 样地中不同林层的标记点格局Fig. 4 Mark point pattern of different forest layers in study plot
为了消除生境异质性带来的潜在影响,在2个亚区中分别检验不同林层个体的胸径空间关联性(图5)。林下层和亚林层林木个体在2个亚区中具有类似的标记点格局,在0~15 m尺度上个体胸径间均没有显著的空间相关性。林冠层林木胸径在A区0~6.9 m尺度和B区0~5.6 m尺度均呈显著的负相关关系。2个亚区不同的作用尺度反映了生境异质性引起了邻体竞争效应上的差异。
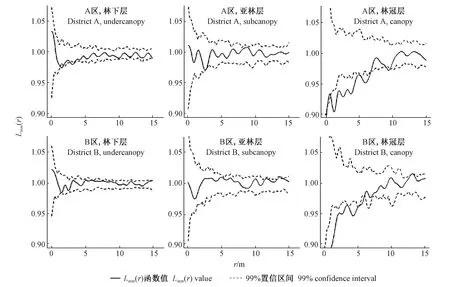
图5 在2个亚区中不同林层林木个体的标记点格局Fig. 5 Mark point pattern of different forest layers of two habitats
4 讨论
4.1 主要树种的标记点格局 森林中的树木空间分布格局是林分的基本特征之一,它由一系列复杂的生态过程综合作用而成,不仅反映了种内与种间关系,同时也反映了物种与生境的关系(梅军林等, 2017; 谭雪莲等, 2019),可以通过空间点格局分析来量化研究(姚杰等, 2018; Ripley, 1981; Diggle, 2013; Hamblyetal., 1996; Illianetal., 2007)。点格局分析的完全空间随机零模型假定研究的环境背景具有同质性(Ripley, 1981)。天然林生境因子多呈异质性分布,而异质性的程度取决于观察尺度(Getisetal., 1987),因此,可以通过生境划分消减生境差异的潜在影响(Houetal., 2004)。
标记相关分析结果显示,绝大多数树种的林木个体胸径在中小尺度上均表现出显著的负相关,这说明胸径较大的树木周围一般生长着胸径较小的同一物种,其主要原因可能是由种子扩散限制和资源竞争所导致(Heetal., 1997; Murrell, 2002; Ertanifardetal., 2017)。大部分种子通过局部扩散的传播方式散落在母株附近,成年树可为周围的幼树提供适宜的生长环境(Getzinetal., 2008; Itohetal., 2010; Canhametal., 1994)。随着树木的生长,其对光照、水分、空间等资源的需求不断增加,个体间的竞争强度也随之增加(Genetetal., 2014)。
通过2个亚区的比较可知,在小尺度上A区林木个体胸径间呈负相关的树种数明显高于B区,这可能与2个亚区的林木密度差异有密切关系。B区海拔高、具有一定坡度、土壤中水分和营养物质流失快。而A区位于低海拔地区,地势平缓、水分充足、光照充足,更适宜林木生长(郝珉辉等, 2017b; Zhangetal., 2012)。A区的林分密度明显高于B区,因此A区的林木对环境资源的需求也加剧,进而导致种内竞争压力增强(李杰, 2018)。Coomes 等(2007)研究发现植物对光照的竞争随海拔升高会减弱。B区中林木个体胸径间表现为负相关的物种比例明显减少,也可能与光照竞争被削弱有关。
4.2 不同林层的标记点格局 在不考虑生境空间异质性的条件下,整个样地中各林层林木个体胸径间的相互作用的分析结果表明,林下层及亚林层林木个体胸径间空间关联性不显著,这与丁胜建(2019)研究发现林下小径级个体需要的资源较少、相邻木间竞争相对较弱的结论一致。此外,林下物种大多具有耐荫特性,可在林冠下很好地存活和生长(臧润国等, 1999)。在0~8 m的尺度范围内,林冠层的树种个体胸径间负相关作用明显,说明林冠层林木对环境资源竞争更强烈。主要原因可能是林冠层多为喜光树种,如红松、水曲柳等,个体间对光照等资源利用的竞争压力随距离减小而增加(李立等, 2010)。通过对不同亚区中各林层林木个体胸径间关系的研究发现,B区林冠层个体作用尺度小于A区,A区个体作用范围为0~6.9 m,B区为0~5.6 m。由此可看出两个生境中的邻木竞争尺度有所差异,这可能是由A、B两区的树木密度差异所导致。森林中树木空间位置分布特征与林木生长之间存在一定关联性(吴初平等, 2018; 郝珉辉等, 2017a; 王宇等, 2020),当林木呈聚集分布时生长特征会表现出明显的负相关作用,负相关强度会随聚集强度的增加而增加(郝珉辉等, 2017a; Fraveretal., 2014)。由A区与B区林木个体作用尺度差异可看出,生境差异性对林冠层林木大小之间的竞争有一定影响,这与Getzin等(2008)认为生境异质性在植物种群结构与空间格局形成中有重要作用的观点相一致。
森林经营是优化林分密度和树种组成,调整林木生长空间的重要举措。如果经营后保留木的间距过小,就会带来生长空间竞争剧烈的风险; 但间距过大,林地的生产潜力又无法充分发挥出来(徐文煦等, 2019)。因此,明确林木竞争范围对森林经营的密度控制具有重要指导意义。本研究仅是对特定森林群落的一个局域性分析,未来有必要对其他森林类型开展进一步研究,以便更深入系统地认识不同群落的竞争效应。
5 结论
复层异龄天然林中生境因子多呈异质性分布,为降低生境异质性对点格局带来的影响,将异质性样地划分为A、B两个亚区(每个亚区内部生境相对同质),利用标记相关函数检验相邻个体间的作用效应。亚林层及林下层个体胸径间空间关系不显著,而林冠层个体胸径在小尺度上具有显著的负相关,A区中林冠层个体间显著负相关的作用尺度为0~6.9 m,B区为0~5.6 m。在不同生境条件下,林冠层内相邻个体胸径的空间负关联尺度是不同的。因此,在森林经营过程中,为确保林木胸径长势良好,确定最终保留密度时要充分考虑生境差异带来的影响。
猜你喜欢
吉林林业科技(2023年6期)2023-11-20 02:13:16
内蒙古林业调查设计(2021年5期)2022-01-05 02:50:40
落叶果树(2021年6期)2021-02-12 01:29:10
水土保持通报(2020年5期)2020-12-23 04:58:48
能源与环境(2020年5期)2020-10-30 00:53:24
安徽医科大学学报(2020年8期)2020-09-09 08:21:20
安徽医科大学学报(2018年9期)2018-09-05 02:55:40
林业勘察设计(2017年4期)2017-07-06 08:46:12
体育科技(2016年2期)2016-02-28 17:06:07
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
