
扁蓄二型性种子休眠及萌发特性
2022-08-10 06:27:46秦启娟严佳玥
草业科学 2022年7期
秦启娟,严佳玥,魏 岩
(新疆农业大学草业学院 / 新疆草地资源与生态重点实验室, 新疆 乌鲁木齐 830052)
种子萌发是植物生活史的开端,在植物种群的持续和扩散中起着重要的作用。种子萌发阶段对环境的抵抗力最弱,容易受外界因素的影响。对于生活在极端环境中的植物来说,种子在任何适宜的条件下萌发都存在一定风险。种子休眠能够躲避恶劣环境,阻止种子在不适宜的时期萌发,减少子代的风险,确保幼苗存活,这是植物适应恶劣的生存环境和保障种群延续的一种有效策略。种子异型性(seed heteromorphism)是指同一植株能够产生不同形态或不同萌发行为的种子的现象[1-2]。具有异型性种子的植物多为一年生草本植物且生命力较为顽强,多出现在干旱荒漠地和盐碱地等环境条件高度多变的地区[2-3]。异型性种子在外部形态特征、萌发、休眠特性等方面存在差异[4]。具有不同形态的种子在萌发时间和空间分布上的差异,能够降低外界环境条件对植物种群建成带来的风险。种子异型性是荒漠植物为适应异质环境而进化出的一种适应方式,在波动环境中,多型种子植物具有更高的适合度[5],这也是植物为了适应严酷的环境、维持种群稳定的结果。在我国干旱、半干旱区以及荒漠和盐碱地,分布着许多旱生、盐生植物类群,其中可能具有种子异型性的物种有待于发现[6],这些具有种子异型性的植物具有潜在的经济价值,能够起到修复生态的作用,极具开发利用价值。因此,研究异型性种子的休眠、萌发特性及生态适应对策是开发利用这些植物的基础。
蓼科植物大约有50 属1500 种,在北温带分布较多[7]。在我国有13 属,268 种,39 变种[8]。蓼科植物种类丰富,在药用、观赏绿化、牧草等方面应用前景广泛[9-10]。有些蓼属植物易成活,可以适应多变的环境条件,能够起到保护生态环境的作用。扁蓄(Polygonum aviculare)为蓼科蓼属一年生草本植物,茎直立或平卧,分布于全国大部分地区。花期4 月 -9 月,结果期6 月 - 10 月,瘦果卵状三棱形或双凸镜状[11],黑褐色或黄褐色,包于宿存的花被内或稍从花被中露出。扁蓄极耐严寒,能够抵抗干旱、水涝、高温等逆境条件,抗污染能力强,绿期长,是园林草坪绿化的优良地被植物[12]。
目前国内对扁蓄的研究主要集中在扁蓄的化学成分、药理作用、质量控制等方面[13-14],而有关扁蓄种子的萌发行为尚未见报道。野外观察到的扁蓄果实(种子)有两种形态,那扁蓄种子具有怎样的形态特征?二型性种子萌发特性有何差异?本研究以扁蓄种子为材料,对比其种子形态特征,并对两种种子进行萌发试验,探讨种子形态差异与萌发行为的关系,为合理开发利用这一野生地被植物资源提供基础资料。
1 材料与方法
1.1 研究材料
扁蓄的果实为瘦果,内含1 粒种子。扁蓄种子于2020 年10 月中下旬采集于准噶尔盆地南缘(43°45′44″~43°47′25″ N,87°33′16″~87°34′56″ E),该地区夏季炎热,干燥少雨,冬季寒冷漫长,昼夜温差大。
1.2 种子形态特征
随机选取大小不同的扁蓄20 株,观察种子的形状、颜色、宿存花被的形态。用万分之一天平测量种子重量,以1000 粒种子为一组,4 次重复,求平均值。用游标卡尺测量30 粒种子的长度和宽度,计算平均值。
1.3 种子在不同变温周期下的萌发
萌发试验均按4 次重复,每重复25 粒种子。种子放入垫有两层滤纸的培养皿中,加10 mL 蒸馏水,然后分别放入5 ℃/15 ℃、10 ℃/20 ℃、15 ℃/25 ℃、20 ℃/30 ℃、25 ℃/35 ℃ (黑暗12 h/光照12 h)的智能光照培养箱。种子萌发以胚根的出现为准,每天观测记录一次。
1.4 黑褐色种子的休眠特性及休眠的打破
1.4.1低温层积处理
将黑褐色种子均匀放入装有湿润河沙的塑料盒内,密封后置于4~5 ℃冰箱中,分别层积0、2、4、6、8 周。层积结束后,分别在5 ℃/15 ℃、10 ℃/20 ℃、15 ℃/25 ℃、20 ℃/30 ℃、25 ℃/35 ℃ (黑暗12 h/光照12 h)变温条件下进行试验,持续20 d。
1.4.2 划破种皮
用解剖刀轻轻划破黑褐色种子的种皮。将处理过的种子和未经任何处理的种子分别在5 ℃/15 ℃、10 ℃/20 ℃、15 ℃/25 ℃、20 ℃/30 ℃、25 ℃/35 ℃ (黑暗12 h/光照12 h)变温条件下培养20 d。
1.5 统计与分析
数据均采用SPSS 22.0 统计分析,结果以平均值 ±标准误表示。用Origin 18.0 对相关数据进行绘图。
2 结果与分析
2.1 种子形态
根据果实(种子)的颜色、大小、重量、形状等,将扁蓄的种子分为两种类型(图1、表1)。一种为卵形,具三棱,包于宿存花被内,与宿存花被近等长,大小约为1.91 mm × 1.34 mm,千粒重1.33 g,黑褐色。另一种为狭卵形,具三棱,伸出宿存花被片之外,大小约为3.15 mm × 1.40 mm,千粒重1.77 g,黄褐色。
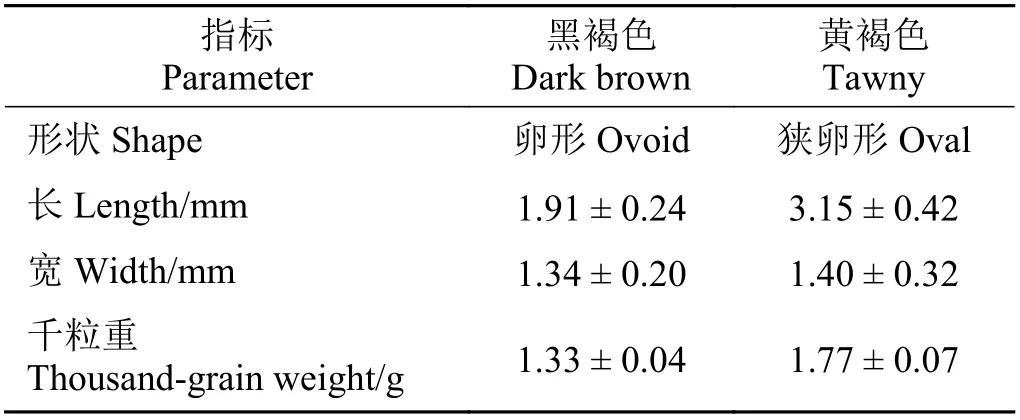
表1 扁蓄种子类型及特征Table 1 Seed types and characteristics of Polygonum aviculare
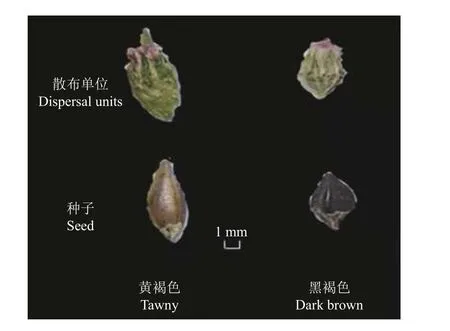
图1 扁蓄的散布单位及果实(种子)类型Figure 1 The dispersal units and seed types of Polygonum aviculare
2.2 种子在不同变温周期中的萌发行为
扁蓄黑褐色种子与黄褐色种子在不同变温周期下的萌发率表明,两种种子的萌发率存在显著差异(F= 2256.980,P= 0.000),两种类型种子的萌发特性显著不同(图2)。种子类型和变温对扁蓄种子的萌发有显著的交互作用(F= 42.758,P= 0.000) (表2)。
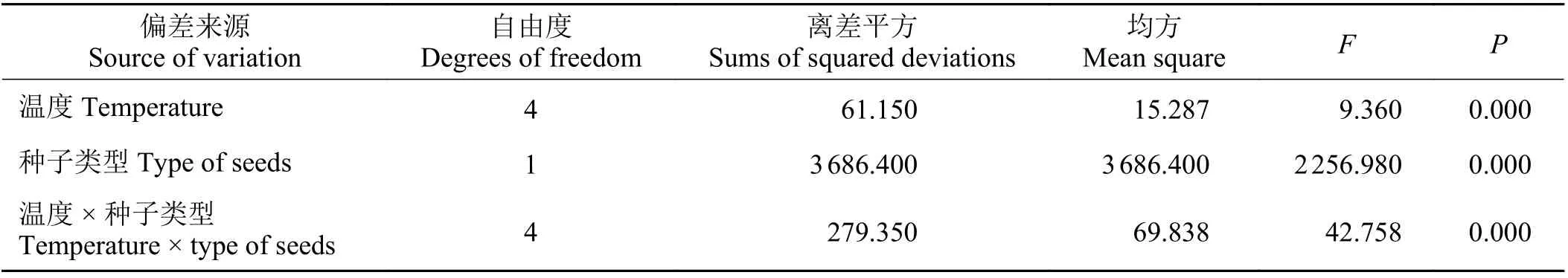
表2 温度和种子类型对扁蓄种子萌发率的影响Table 2 Analysis of variance for the effects of temperature, types of seed, and their interaction on the percentage germination of Polygonum aviculare seeds
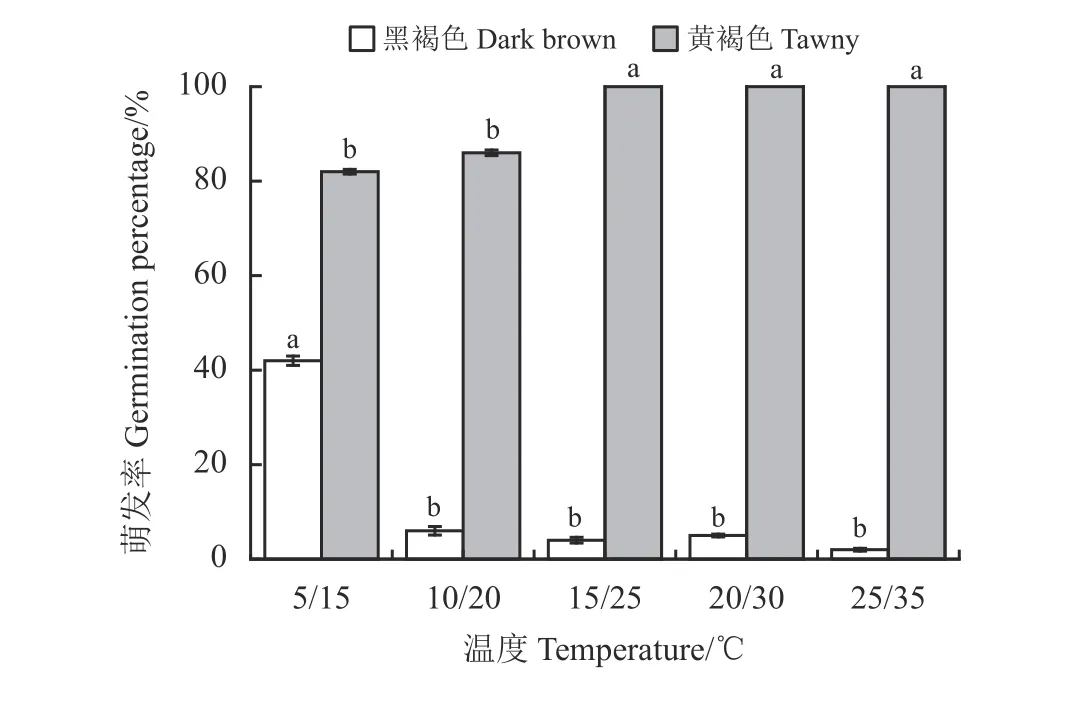
图2 扁蓄2 种类型种子在不同变温条件下的萌发率Figure 2 Percentage germination of the two types of Polygonum aviculare seed under different temperatures
在5 个不同变温周期下的萌发试验表明,黄褐色种子适宜萌发的温度范围较广,萌发的适宜温度为20 ℃/30 ℃和25 ℃/35 ℃,最终萌发率达100%,温度越高,萌发速度越快,高温度条件下的(15 ℃/25 ℃、20 ℃/30 ℃和25 ℃/35 ℃)种子在5 d 内萌发完毕,低温条件下的(5 ℃/15 ℃和10 ℃/20 ℃)种子则呈持续性萌发;而黑褐色种子萌发的适宜温度为5 ℃/15 ℃,在5 ℃/15 ℃条件下第6 天才开始萌发,最终萌发率为42%,在其他4 种温度下的萌发率均低于10% (图3),表明黑褐色种子具有休眠特性。
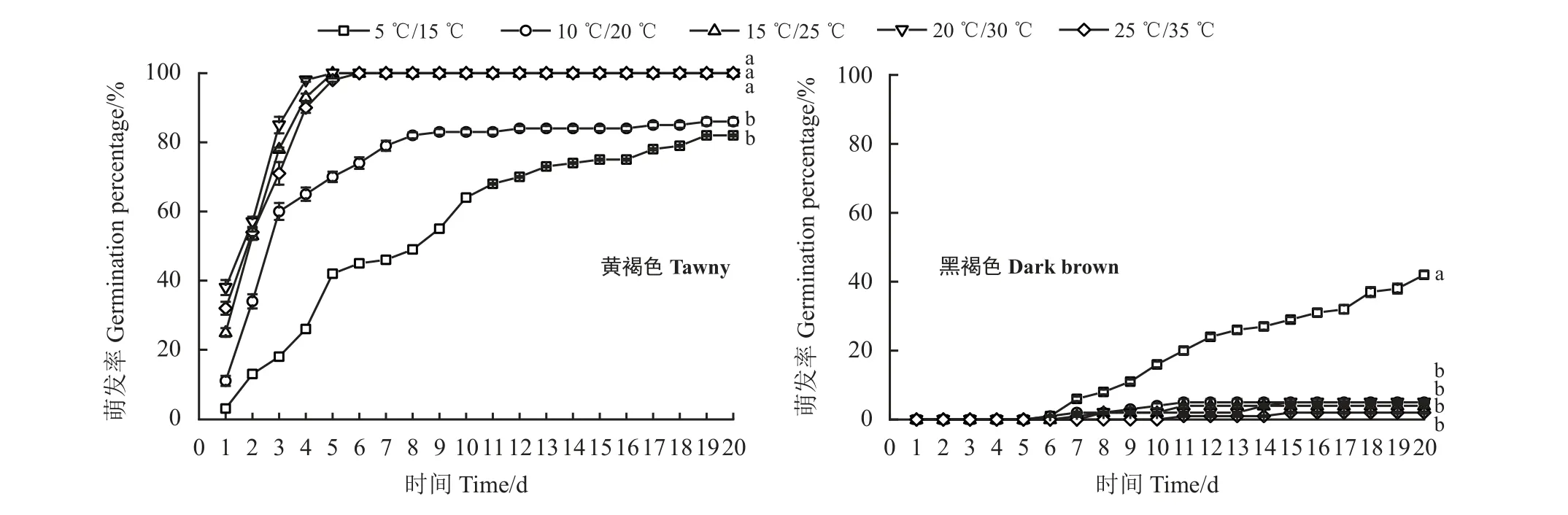
图3 扁蓄2 种类型种子在不同变温条件下的累积萌发率Figure 3 Accumulative germination of the two types of Polygonum aviculare seed under different temperatures
2.3 黑褐色种子的休眠特性及休眠的打破
2.3.1 低温层积处理
种子萌发特性试验结果表明,黑褐色种子在5 个变温条件下的萌发率较低,具有休眠特性。随层积时间的延长,在5 个变温周期下,黑褐色种子的萌发率显著提高(5 ℃/15 ℃:F= 38.811,P= 0.000;10 ℃/20 ℃:F= 32.057,P= 0.000; 15 ℃/25 ℃:F=25.496,P= 0.000; 20 ℃/30 ℃:F= 33.413,P= 0.000;25 ℃/35 ℃:F= 23.077,P= 0.000)。层积处理8 周后,在低温条件5 ℃/15 ℃萌发率在90%以上,表明低温层积处理能有效解除黑褐色种子的休眠(图4)。
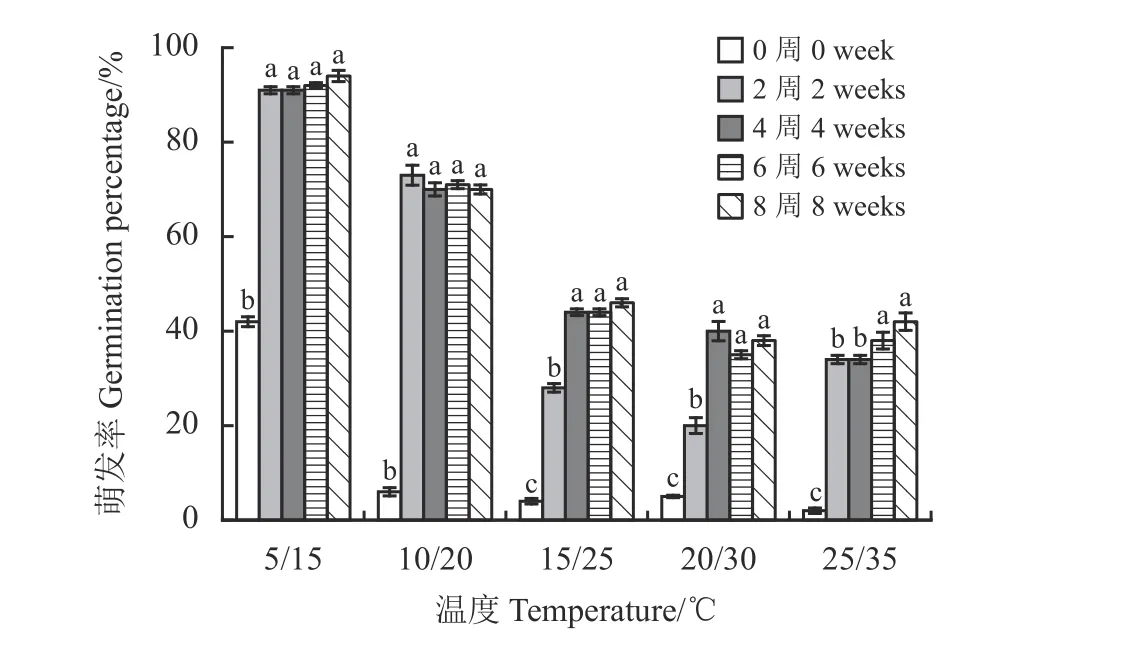
图4 不同变温条件下低温层积处理对扁蓄黑褐色种子萌发率的影响Figure 4 Effect of cold temperature stratification on the percentage germination of the dark brown seeds of Polygonum aviculare at different temperatures
2.3.2 划破种皮
和对照相比,划破种皮对黑褐色种子在5 个不同变温周期中的萌发率和萌发速率都有极显著的促进作用(图5)。划破种皮后,黑褐色种子在不同温周期下的萌发率(F= 392.442,P= 0.000)和萌发指数(F= 392.442,P= 0.000)均极显著提高,在5 个变温条件下的最终萌发率均在80%以上,这表明划破种皮可以有效解除扁蓄黑褐色种子的休眠(图5)。
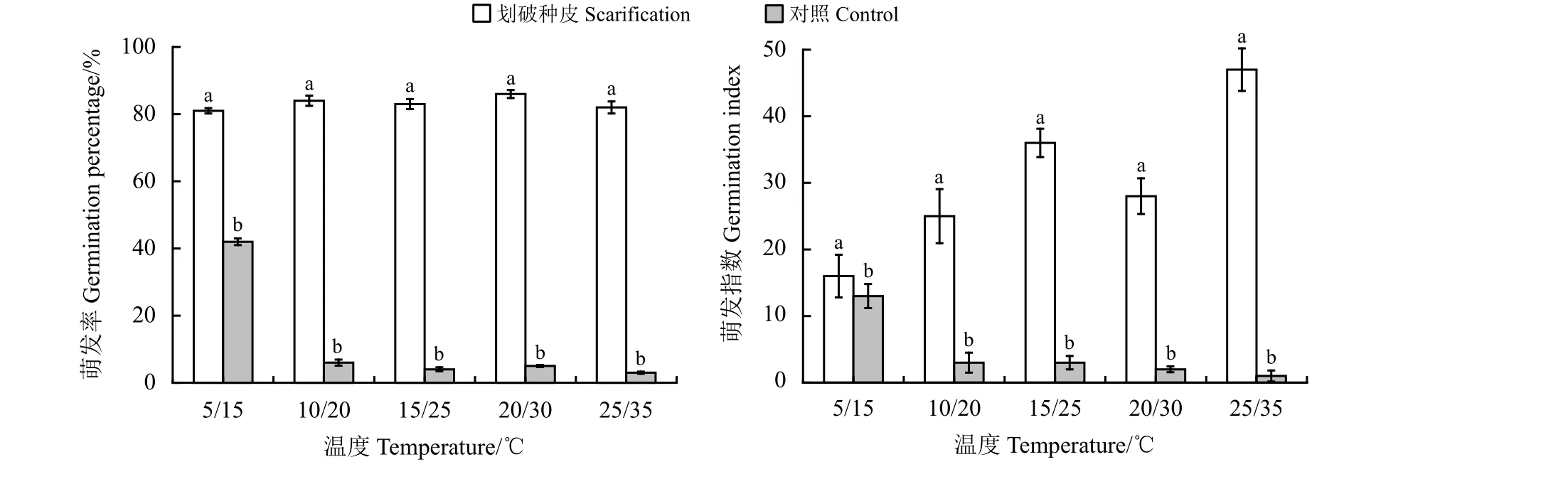
图5 不同变温条件下划破种皮对扁蓄黑褐色种子萌发率和萌发指数的影响Figure 5 Effect of scarification on the percentage germination and germination of the dark brown seeds of Polygonum aviculare at different temperatures
3 讨论
为了保证在多变复杂的胁迫生境中成功完成幼苗建植与种群繁衍[4],许多一年生植物的种子会发育出多种不同的形态及生理特性来影响其传播和萌发行为[15]。其中,种子异型性就是许多植物应对恶劣环境的一种有效策略。种子异型性常出现在那些生长于不可预知的环境中的植物上,是许多一年生植物适应异质环境条件的一种有效策略[16-18]。异型种子在形态特征、萌发、休眠及传播等方面均存在差异[19]。本研究表明,扁蓄同一植株上产生两种不同类型的种子,其在形状、大小、颜色上存在着明显差异,这使得黑褐色大种子和黄褐色小种子表现出不同的休眠和萌发特性。在准噶尔荒漠异苞滨藜(Atriplex micrantha)和藜(Chenopodium album)形成的不同形态种子中,颜色浅、质量重的种子不休眠,萌发率高,而颜色深、质量轻的种子休眠深,萌发率低[20-21]。本研究结果亦表明,扁蓄产生的黄褐色大种子,颜色浅、不休眠,小的黑褐色种子颜色深,休眠深。扁蓄具有的两种不同萌发能力的种子,能保证每个时期都有种子萌发,避免了种内竞争。
温度是种子萌发的重要调节者,适宜的温度能促进植物种子萌发和幼苗生长[22]。恶劣环境条件下有利于植物种子萌发和幼苗生长的时期短暂且难以预测。如果因春季某一次降水或温度的显著波动,损伤早期萌生的幼苗,则推迟萌发的小种子可补充因环境条件而导致的整个植物种群数量的降低。本研究表明,扁蓄黄褐色大种子在5 ℃/15 ℃、10 ℃/20 ℃、15 ℃/25 ℃、20 ℃/30 ℃、25 ℃/35 ℃这5 个变温条件中的萌发率都较高,萌发速度快,萌发温度范围广,即使在5 ℃/15 ℃的低温条件下萌发率也在75%以上。外界环境条件适宜时,黄褐色种子能够快速萌生大量的幼苗,这种在大的温度范围内快速萌发的能力使扁蓄在早春利用适量的降水迅速萌发出苗[5],从而迅速占据有利的生境。而黑褐色小种子在4 个变温条件下的萌发率始终维持在低的水平( < 20%),呈持续性萌发,具有一定的休眠特性,存在延迟发芽现象,就延迟发芽而言,前一年生产的一部分种子不能完全发芽,通过休眠将萌发的时间分散,在土壤中形成种子库,确保生长季内条件适宜时,能够在不同的时间段内均有萌发[23],可以避免种子全部萌发完的风险,补充因环境条件导致整个植物种群数量的降低[24],减少了植物生存的冒险性,有利于种群的持续生存。
种子休眠是植物长期进化而获得的抵抗外界不利因素的一种适应策略[25],在逆境条件下暂不萌发,一旦环境条件适宜时,便会萌发成幼苗,从而保证物种的延续[26]。本研究发现,扁蓄黄褐色种子在5 个变温周期下萌发率都很高,说明黄褐色种子无休眠,而黑褐色种子萌发温度范围窄,萌发持续时间长,只在5 ℃/15 ℃下萌发率较高,并且经过低温层积处理后,其适宜的萌发温度范围变宽,萌发率和萌发速率都得到提高,说明黑褐色种子存在休眠现象[27]。温带荒漠植物如新疆大赖草(Leymus racemosus)[28]、紫翅猪毛菜(Salsola affinis) [29] 也能够通过层积处理来解除休眠。扁蓄黑褐色种子当年成熟散落后,需经过冬天低温条件来解除休眠。早春地表温度回升,黑褐色种子利用融化的雪水开始萌发。伴随着气温升高,降水逐渐丰富,休眠解除的黑褐色种子大量萌发,补充整个植物种群数量。干旱恶劣环境中的植物种子能够持续性萌发,外种皮附着翅扩散到不同的距离或部分种子进入土壤种子库[29],占领最适生存生境,避免种群灭绝,这种现象可能是植物为应对严酷多变环境的一种生态策略[30]。本研究表明划破扁蓄黑褐色种子种皮能明显提高萌发率,说明黑褐色种子处于非深度生理休眠状态,这可能是由于黑褐色种子坚硬的种皮限制了胚根的伸长,从而影响萌发。扁蓄产生两种具有不同萌发策略的种子,黄褐色大种子不休眠,萌发的温度范围广,能在早春较低温度下萌发,而黑褐色小种子具有非深度生理休眠特性,萌发温度范围窄,更多的种子可能在后期温度逐渐回升后陆续萌发,这拓宽了扁蓄种子萌发的时间范围,降低种子单次萌发完的风险,能提高对异质生境的适应性,从而保证种群的成功扩散、定居和繁衍。
4 结论
扁蓄能够产生黄褐色和黑褐色两种形态的种子,种子萌发具有不同的行为。黄褐色种子萌发率高,萌发温度范围广,最适宜萌发温度是20 ℃/30 ℃和25 ℃/35 ℃,温度越高萌发速度越快,具有“冒险”的萌发策略,在适宜环境条件下快速萌发以扩大生存空间。而黑褐色种子具有非深度生理休眠特性,萌发温度范围较窄,具有“谨慎”的萌发策略,能够在时间和空间上避免与黄褐色种子萌生的幼苗产生竞争,分散植物生存的冒险性,增加子代存活的机会。划破种皮和低温层积处理可以有效破除扁蓄黑褐色种子的休眠。
猜你喜欢
植物资源与环境学报(2022年6期)2022-12-16 11:35:12
潍坊学院学报(2020年6期)2020-11-22 08:04:08
种子(2019年11期)2019-12-16 02:16:54
小天使·四年级语数英综合(2019年10期)2019-11-09 13:35:07
中国诗歌(2018年5期)2018-11-14 20:52:30
儿童故事画报·自然探秘(2018年9期)2018-11-06 02:39:26
许昌学院学报(2018年8期)2018-09-05 02:05:18
中国绿色画报(2018年5期)2018-08-07 07:12:26
知识窗(2017年8期)2017-08-23 02:10:26
雪花(2015年6期)2015-12-10 23:56:02
