
IAA与KT对菊芋镉富集和生理响应的影响
2022-08-16 07:05:22郭书亚王燕燕杨颜裴陈虹宇倪晓菁张佩佩吴玉环
水土保持学报 2022年4期
郭书亚,唐 可,王燕燕,杨颜裴,陈虹宇,倪晓菁,张佩佩,吴玉环,刘 鹏
(1.浙江师范大学植物学实验室,浙江 金华 321004;2.杭州师范大学生命与环境科学学院,杭州 310036;3.中国科学院沈阳应用生态研究所,沈阳 110016)
镉(cadmium, Cd)是常见的重金属污染物,随着工业现代化的不断推进,燃煤、冶炼、矿业开采等工程使大量镉进入水体和土壤,造成局部地区镉污染形势严峻。土壤中镉的迁移性、化学活性较强,易被植物吸收并产生毒害,其毒害作用表现在多个方面,如抑制根系生长、破坏叶绿体结构、打破自由基产生与清除的动态平衡、损伤染色体、抑制细胞的有丝分裂等。不同植物的耐镉性存在差异,耐性较强的植株可通过多种机制来抵抗或削弱镉胁迫程度,从而维持正常生长,如提高抗性酶活、螯合镉离子,将细胞内镉离子区域化等。改善土壤镉污染刻不容缓,目前镉污染修复主要有物理、化学、植物、微生物等多种修复技术,植物修复技术近年来获得广泛关注,具有成本低、原位、不破坏土壤结构等优点。因此,耐镉性强、富集能力强的植物在修复重金属污染方面具有较大的潜力。
吲哚乙酸(indole-3-acetic acid, IAA)和激动素(kinetin, KT)是植物激素的典型代表,前者可以调节生长发育和生理代谢过程,后者则在促进细胞扩增、诱导分化及延缓衰老中起着关键作用。外源激素的作用不只是单一调节,而是依赖于相互作用的复杂调控网络。目前,利用植物激素调控作物非生物胁迫下的耐受性已经展开大量研究,在耐盐性、抗旱性、耐重金属等方面均有报道。Bao等研究发现,IAA有效调节了镉胁迫下东南景天(Hance)植株的地上部生物量、叶片叶绿素含量和酶活性,增强植株对镉的吸收和转运;Aldesuquy等研究证明,KT缓解了高粱((L.)Moench)遭受的镉毒,提高叶片中光合色素含量和光合活性,且增加谷物的生物量。可知植物激素在作物生长发育中起着必要的调控作用,可以增强对金属胁迫的耐受性并促进吸收。
菊芋(L.)是菊科(Asteraceae)向日葵属(L.)多年生宿根性草本植物,其生态适应性高,具有较强的耐旱能力。菊芋还具有药用价值,提取出的菊糖可治疗糖尿病;块茎香甜可口,能作为蔬菜食用;其根系具有牢固的抓沙能力,在流动沙丘上长势良好;同时发达的根系可编织成一层防护网络,牢固保持地表层的水土。此外,菊芋也是一种能源植物,地上茎燃点高、热量大,是农村可利用的燃料,其块茎经微生物发酵可转化为乙醇和碳氢燃料,同时,能够作为原料制备氢能及多种化学品。菊芋具有镉积累的潜力,虽然关于菊芋镉富集的研究不少,但通过IAA和KT来增强菊芋对镉胁迫的耐受性及其对镉富集能力的研究还未见相关报道。因此,本项目以耐镉型榆林菊芋和镉敏感型成都菊芋作为试验材料,通过预试验,得到IAA和KT施用浓度,设置不同激素处理组,通过测定菊芋生长、生理生化及荧光特性等指标的变化,并分析不同条件下菊芋镉富集能力,以期探明IAA与KT配合施用促进菊芋吸收镉的作用机理及其内部解毒机制,为多种外源激素协同改良植物修复镉污染土壤技术的应用提供理论依据。
1 材料与方法
1.1 试验材料
本试验于2019年7月1日至2021年5月30日在浙江师范大学化学与生命科学学院植物逆境生物学实验室进行。以前期筛选出耐镉性差异大的2个产地菊芋为试验对比材料,分别对耐镉型榆林菊芋和镉敏感型成都菊芋进行镉胁迫并外源喷施IAA和KT。
1.2 试验设计
将2个产地菊芋块茎用蒸馏水进行冲洗,选择大小一致且芽眼较多的菊芋种于装有石英砂的育苗穴盆中,1孔1块,放置到恒温光照培养箱进行萌发,白天温度22 ℃,夜晚温度18 ℃,光照12 000 lx,湿度调节至50%,每日上午8:00和下午5:00,各组沿着植物根部浇灌1/2 Hoagland营养液,待菊芋幼苗茎长至5~7 cm时,选取长势相近的幼苗移盆至镉含量为300 mg/kg的石英砂中,花盆高为16.5 cm,外口直径25 cm,盆底垫滤纸防石英砂漏出,待菊芋幼苗长至4叶期时,叶片喷施外源IAA、KT,浓度分别对应4,8 mg/L,由二因素四水平正交试验确定激素浓度。共设置5个处理组:CK(空白对照,无Cd)、T1(Cd)、T2(Cd+IAA)、T3(Cd+KT)及T4(Cd+IAA+KT),每盆定植4株幼苗,每处理组设置3个重复,试验重复3次。喷施后第10,20,30天进行相关生理指标的检测,并在终期进行根系活力、切片观察及镉含量的测定,每个指标均重复测定3次。
1.3 项目测定与方法
1.3.1 生长特性测定 采用精度为0.05 cm的直尺测量幼苗株高,用蒸馏水洗去叶、根部残留的沙子并吸干水分,然后用直尺测定主根长度;每株选择多片功能叶,CanoScan4400F扫描仪扫描植物叶片,获得TIF格式的叶面积图,将其导入图形分析软件WinFOLIA中,直接读取总叶面积参数。
1.3.2 光合指标测定
(1)丙酮浸提比色法测定叶绿素含量;(2)PAM-210叶绿素荧光测定仪检测叶绿素荧光参数,晚上7:00后进行试验,黑暗条件下放置20 min,随后测定获得(初始荧光)、(非光化学猝灭系数),(PSII最大光化学量子产量或PSII原初光能转化效率)、(光化学猝灭系数)以及ETR(表观电子传递速率)。
1.3.3 生理指标测定
(1)NBT光化还原法测定超氧化物歧化酶(SOD)活性。各组取叶片0.2 g,放置于预冷的研钵中研磨成匀浆,并加入适当的磷酸缓冲液(PBS),4 ℃下离心20 min以获得酶提取液,按照一定比例和浓度配置反应混合液(甲硫氨酸、核黄素、EDTA-Na、氮蓝四唑),将酶提取液与反应液在温度为25 ℃、光照4 000 lx的培养箱中反应20 min,随后在560 nm波长下测定OD值;愈创木酚法测定过氧化物酶(POD)活性,酶液提取方法同上,取0.2 g愈创木酚,用中性的PBS定容至100 mL,加热搅拌使其溶解即得到反应混合液,470 nm波长下测定OD值;紫外吸收法测定过氧化氢酶(CAT)活性,酶液提取方法同上,取适量30%的过氧化氢,PBS定容至100 mL并摇匀,240 nm测定OD值。SOD、POD、CAT均40 s测1次,测定3次,严格控制反应时间。
(2)硫代巴比妥酸(TBA)法测量MDA含量。各组取叶片0.5 g,放置于预冷的研钵中研磨成匀浆并加入5 mL三氯乙酸(TCA)溶液,质量分数为10%,4 ℃下离心10 min获取上清液,取等体积的0.67% TBA与之反应,在450,532,600 nm波长下测定OD值。
(3)茚三酮显色法测量Pro含量。各组取叶片0.2 g,研磨成匀浆并加入5 mL磺基水杨酸,质量分数为3%,100 ℃水浴加热10 min获取Pro提取液,上清液、水、冰乙酸、2.5%茚三酮按照1∶1∶1∶2进行混合,水浴加热40 min,冷却后加入甲苯2 mL,振荡后静置分层,取上层液体于520 nm波长下测定。
(4)甲烯蓝法检测根系活力,用排水法测定根系体积,约10倍根系体积的0.000 2 mol/L甲烯蓝溶液倒入烧杯中,洗净擦干的根系并浸入烧杯中,放置约1.5 min,随后取出根系并让甲烯蓝溶液回流至烧杯,各组吸取1 mL甲烯蓝溶液且稀释10倍,660 nm下测定吸光度,由标准曲线计算甲烯蓝浓度。

1.3.5 镉含量测定 各组菊芋植株的根、茎、叶分别进行烘干,加入液氮使之研磨充分,各器官取0.2 g样品,用硝酸—氢氟酸进行消解,体积比为5∶1,原子荧光光谱仪测定镉含量。富集系数=植物组织中镉含量/石英砂中镉含量;植物镉提取效率=植物组织中镉的累积量/(石英砂中镉含量×石英砂质量);镉转运系数=叶与茎中的镉含量/根部镉含量。
1.4 数据处理
试验数据采用SPSS 21.0统计软件进行处理,通过单因素方差分析(one-way ANOVA)和Duncan法计算平均值和标准误,分析显著性差异(=0.05及=0.01),利用Origin 8.5软件作图。
2 结果与分析
2.1 外源IAA、KT对镉胁迫下菊芋生长的影响
由表1可知,2个产地菊芋T1组(Cd)的株高伸长率、叶面积、根系伸长率及干重较CK处理均有所下降,榆林菊芋各指标降幅分别为61.75%,9.02%,8.16%,30.14%,成都菊芋降幅分别为30.51%,7.72%,53.38%,32.27%,表明镉处理后2个产地植株的生长均受到抑制,但各指标的降幅存在明显差异。将T2(Cd+IAA)、T3(Cd+KT)处理与T1处理对比得出,IAA与KT单独喷施时可缓解菊芋镉胁迫,2个产地菊芋在T2处理下的4个指标值高于T1处理,榆林菊芋增幅分别为185.60%,5.72%,214.64%,31.83%,成都菊芋增幅分别为48.93%,7.80%,219.82%,21.80%;T3处理各指标值高于T1处理,榆林菊芋4个生长指标的增幅分别为86.62%,3.62%,174.39%,13.12%,成都菊芋分别为188.90%,15.19%,386.77%,43.15%。可以看出,IAA对榆林菊芋的缓解效果较好,而KT则有利于镉胁迫下成都菊芋的生长。IAA和KT同时喷施(T4)处理对镉胁迫下植物的缓解效果最为理想,成都菊芋各指标增幅(与T1处理比较)分别为242.53%,23.87%,898.44%,55.73%,是榆林菊芋的1.21,2.47,2.19,1.19倍,外源激素IAA和KT更加适用于镉敏感型成都菊芋。

表1 不同激素处理对镉胁迫下菊芋生长的影响
2.2 外源IAA、KT对镉胁迫下菊芋光合特性的影响
2.2.1 外源IAA、KT对镉胁迫下菊芋叶绿素含量的影响 叶绿素是植物进行光合作用的重要色素,其含量的高低直接反映植株光合能力的强弱。由图1可知,随着胁迫时间的延长,榆林菊芋T4~T2处理的叶绿素含量先升后降,镉敏感型成都菊芋T3~T2处理呈下降趋势,其T1处理较CK降幅可达16.26%;而耐镉型榆林菊芋能通过内调节提高叶绿素的含量来缓解毒害,其T1处理较CK的增幅可达101.86%。处理30天时,榆林菊芋T2处理的叶绿素含量较CK增加132.52%,成都菊芋T2处理的叶绿素含量较CK增加16.45%;T4处理30天时,2个产地菊芋叶绿素含量较T1处理增幅分别为162.87%,82.36%。同时间下,IAA与KT共同喷施的缓解效果优于单一喷施,榆林菊芋T4处理叶绿素含量较T1处理增长30.22%,成都菊芋T4处理叶绿素含量较T1处理增长114.39%。2个产地菊芋的叶绿素含量均在IAA、KT共同喷施20天时最高。因此,需要合理控制激素的作用时间。

注:图中不同字母表示不同处理间差异显著(p<0.05)。下同。
2.2.2 外源IAA、KT对镉胁迫下菊芋叶绿素荧光特性的影响 单镉处理10天时,2个产地菊芋T1组的(初始荧光)、(非光化学猝灭系数)、ETR(表观电子传递速率)较CK显著升高(<0.05),而(PSII最大光化学量子产量或PSII原初光能转化效率)、(光化学猝灭系数)明显下降(表2)。30天时,榆林菊芋T1处理对应的5种荧光参数较CK处理变化不同,增加10.53%、、、及ETR的下降程度依次为3.14%,15.95%,7.22%,18.47%,表明镉胁迫下植物进行了热耗散,PSII结构受到破坏,电子传递效率降低。外源IAA、KT同时喷施30天的缓解效果优于单独喷施,2个产地菊芋在T4~T2处理所测值均高于T1处理。T4处理条件下,榆林菊芋较T1处理增加46.03%,成都菊芋增加9.52%;榆林T4处理的与ETR较T1处理分别增加6.73%,24.06%,成都菊芋分别增加5.29%,7.92%,且T4处理的2个指标值总体高于T3~T1处理。可以看出,镉破坏菊芋叶片PSII结构,抑制电子传递效率,促进植物的热耗散,外源激素可增强植物的逆境耐受能力,且IAA与KT共同喷施对镉胁迫下榆林菊芋荧光特性的缓解效果优于成都菊芋。

表2 不同激素处理对镉胁迫下菊芋叶绿素荧光特性的影响
2.3 外源IAA、KT对镉胁迫下菊芋Pro含量和MDA含量的影响
植株自身具有一定抗逆性,Pro和MDA含量是衡量其抗镉能力的重要指标之一。由表3可知,激素喷施后镉胁迫下菊芋的Pro有所增加,与T1处理相比,T2、T3处理中成都菊芋Pro含量均在处理30天时的增幅最大,分别为67.21%,90.08%,T4处理中,榆林菊芋在20天时增幅可达223.94%,成都菊芋为112.73%。镉胁迫30天时,榆林菊芋T4~T2处理的Pro含量较T1处理的增长率分别为106.47%,70.82%,57.31%,成都菊芋分别为192.38%,90.08%,67.21%,后者的增长幅度明显更大。此外,单镉胁迫下MDA含量始终维持在较高水平,激素喷施后榆林菊芋、成都菊芋的降幅分别可达39.13%,38.35%,且在T4条件下处理30天时出现。MDA含量的变化趋势与Pro相反,镉处理30天后,榆林菊芋T4~T2处理降幅分别为39.13%,26.09%和28.26%,成都菊芋降幅分别为38.39%,33.93%,33.93%。综上,适当IAA和KT处理能通过增加植株体内Pro含量并减少MDA含量缓解镉毒,以二者共同喷施使菊芋获得更强的抗逆性。
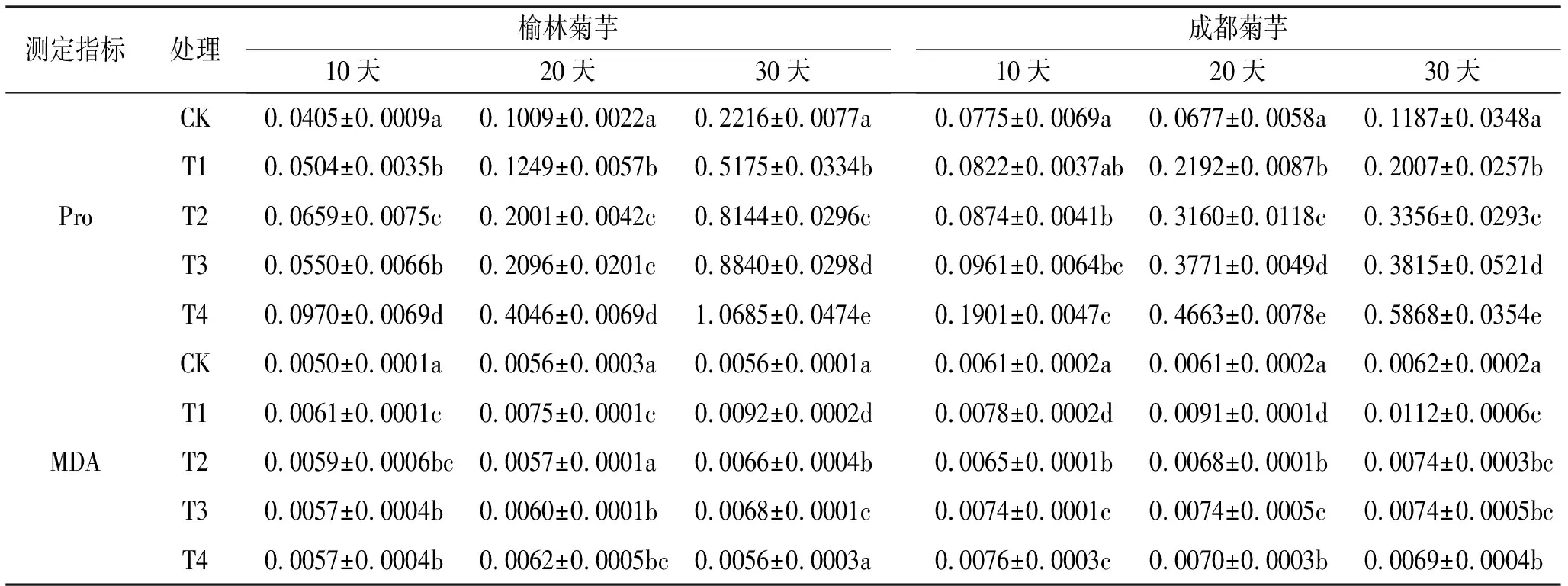
表3 不同激素处理对镉胁迫下菊芋Pro和MDA含量的影响 单位:μmol/g
2.4 外源IAA、KT对镉胁迫下菊芋抗氧化酶的影响
抗氧化酶系统能有效抵抗胁迫下由活性氧自由基所造成的伤害,与植物抗逆性密切相关。由图2~图4可以看出,激素喷施后2个产地菊芋的抗氧化酶活性出现不同程度的变化,这与菊芋对镉的耐受性有关。处理30天后,榆林菊芋T3、T2处理SOD活性的变化趋势相同,较T1处理分别下降11.15%,11.04%,POD活性则分别增长42.54%,83.57%,CAT分别上升0.87%和1.65%;成都菊芋T3、T2处理的SOD活性分别提升13.81%,17.47%,POD活性分别增加30.99%,13.87%,CAT活性的增幅分别为12.05%,14.98%。受镉毒害30天后,T3、T2处理中只有榆林菊芋的SOD活性降低,2个产地植株的CAT、POD活性均增加。榆林菊芋POD、CAT活性在T4处理下的增幅分别可达315.12%和400.72%,成都菊芋则分别为16.24%和368.91%。
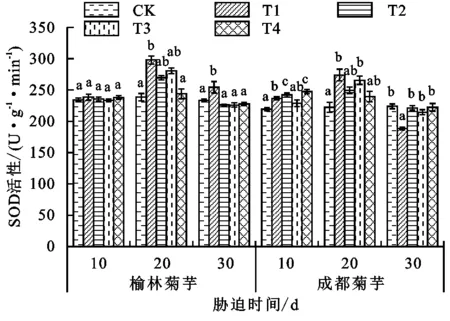
图2 各处理下菊芋SOD活性
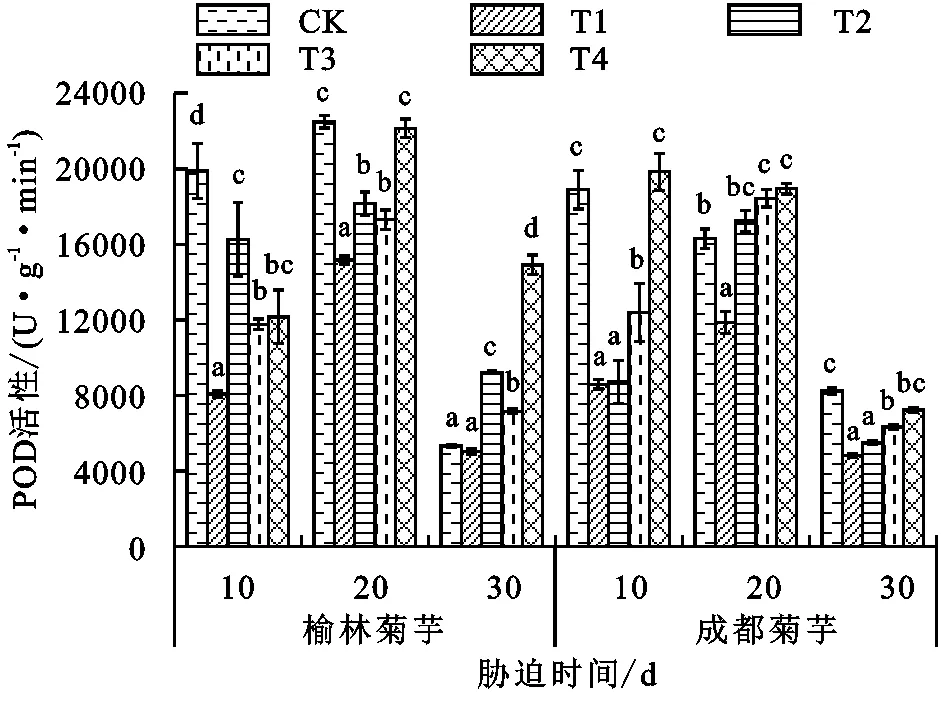
图3 各处理下菊芋POD活性
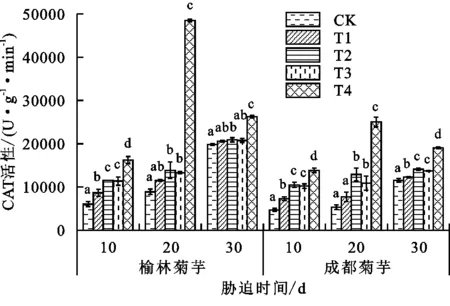
图4 各处理下菊芋CAT活性
由此可得,IAA和KT共同喷施能提升菊芋清除自由基的能力,且不同产地菊芋在耐受性方面存在差异。
2.5 外源IAA、KT对镉胁迫下菊芋根系活力的影响
植物根系是活跃的吸收合成器官,最先接受外界胁迫信号,根系活力能够直接反映作物的生长情况和耐镉能力。由图5可知,T1处理下,榆林菊芋的根系活力较CK处理增长20.96%,成都菊芋的根系活力较CK处理下降16.06%;在T3、T2处理下榆林菊芋的根系活力最大增幅为31.14%,成都菊芋的根系活力最大降幅为11.18%,成都菊芋受镉毒影响根系活力下降,单一激素喷施均能缓解根系损伤,且增强镉胁迫下榆林菊芋的根系活力;激素共同喷施后,2个产地菊芋根系活力分别上涨39.00%,9.10%。各处理组中榆林菊芋的根系活力始终高于成都菊芋,单镉胁迫下榆林菊芋的根系活力上升,成都菊芋的根系活力下降,IAA对植株镉毒的缓解效果优于KT。可见,IAA和KT共同喷施能有效缓解镉对成都菊芋根系的毒害,且增强耐镉型榆林菊芋的根系活力。

图5 各处理下菊芋根系活力的变化
2.6 外源IAA、KT对镉胁迫下菊芋茎横切片的影响
茎是根和叶之间起输导和支持作用的重要营养器官,在逆境条件下根部镉含量增加使根系结构受损,造成堵塞,阻碍水分运输,因此,在显微镜下能观察到茎横切片髓部形成气泡。从图6可以看出,单一喷施外源激素后,榆林菊芋、成都菊芋黑色区域增幅可分别达85.19%,343.75%;T4处理下,2个产地植株黑色区域较T1处理分别扩增142.59%,700.00%,成都菊芋的扩增幅度远大于榆林菊芋。由此得出,IAA和KT共同喷施后茎部水分运输受阻愈发严重,推测激素促进菊芋根部对镉的吸收,引起根系损伤,进而影响根系从环境中获取水分。

图6 不同激素处理对镉胁迫下榆林菊芋、成都菊芋茎横切片的影响
2.7 外源IAA、KT对镉胁迫下菊芋镉富集效能的影响
富集系数和转运系数表示镉的累积和转运,镉提取效率则直观表明植株的镉富集能力。由表4可知,菊芋各器官镉含量的大小依次为根>茎>叶。T4处理中,榆林菊芋镉含量较T1处理增幅为21.05%~32.23%,成都菊芋镉含量较T1处理增幅为20.29%~27.19%,均大于T2、T3处理。可见,外源激素增强菊芋的镉富集能力,IAA与KT共同喷施后榆林菊芋的吸镉能力明显升高。T4处理下,2个产地菊芋的镉富集系数分别为5.24~7.51,4.10~6.83,转运系数均大于1。2个产地菊芋的镉提取效率均在T4处理出现最大值,与T1~T3处理相比达到显著水平(<0.05),分别为6.05%,5.02%,榆林菊芋T4处理的镉提取效率分别是T3~T1处理的2.52,1.70,3.29倍,成都菊芋T4处理的镉提取效率分别是T3~T1处理的2.52,1.79,3.72倍。由此可知,IAA和KT共同喷施可增强菊芋镉富集、转运能力及镉提取效率,且2种激素对2个产地菊芋的增强效果相近。

表4 不同激素处理对镉胁迫下菊芋镉富集效能的影响
3 讨 论
3.1 外源IAA、KT对镉胁迫下菊芋生长特性及光合特性的影响
镉对植物的毒害作用直观表现为阻抑生长发育,同时破坏叶绿体膜,改变叶绿素含量和荧光参数,限制光合作用的进行。Alyemeni等研究发现,镉对植株的生长具有负面影响,同时镉积累导致叶绿体结构受损,从源头遏制光合作用,阻碍有机物的合成,而外源激素能有效减轻毒害效果,本次试验也得出相似结果。T1处理下菊芋的生长受到抑制,表现为株高伸长率、叶面积、根系伸长率及干重低于CK处理,叶片叶绿素含量降低,施用外源激素后则明显升高,且在共同喷施时达到最大。IAA是植物体内的一种内源生长素,合成前体为色氨酸或非色氨酸物质,主要通过吲哚乙醛氧化产生,在植物组织中常以结合态形式存在,能够促进细胞伸长,其生理作用具有两重性,与其浓度大小有关;KT是细胞分裂素的一种,具有促进细胞分裂、增加气孔开度、延缓植物衰老的作用,二者具有高度的生理活性。植物可从环境中吸收外源激素,以调控自身生长及提高逆境耐受能力。单镉胁迫下、显著上升,、以及ETR呈降低趋势,说明菊芋以增加热耗散的方式释放过剩的激发能,同时非生物胁迫抑制光化学能力、PSⅡ反应中心的开放程度降低和电子传递链受阻;外源激素处理后,与下调,、以及ETR升高,并在激素共同喷施下达到最高,进一步表明IAA和KT可缓解镉胁迫,在一定程度上提升光合速率。荧光参数可用来分析光合速率在逆境下降低的原因,也能反映关于光合机构的信息,张婷婷等研究不同浓度IAA对镉胁迫下栝楼(Maxim.)荧光参数的影响发现,35 μmol/L的IAA能有效缓解镉胁迫,提高光合电子传递效率及PSII反应中心的开放程度,这与本研究结果相同。综上所述,施加一定浓度的IAA和KT有利于削弱镉对叶片叶绿素浓度以及光系统的破坏和抑制程度,通过修复光反应中心活性,维持光合作用的正常进行,使菊芋长势保持良好,但IAA与KT的交互作用复杂,阐明镉胁迫下2种激素间的作用机制还有待研究。
3.2 外源IAA、KT对镉胁迫下菊芋抗氧化系统及渗透调节的影响
植物在逆境条件下,活性氧(ROS)产生与清除的动态平衡被打破,导致细胞内的过氧化氢、羟自由基、超氧阴离子等ROS大量爆发并积累,为了减少其对细胞结构造成的氧化损伤,植物体内演化出了一套抗氧化酶系统以清除过量的氧离子、过氧化物及含氧自由基。植物在逆境下可增强抗氧化酶基因的表达,王艳等研究发现,水稻能够提高保护性酶活的表达水平以应对高温胁迫,该响应机制与本研究相似,即镉胁迫下SOD与CAT活性均有不同程度的提高,2种酶扮演的角色不同,SOD负责歧化超氧阴离子,生成过氧化氢,CAT则将过氧化氢催化分解为水和氧气,从而清除ROS。本试验中,镉抑制菊芋的抗氧化酶活性,2种激素共同喷施后3种保护酶活性的增幅优于单独喷施,且2个产地菊芋变化幅度也存在明显差异,这与植株对非生物胁迫的耐受性有关。此外,在镉胁迫环境下,IAA与KT共同处理可激活SOD、POD和CAT活性,并使其生长状况更快接近CK水平,这与吴东墨等在蜈蚣草((Linn.)Merr.)上的试验结果相一致。菊芋叶片游离Pro和MDA含量的变化情况再次验证了上述结论。已有研究发现,Pro在植物细胞体内具有多种功能,可通过清除活性氧、保护抗氧化酶系统、维持水分平衡等多种方式缓解镉毒,其含量积累有利于提高植株的非生物胁迫耐受性;MDA是膜脂过氧化的产物之一,它的产生能进一步加剧膜损伤。本试验证明,菊芋在遭受镉胁迫时,体内Pro和MDA含量均大幅提升,在加入外源激素后MDA下降,而Pro则仍呈上升趋势,2个指标在激素共同喷施下均达到最佳,且其效果随着周期延长愈发明显。综上可知,IAA和KT可通过维持相对较高的Pro含量并启动抗氧化系统以减弱细胞脂膜过氧化损伤程度来保护细胞完整,维持细胞正常生理活动,提高作物耐镉性。
3.3 外源IAA、KT对镉胁迫下菊芋根尖损伤及镉富集、转运的影响
根系活力与植株生长密切相关,是反映根部活性和植株耐受性的重要参考指标。李萍等研究发现,喷施KT能增强玉米幼苗的根系活力和生物量,提升其对土壤中镉的提取效率。本次试验也呈现出相似结果,IAA和KT使镉胁迫下菊芋的根系活力上升,较单一施用效果更佳。茎具有输导营养物质和水分的功能,根系直接与外界环境接触,其结构受损将导致茎部缺水,从而影响植物的正常生长。Shen等研究指出,植物在非生物胁迫下导管发生栓塞,形成大量气泡,此时植物处于缺水状态,重新湿润切片可使气泡先伸展,然后塌陷消失,从而修复导管栓塞。本研究得出,4 mg/L IAA、8 mg/L KT共同喷施使得菊芋茎横切片的气泡数目明显增多,这说明植物激素共同喷施不能修复镉毒对菊芋茎部造成的栓塞,但能改善根系统,以增强吸收重金属的能力,从而帮助菊芋有效应对镉胁迫环境并强化其富集能力,这很可能是超富集植物在重金属污染地区生存的内部保护机制。
已有研究发现,苎麻((L.)Gaudich.)、狼把草(L.)、杨树(spp)等植物耐镉性强,且它们的镉吸收能力存在差异,这与其自身的遗传特性有关。根部较其他器官往往吸收更多重金属,菊芋具有根系发达、生物量大、易种植等特点,因此,选择2个耐镉差异大的植株进行对比试验,结果得出,2个产地菊芋各组的镉含量情况都表现为根>茎>叶,根部直接吸收营养物质和水分,其镉含量最高,茎次之。榆林菊芋各器官的镉含量、富集系数、转运系数和提取效率均高于成都菊芋,这也进一步证明该地区植株的镉富集能力和耐镉性强,且外源IAA、KT喷施后榆林菊芋植株根、茎、叶的镉含量、富集系数等均达到最大值,这与Liang等探索得出IAA和草酸(OA)联合施用有效增强植物对镉、铅吸收的结论相似,该文也指出,植物激素共同处理效果大于单一喷施。植物能调节自身的内源激素水平以缓解重金属胁迫,外源喷施激素同样可提高植物的逆境耐受性,IAA和KT通过保护菊芋根系,提高抗氧化能力,减少MDA含量,促进Pro生成及光合系统修复等途径,以维持菊芋在镉环境中生长并达到吸附目的,但IAA、KT单独及共同作用下提高菊芋耐受镉毒的相关机制及其对植物修复效率的影响还需要更为深入的研究。菊芋具有食用、药用和饲用价值,是一种人畜共用作物。此外,菊芋还是一种能源植物,从中提取出的菊糖加工后发酵可产生酒精,同时地上茎晒干后燃点高,是农村地区很好的燃料。综上可知,在镉污染地区种植能源植物菊芋,同时施用IAA、KT(浓度对应4,8 mg/L)以促进菊芋的镉富集能力,这不仅改善重金属污染环境,而且还兼具经济效益。
4 结 论
(1)300 mg/kg镉胁迫下,2个产地菊芋生长特性、光合特性均受到不同程度的抑制,株高伸长率、叶面积、根系伸长率及干重显著降低(<0.05)。4 mg/L IAA、8 mg/L KT最适配比喷施可有效缓解植株受到的镉胁迫,生长30天后,成都菊芋各指标普遍高于榆林菊芋;榆林菊芋植株与ETR增幅分别是成都菊芋的1.29,3.04倍;榆林菊芋和成都菊芋的叶绿素增长率分别为30.22%,114.39%,从生长情况及叶绿素含量变化来看,IAA与KT对成都菊芋的缓解效果优于榆林菊芋。
(2)抗氧化酶系统与植物的抗逆性密切相关,可清除ROS以减少其对细胞造成的氧化损伤,Pro同样具有此功能。镉胁迫30天后,榆林菊芋、成都菊芋T2、T3处理的CAT、POD活性均增加,T4处理下,2个指标均可达最大值,且2个产地菊芋相差298.88%,31.81%。Pro在T4处理达最大增幅;而MDA在T4处理的降幅大于T2、T3处理,说明IAA和KT可通过维持相对较高的Pro含量,并启动抗氧化系统以减弱细胞脂膜过氧化损伤程度来保护细胞完整,维持细胞正常生理活动,提高作物耐镉性。
(3)镉胁迫造成菊芋根系损伤,阻碍水分运输,2个产地菊芋T4处理的根系活力分别提高39.00%,9.10%,增幅大于T2、T3处理;镉含量均表现为根>茎>叶,IAA与KT单独使用均可促进菊芋对镉的吸收,但两者共同喷施的效果更佳,且T4处理的富集系数、转运系数及提取效率均高于其他处理,进一步表明外源激素能够通过增强根系活力等途径,帮助菊芋有效应对逆境并强化其富集和转运能力。
猜你喜欢
公民与法治(2022年4期)2022-08-03 08:20:56
舰船科学技术(2022年11期)2022-07-15 07:51:56
音乐天地(音乐创作版)(2021年7期)2021-10-13 08:10:42
人大建设(2020年1期)2020-11-17 05:49:22
西藏农业科技(2019年3期)2019-11-04 00:35:10
今日农业(2019年16期)2019-09-10 08:01:36
现代园艺(2018年3期)2018-02-10 05:18:12
——为榆林抗洪救灾而作
音乐天地(音乐创作版)(2017年6期)2017-10-14 11:12:01
上海农业学报(2017年3期)2017-04-10 12:39:12
湖南农业(2016年11期)2016-03-07 15:55:35
