
小熊猫犬瘟热病毒分离鉴定及其H基因序列分析
2022-08-11 02:35:26陈诗涵黄紫贝管飘萍王海燕金文杰刘文博
野生动物学报 2022年3期
陈诗涵 黄紫贝 管飘萍 郭 鑫 王海燕 金文杰 刘文博,4*
(1.扬州大学兽医学院传染病学教研室,扬州,225009;2.江苏省动物重要疫病与人兽共患病防控协同创新中心,扬州,225009;3.江苏农牧科技职业学院,泰州,225300;4.扬州大学动物疾病检测与技术服务中心,扬州,225009)
犬瘟热病毒(Canine distemper virus,CDV)是一种高度传染性病毒,可在家养和野生犬科(Canidae)、熊科(Ursidae)及非人灵长类动物等多种动物中引起多系统、亚临床甚至致命的疾病[1-4]。近年来,由于野生动物园的快速发展,犬瘟热病毒的自然宿主暴露和接触频率有所提高,对野生动物的健康造成了巨大的威胁。犬瘟热病无季节性,不同年龄和性别均可感染发病,幼龄动物死亡率高达80%以上,危害严重[5-8]。据研究,CDV编码的H蛋白的糖基化与其对宿主的致病性有关[9],且H蛋白较易发生变异,因此常以血凝素H基因的变化来评估CDV毒株间的遗传变异[10]。目前根据H基因编码的氨基酸序列,CDV毒株已分为18个主要遗传谱系,主要包括America Ⅰ、America Ⅱ、Asia-1、Asia-2、Asia-3、Europe、European wildlife、Arctic、South America和Southern Africa等,之后又增加了Asia-4型[11]。本研究对江苏某动物园小熊猫(Ailurusfulgens)犬瘟热病诊断和病原学研究,通过对H基因测序,并对其序列进行同源性比较和遗传发生分析,鉴定2株CDV病毒分别为Africa毒株和European wildlife毒株,以期为野生动物犬瘟热病的防控提供参考。
1 基本情况
2019年10月21日,江苏某动物园1只小熊猫突然死亡,由于事发突然,未检测。10月31日,又有2只小熊猫发病,用犬瘟热胶体金试纸条检测,结果为弱阳性,对2只小熊猫进行血常规检查,其中一只小熊猫红细胞略升高,其他无异常,另一只小熊猫嗜中性粒细胞和单核细胞显著升高。
11月19日起,对患病小熊猫皮下注射头孢拉定。11月31日起,对全体小熊猫进行干扰素和犬瘟热单抗治疗,连续4 d,并继续注射头孢类药物,但呼吸道症状未见明显改善。改用康卫宁给药,2~3 d后未见效果,加用阿奇霉素1 d,恩诺沙星3~4 d,仍未见效果,改用美罗培南持续给药。12月10日起,用抗CDV高免血清治疗,连续4 d,期间小熊猫进食少时,给与葡萄糖氯化钠注射液补液治疗,随后病情得到控制。
2 疾病诊断方法
2.1 病理剖检及采血
对病死小熊猫剖检。无菌采集2管小熊猫的尾静脉血,用PBS以1∶3倍稀释,5 000 r/min离心5 min,取上清,用0.22 μm的滤器过滤,-80 ℃保存备用。
2.2 PCR检测
取稀释后的样品100 μL,根据RNA simple提取试剂盒(天根生化科技(北京)有限公司)说明书提取RNA。根据TaKaRa RNA PCR Kit (AMV) Ver.3.0试剂盒(宝日医生物技术(北京)有限公司)说明书,用提取的RNA反转录合成cDNA。RT-PCR反应体系为:MgCl22.00 μL,10×RT Buffer 1.00 μL,RNase Free dH2O 3.75 μL,RNase Inhibitor 0.25 μL,AMV Reverse Transcriptase XL 0.50 μL,Random 9 mers 0.50 μL,Sample 1.00 μL。反转录反应条件为:30 ℃保温10 min,使Random 9 mers延伸达到足够长度,50 ℃退火,与模板RNA充分结合30 min,95 ℃加热5 min,使AMV RTase失活,5 ℃冷却5 min。参考相关文献[12-13],合成鉴定犬瘟热病毒的引物CDV-F和CDV-R(表1),预期扩增的目的片段长度为455 bp。

表1 CDV鉴定及H基因扩增的引物
PCR反应体系:cDNA 10.00 μL,5×PCR Buffer 10.00 μL,TaKaRa E×TaqHS 0.25 μL,CDV-F 0.50 μL,CDV-R 0.50 μL,ddH2O 28.75 μL。PCR反应条件:94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1 min,共30个循环;72 ℃延伸10 min。PCR产物经1%琼脂糖凝胶电泳得到符合要求的PCR产物送往南京擎科生物科技有限公司测序。
2.3 病毒分离培养
取出Vero-CD150细胞(此前在本实验室-80 ℃保存)复苏,待培养长至80%后弃去培养基,用PBS清洗1或2次。取过滤除菌的小熊猫CDV阳性血清样本100 μL,按1∶10的比例加入无血清DMEM(Gibco公司),混匀后接种单层Vero细胞,细胞培养箱中37 ℃、5%CO2吸附2 h,弃去培养基,补加含1%G418(赛默飞世尔科技公司)的抗生素和2%胎牛血清(FBS)细胞培养基(Gibco公司),同时将未接毒的正常Vero-CD150细胞作为阴性对照,于细胞培养箱中37 ℃、5%CO2培养,每日观察细胞病变情况。当Vero-CD150细胞有80%及以上出现细胞病变时进行收毒,反复冻融3次,传至第3代。所有细胞培养物置于-80 ℃冻存备用。
2.4 H基因扩增
合成3对针对犬瘟热病毒H基因的特异性引物:CDV4F/CDV4R、CDV5F/CDV5R和CDV6F/CDV6R(表1),预期扩增的目的片段长度分别为471、571、1 014 bp。以上引物均由生工生物工程(上海)股份有限公司合成。扩增细胞培养物H基因,PCR反应条件参考2.2。
2.5 同源性和遗传进化分析
在GenBank中下载不同基因型(Asia-1、Asia-2、America-1、America-2、Africa、Arctic like、Europe和European wildlife)的CDV参考毒株(35株),应用DNASTAR软件中的MegAlign分析核苷酸序列,并比对这35株参考毒株和2株分离株的基因序列。使用BioEdit进行Clustal同源性比较,并用MEGA-X软件的Neighbor-joining(邻位相连法)绘制遗传进化树,Bootstrap设为1 000。
3 结果与分析
3.1 病理剖检
剖检发现,病死小熊猫胸腔内肺部充血并伴有斑块状出血(图1A),肝脏肿大,心内膜出血(图1B),胃肠黏膜肿胀、出血(图1C)。

图1 病死小熊猫出血病变
3.2 PCR扩增
PCR扩增后得到长度约455 bp的目的条带(图2)。PCR产物测序结果经BLAST分析,2份血样均为CDV阳性,测序确定为CDV基因片段。

图2 小熊猫血样PCR扩增结果
3.3 CDV分离培养
接种病毒的Vero-CD150细胞,24 h后均出现圆缩、细胞颗粒增多、融合和聚集成多核巨细胞等细胞病变效应(图3A),且部分细胞脱落、坏死。连续传代后细胞病变效应(cytopathic effect,CPE)稳定产生,2个毒株分别命名为SZ2019CDV1H和SZ2019CDV2H。而未接种病毒的Vero-CD150细胞则无变化(图3B)。
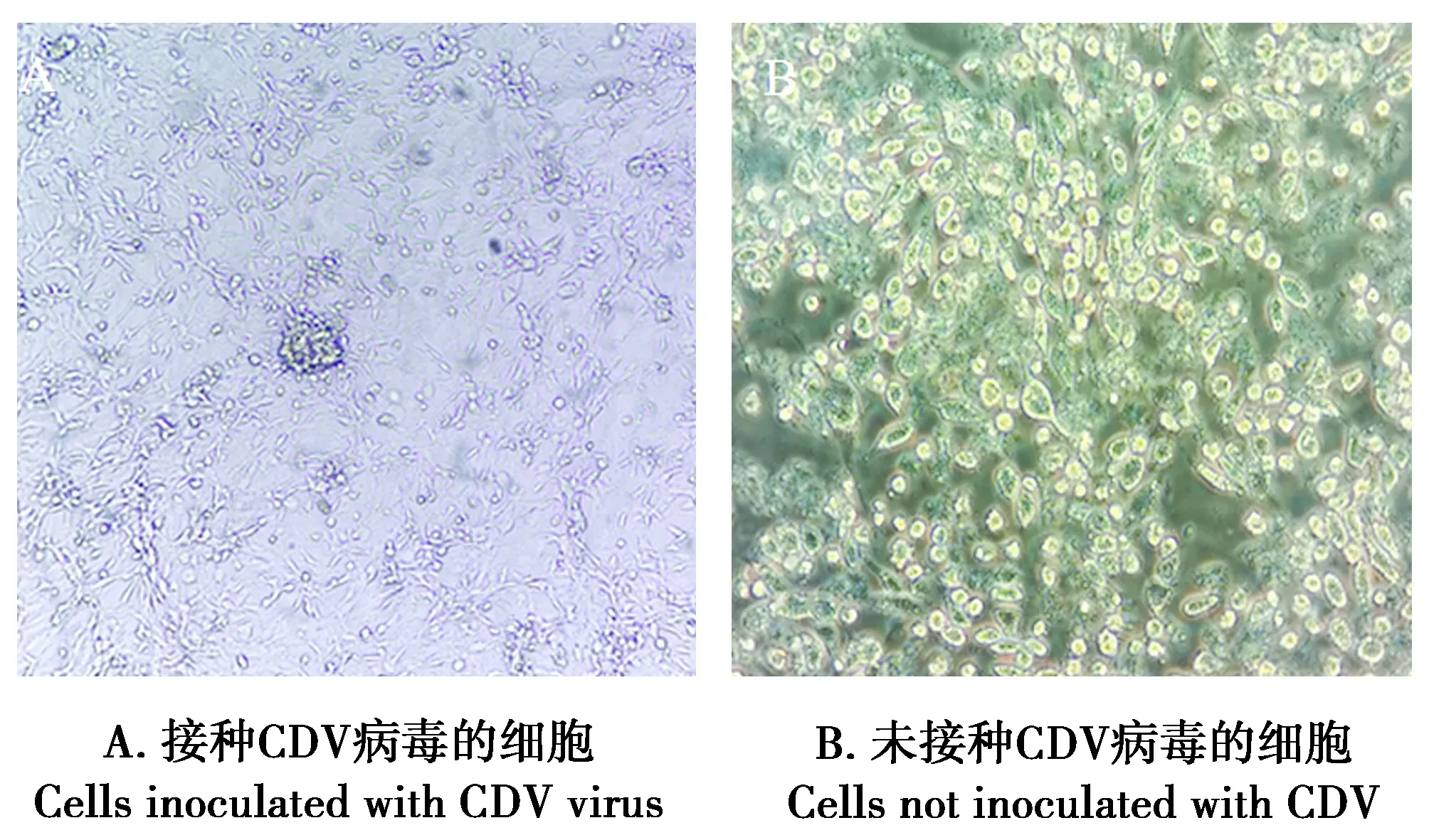
图3 分离株的犬瘟热病变(100×)
3.4 H基因的扩增
用犬瘟热病毒H基因的特异性引物对细胞培养物进行H基因扩增,扩增出H基因的3段序列,电泳得到预期长度的目的条带(图4),测序后拼接得到完整的H基因序列,总长度为1 824 bp。
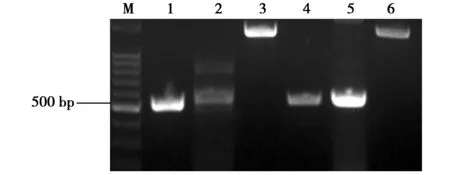
图4 细胞培养物中H基因3个片段的PCR产物
3.5 同源性分析
利用MegAlign软件对已发表在NCBI上的其他35株CDV的H基因序列与本试验测得的2株CDV基因序列进行比对。SZ2019CDV1H株与SZ2019CDV2H株的基因同源性为97.2%,两毒株与标准European wildlife毒株Rockborn-Candur (GU266280.1)和Rockborn (GU810819.1)的同源性较高,全基因同源性达96.7%,但与America毒株同源性相对较低,特别是America-1疫苗株Snyder Hill (AF259552.1),同源性分别仅为91.3%和91.0%(图5)。

图5 不同CDV毒株基因同源性
3.6 遗传进化树分析
由图6可以看出,小熊猫源SZ2019CDV1H、SZ2019CDV2H株与其他35株代表毒株可分为8个遗传系。从遗传进化树分析,SZ2019CDV1H株与Africa毒株亲缘关系较近,SZ2019CDV2H株与European wildlife毒株亲缘关系较近,两毒株均不与Convac、Vacc-Q和Vaccine等America-1疫苗毒株在同1个分支上,遗传进化距离相距甚远。

图6 不同CDV毒株H基因核苷酸序列系统进化树
4 讨论
根据小熊猫的临床发病情况首先怀疑为犬瘟热病毒感染,将临床送检的血样进行分子生物学检测,在获得阳性结果后分离和鉴定病毒,并对分离得到的病毒H基因扩增和测序,以确定其遗传谱系。近年来,国内报道的引起犬[14-15]、东北虎(Pantheratigrisaltaica)[16]、水貂[17]和小熊猫[18-19]等野生动物高病死率的主要流行犬瘟热遗传系为Asia-1。经同源性分析,本研究获得的CDV分离株与Europe毒株同源性较高;但在遗传进化分析上,SZ2019CDV1H株与Africa毒株同处1个分支,SZ2019CDV2H株与European wildlife毒株同处1个分支,与常见Asia-1差距较大,与America-1疫苗株遗传进化距离最远。这说明CDV的遗传演变存在不同的方向性,这对基因分型和演化关系分析可能会存在一定的干扰。
因目前还没有小熊猫源性的犬瘟热疫苗,我国预防此病多用犬用弱毒苗,该苗对野生动物并不一定适用。在高强度的疫苗免疫压力下,流行毒株发生变异,从而造成了市面上商品化的犬瘟热弱毒疫苗对犬以外的物种免疫效果不佳[18]。
基因分型往往基于病毒所分离的地域[19],而遗传关系代表了毒株本身的变异程度。本研究在一次疫病暴发事件中分离得到了2种不同的基因型病毒,可以推断病毒来源相对比较复杂,可能与动物的来源及病原体在动物园内的散播有一定的联系。由于数据和信息的缺乏,还不能确定此次疫病暴发的原因,但是这些复杂的病原学特征为临床发病后的治疗造成很大的困难。本研究中的数据和信息将为犬瘟热病毒地方性流行毒株的分子流行病学调查提供基础数据,流行毒株遗传系分析结果可为临床筛选疫苗和治疗提供一定参考。
猜你喜欢
科学大观园(2022年2期)2022-01-23 11:05:15
现代畜牧科技(2021年7期)2021-07-28 06:40:42
疯狂英语·新读写(2020年1期)2020-04-20 13:30:10
学生天地(2018年36期)2019-01-28 08:13:44
小哥白尼(野生动物)(2018年11期)2018-12-18 02:01:34
学生天地·小学低年级版(2018年12期)2018-01-15 11:33:56
动物医学进展(2015年10期)2015-12-07 05:46:18
特产研究(2014年4期)2014-04-10 12:54:12
郑州大学学报(理学版)(2014年3期)2014-03-01 04:21:08
养殖与饲料(2014年10期)2014-02-28 22:14:59
