大戟叶绿体基因组特征与系统关系分析
2022-08-19 08:06:38王玉良
安徽科技学院学报 2022年3期
王玉良, 王 松
(安徽科技学院 生命与健康科学学院院,安徽 凤阳 233100)
大戟(Euphorbiapekinensis)是大戟属(Euphorbia)大戟科(Euphorbiaceae)的一种多年生草本植物。大戟属植物遍布全球,数量超过2 000种,是世界性被子植物大属之一,以通体具白色乳汁和独有的杯状聚伞花序为主要识别特征。主要分布于非洲和中、南美洲;约有80种大戟属植物分布于我国的西南和西北[1]。大戟属的很多植物可以入药,大戟即其中之一。大戟也称京大戟、湖北大戟,分布于除新疆、西藏、台湾和云南的全国各地,北方尤为普遍[2]。大戟作为中药材使用已经有2 000余年历史,根含黄酮、萜类等,具有抗炎、抗白血病等作用[3]。大戟市场流通品种比较混乱,作为正品大戟使用的药材来源有大戟和茜草科(Rubiaceae)红大戟(Knoxiavalerianoides),作为非正品使用的有豆科(Leguminosae)的美丽胡枝子(Lespedezaformosa)(草大戟)和瑞香科(Thymelaeaceae)狼毒(Stellerachamaejasme)(绵大戟)[4],各地多以红大戟作为主流品种使用[5-6]。因为大戟属植物生活环境呈现多样性,因而外形特征差异较大。有人提出大戟属应再行划分[7];而有的学者依据分子资料,认为该属应扩大[8];基于传统形态分类学性状,大戟科有狭义的分类系统[9]和广义的分类系统[10]。目前,无论大戟属还是大戟科,其范围的界定仍然难于统一,传统上,大戟科曾被拆分为十几个科[1],而最新资料表明,大戟科可以拆分为4个科,即叶下珠科(Phyllanthaceae),核果木科(Putranjivaceae),狭义的大戟科(Euphorbiaceae)和蚌壳木科(Peraceae)。因此,对野生种群的大戟叶绿体基因组的遗传多样性进行研究,无论对种质资源保护还是物种鉴定,都显得很重要。截至目前,关于大戟叶绿体基因组的研究未见报道。本研究通过对大戟叶绿体基因组的结构进行解析,为进一步研究大戟遗传背景、种质资源保护与开发利用、以及在植物系统发育进化等方面奠定基础。
1 材料与方法
1.1 材料
野生大戟(EuphorbiapekinensisRupr.)幼嫩新鲜叶片采自凤阳县(117°33′35.4″E,32°52′53.6″N),自封袋密封带回实验室,无菌水冲洗若干次,自然晾干后-80 ℃下存放。凭证标本存放于安徽科技学院标本室(AHSTU003289)。
1.2 方法
1.2.1 DNA的提取、纯化和测序 采用CTAB法提取叶片基因组总DNA[10]。DNA质量和完整性采用琼脂糖凝胶电泳检测,浓度和含量利用NanoDrop2000(Thermo Scientific,美国)检测,样品达到测序要求后,送南京集思慧远生物公司(www.jcdetect.cn)构建DNA基因组文库,利用Illumina HiSeq 4000(Illumina, San Diego, CA USA)、采用双端(PE)策略进行高通量测序,读长PE150,共获得原始数据2.28 G。利用NGS QC Toolkit进行质量控制,运行perl程序去除接头和低质量序列,参数默认。
1.2.2 叶绿体基因组的组装与注释 筛选出的clean reads 在GetOrganelle中组装[11],最大组装延伸轮数设置为15,K-mer值和参考数据库默认,组装在Linux系统下运行,挑选最优K-mer值下的结果,用于后续的注释。叶绿体基因组的注释用在线软件GeSeq(Https://chlorobox.mpimp-golm.mpg.de/geseq.html)完成[12],选中tRNAscan-SE和ARAGORN预测tRNA。然后用Geneious R11软件进行手动校正[13]。将注释好的.GB格式文件,使用GB2sequin在线软件(Https://chlorobox.mpimp-golm.mpg.de/GenBank2Sequin.html),转化成tbl和sqn格式的文件,借助在线系统(Https://www.ncbi.nlm.nih.gov/WebSub/)提交到Genbank[14]。叶绿体基因组图利用在线工具OGDRAW绘制[15]。
1.2.3 密码子偏好性分析 相对同义密码子(Relative Synonymous Codon Usage,RSCU)使用情况利用Mega X软件计算和分析。计算公式为:
式中,第i个氨基酸的第j个密码子的出现次数用Xij表示;编码第i个氨基酸的同义密码子的数量用ni表示。当RSCU的值大于1时,表示该密码子的使用有正偏好性(频率较高);RSCU=1表示该密码子的使用无偏好;在RSCU的值小于1的时候,表示该密码子的使用有负偏好性(频率较低)。
1.2.4 系统进化分析 为了确定大戟的系统位置,自NCBI数据库中下载23种大戟科植物、2种黄杨科植物的叶绿体基因组,连同本研究提交的大戟的叶绿体基因组,构建系统发育树,26种植物的名称和NCBI登录号见图3。
使用在线程序MAFFT 7(https://mafft.cbrc.jp/alignment/server/),对26种植物的叶绿体基因组进行多重比对。与大戟科亲缘关系较近的黄杨科植物黄杨木(Buxusmicrophylla)和顶花板凳果(Pachysandraterminalis)设置为为外类群,比对结果导入MEGA X,构建最大似然(Maximum Likelihood,ML)系统发育树(Bootstrap 重复1 000 次)。
2 结果与分析
2.1 叶绿体基因组基本特征
测序后共得到25 944 616条长度约为150 bp 的clean reads,Q20和Q30分别为97.43%和92.64%。利用GetOrganelle进行de novo组装,经注释后得到大戟叶绿体基因组(GenBank登录号:MZ707776)。大戟叶绿体基因组大小162 002 bp,为呈典型四段式结构的双链环状DNA:大单拷贝区LSC、小单拷贝区SSC,以及将二者分开的2个反向重复区域(inverted repeat regions,IRa/IRb),3个区域的大小分别为90 225 bp、18 067 bp和26 857 bp。总嘧啶(GC)和总嘌呤(AT)含量分别为35.7%和64.3%,有明显的AT偏向性(表1)。IR区有4个GC含量较高的rRNA基因,因而该区GC含量最高(56.67%)。这与其他叶绿体基因组的分析结果是一致的[16]。
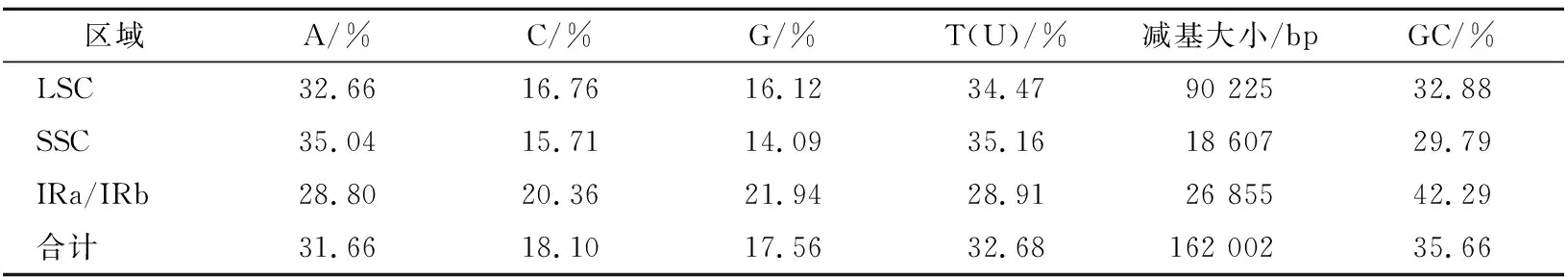
表1 大戟叶绿体基因组的碱基组成
2.2 叶绿体基因组基因分布
大戟叶绿体基因组包括85个蛋白质编码基因(含9个重复基因)、8个rRNA基因(含4个重复基因)和36个tRNA基因(含16个重复基因)。总基因数量129个,非重复基因100个。在反向重复区域(Ira和IRb),有rpl2、rps12和rpl23三个核糖体蛋白大亚基基因、rps7和rps19两个核糖体蛋白小亚基基因、1个NADH脱氢酶基因(ndhB)、ycf1、ycf2和ycf15三个未知功能的基因,以及6个tRNA基因(trnI基因有两个拷贝)和4个rRNA基因。ycf1基因分布在SSC和IRb的交界(图1)。在SSC区,共有10个CDS和1个tRNA基因(trnL)。绝大多数的基因在LSC 区,有57个CDS和17个tRNA基因(表2)。
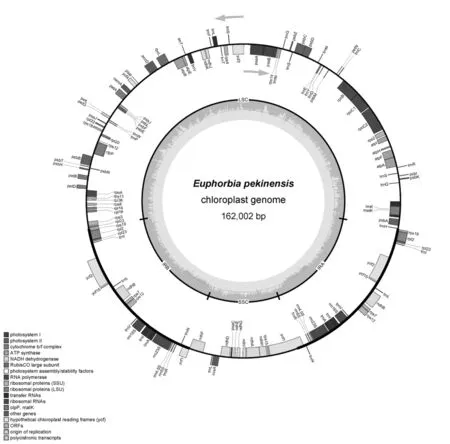
图1 大戟叶绿体基因组基因分布图

表2 大戟叶绿体基因组各区的基因组成
2.3 大戟叶绿体基因组编码基因的分类
在功能上,把大戟叶绿体的全部基因分为4类(表3):自我复制基因、光合作用相关基因、成熟酶基因(matK)、囊膜蛋白(cemA)等其他基因。所有基因中总共有11个内含子,其中6个在IR区,其他均在LSC区(trnL外除)。其中atpF、ndhB、rpoC1、rpl2、trnA、trnI、trnK、trnL和trnV基因各有1个内含子,clpP和ycf3有2个。位于LSC区的trnK-UUU基因序列最长(2 589 bp),matK基因位于其中。
2.4 密码子偏好性分析
有机体基因选择简并密码子有一定偏向性。研究发现,大戟叶绿体全基因组共有64种密码子、编码全部20种氨基酸。密码子数量从230个到2 575个,相对密码子的使用比例从最小的0.43到最大的2.11。Met和Trp只有1个密码子,其余的2~6个密码子(图2)。

表3 大戟叶绿体基因组基因的类别
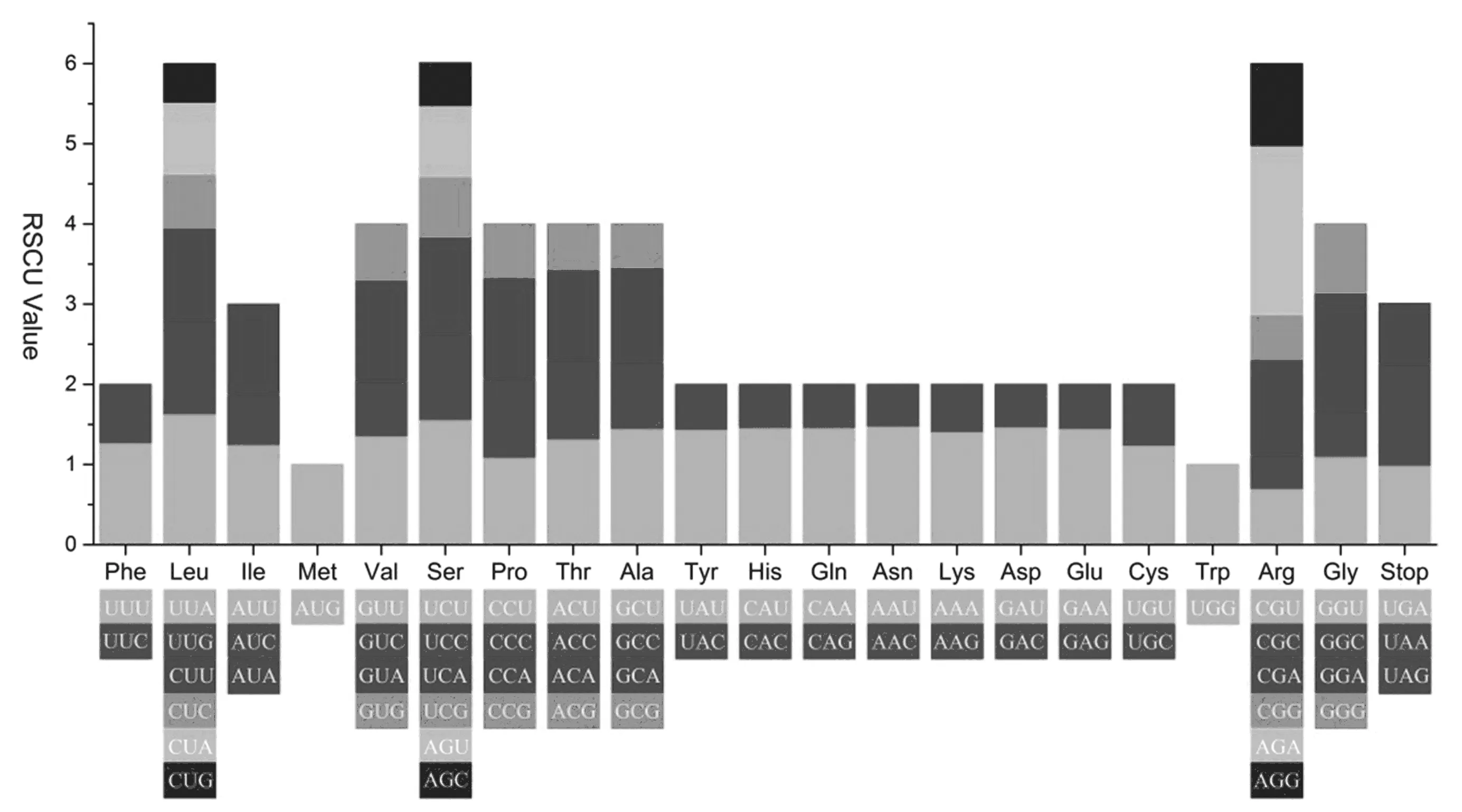
图2 大戟叶绿体基因组20种氨基酸和终止密码子相对同义密码子使用(RSCU)
2.5 大戟的系统发育分析
利用24个大戟科植物叶绿体基因组(23种自NCBI下载)作为内类群、2个黄杨科植物叶绿体基因组为外类群,采用最大似然法构建系统发育树,以确定大戟的系统位置。在ML系统树中,24种大戟科植物大致呈4个分支:白饭树属(Flueggea)、算盘子属(Glochidion)和叶下珠属(Phyllanthus)聚为一个分支;核果木属(Drypetes)的6个种聚为一支;巴豆属(Croton)、麻风树属(Jatropha)、野桐属(Mallotus)和蓖麻属(Ricinus)聚为一支;木薯属(Manihot)、橡胶树属(Hevea)、油桐属(Vernicia)和东京桐属(Deutzianthus)聚为一支;而大戟属的6个种聚为一支。分析结果强烈支持大戟与同属的E.ebracteolata聚为一支,互为姊妹类群。同时,ML系统发育树的拓扑结构中,大戟属的6个种聚为一大支,支持率为100%,与传统分类学的分类结果完全一致(图3)。
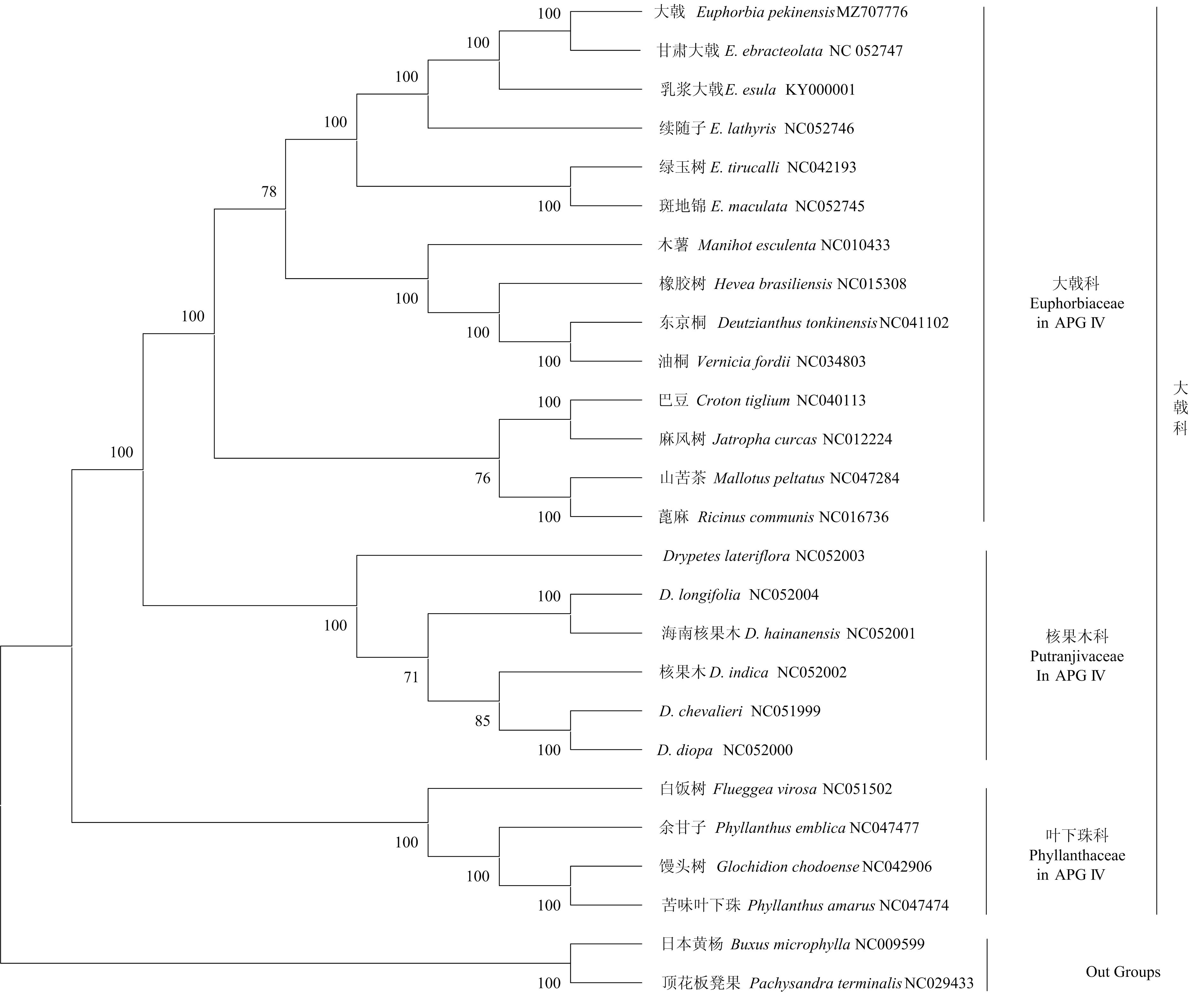
图3 24种大戟科植物叶绿体基因组序列及其外类群构建的系统进化树
3 讨论
中药材的研究具有较大的意义[17-18]。为了保护传统中药大戟的野生种质资源,已有不少学者对大戟进行了各个方面的研究,如生态适宜性[5]、化学成分[19]、药理药性[20]、生药鉴定[21-22]、部分基因的克隆与分析[23]等,而大戟叶绿体全基因组的研究鲜有报道。本文研究结果表明,大戟叶绿体基因组总体结构与其他高等植物相似,例如,全长在160 kbp左右,IR区的嘧啶(GC)含量比LSC区和SSC区的要高,基因组中存在着功能未知的基因等。与大多数被子植物一样,因为IRb/SSC边界的扩张,导致ycf1基因处于IRb和SSC的边界。
除了作为外类群的两种黄杨科植物外,在传统分类学中,本研究涉及的14属、24种植物均属大戟科。ML系统树基本支持传统的分类,即ML系统树支持叶下珠族(Trib. Phyllantheae)、核果木属(Drypetes)和大戟属各自成为一个自然类群。其中本文所涉及的叶下珠族的种类在APG IV中已经划归到新成立的叶下珠科(Phyllanthaceae),核果木属的种类叶已经独立为核果木科(Putranjivaceae)[24]。6个大戟属的种类聚为一支(仍属大戟科),且与APG IV的叶下珠科和核果木科亲缘关系较为疏远(图3)。传统分类结果、ML系统树分析结果和APG IV(2016)分类结果基本一致,在ML系统树中,这些分支都具有较高的支持率,表明传统上利用杯状聚伞花序和通体具有乳汁这两个性状作为大戟属的鉴别性特征,仍具有可操作性。同时,在ML系统树中,叶下珠属的两个种并没有被聚为亲缘关系最近的两个种,而是苦味叶下珠先是与算盘子属的馒头树形成一个亲缘关系最近的分枝,之后该分枝再与叶下珠属的余柑子互为姊妹类群,而且受到了强烈的支持,APG IV对类群的处理没有到种一级水平,因此,形成这种现象的原因值得进一步研究。
猜你喜欢
生物学通报(2020年11期)2020-10-22 01:20:20
中成药(2018年7期)2018-08-04 06:04:10
中成药(2018年7期)2018-08-04 06:04:08
中成药(2018年5期)2018-06-06 03:11:57
中国医药导报(2018年28期)2018-01-18 10:07:26
广西林业科学(2016年1期)2016-03-20 05:33:01
生命科学研究(2014年1期)2014-04-29 00:44:03
中成药(2014年9期)2014-02-28 22:29:01
食品科学(2013年6期)2013-03-11 18:20:13
中国烟草学报(2012年2期)2012-04-09 06:44:56