
Rapid GFAP and Iba1 expression changes in the female rat brain following spinal cord injury
2022-08-08 02:11:26MawjMandwieJordanPiperCatherineGorrieKevinKeayGiuseppeMusumeciGhaithAlBadriAlessandroCastorina
中国神经再生研究(英文版) 2022年2期
Mawj Mandwie,Jordan A. Piper,Catherine A. Gorrie,Kevin A. Keay,Giuseppe Musumeci,Ghaith Al-Badri,Alessandro Castorina,,
Abstract Evidence suggests that rapid changes to supporting glia may predispose individuals with spinal cord injury (SCI) to such comorbidities. Here,we interrogated the expression of astrocyte- and microglial-specific markers glial fibrillary acidic protein (GFAP) and ionized calcium binding adaptor molecule 1 (Iba1) in the rat brain in the first 24 hours following SCI. Female Sprague-Dawley rats underwent thoracic laminectomy; half of the rats received a mild contusion injury at the level of the T10 vertebral body (SCI group),the other half did not (Sham group). Twenty-four hours post-surgery the amygdala,periaqueductal grey,prefrontal cortex,hypothalamus,lateral thalamus,hippocampus (dorsal and ventral) in rats were collected. GFAP and Iba1 mRNA and protein levels were measured by real-time quantitative polymerase chain reaction and Western blot. In SCI rats,GFAP mRNA and protein expression increased in the amygdala and hypothalamus. In contrast,gene and protein expression decreased in the thalamus and dorsal hippocampus. Interestingly,Iba1 transcripts and proteins were significantly diminished only in the dorsal and ventral hippocampus,where gene expression diminished. These findings demonstrate that as early as 24 hours post-SCI there are region-specific disruptions of GFAP and Iba1 transcript and protein levels in higher brain regions. All procedures were approved by the University of Technology Sydney Institutional Animal Care and Ethics Committee (UTS ACEC13-0069).
Key Words: affective disorders; astrocytes; glial fibrillary acidic protein; ionized calcium binding adaptor molecule 1; memory; microglia; neurotrauma; spinal cord injury
Introduction
Spinal cord injuries (SCI) can result in long-term and permanent deafferentation of cortical circuits of the central nervous system (CNS) (Wall and Egger,1971; Ziemann et al.,1998). Such changes can result in a substantial reorganization of cortical maps,exemplifying the plastic properties of the CNS (Aguilar et al.,2010). Sleep disturbance,anxiety,depression and cognitive dysfunction is highly prevalent in SCI patients (Davidoff et al.,1990; Kennedy and Rogers,2000; Biering-Sørensen and Biering-Sørensen,2001). This suggests strongly that following spinal cord trauma,in addition to changes in cortical circuits,other brain regions critical for the regulation of sleep,mood and cognition are also significantly impacted. For a complete understanding of the neurochemical bases of these changes in complex behaviors,it is essential to understand the changes in the brain triggered during the earliest stages of spinal cord injury,from which these longterm changes evolve.
Glial cells are the supporting cells of the CNS (He and Sun,2007). Alterations within the astrocyte and microglia compartments play significant roles in the onset and progression of several pathophysiological processes that can lead to a spectrum of affective dysfunctions (Öngür and Heckers,2004; Pav et al.,2008),as well as synaptic alterations (Honer et al.,1999; Coyle and Schwarcz,2000; Cotter et al.,2001; Scholz and Woolf,2007). Both astrocytes and microglia play major roles in shaping these CNS functions,and are likely to be the first cell populations primed following trauma,such as is associated with SCI. Glial fibrillary acidic protein (GFAP) is well established as the primary filament present in mature astrocytes within the CNS,where it is involved in modulating the structural stability,shape,and motility of the cells,as well as the cell-to-cell interactions with neurons (Eng,1985; Eng et al.,2000; Li et al.,2020). Ionized calcium binding adaptor molecule 1 (Iba1),is expressed in the cells of several tissues,including brain,testis,spleen and,to a lesser extent,in the kidneys and lungs. In the brain,Iba1 is expressed uniquely by microglia (Ito et al.,2001; Hwang et al.,2008),where it elicits actin-bundling activity and participates in membrane ruffling and phagocytosis when the microglia are activated (Ohsawa et al.,2004).
In a number of studies examining mood change and cognitive dysfunction identical to that seen in individuals with SCI,there are reports of regionally specific reductions in glial cell populations and/or glial activities in the amygdala,prefrontal cortex,hippocampus and periaqueductal gray (Öngür et al.,1998; Bowley et al.,2002; Imbe et al.,2012). Examining changes in GFAP and Iba1 transcript and protein levels may provide important insights into the temporal and topographical responses of glial cells occurring in higher brain regions after spinal cord injury.
In this study,we evaluated the hypothesis that within the initial 24 hours after an injury,SCI leads to rapid mRNA and protein changes in glial cells of discrete brain regions critical for the regulation of mood/emotion,stress responsivity,memory and decision-making. To answer this question,we investigated the gene and protein expression of GFAP and Iba1 in the amygdala,periaqueductal gray,prefrontal cortex,hypothalamus,thalamus and dorsal and ventral hippocampus of female rats 24 hours after SCI.
Materials and Methods
Ethics statement
All procedures were carried out with the approval of the University of Technology Sydney Institutional Animal Care and Ethics Committee (UTS ACEC13-0069),according to the guidelines set out by the National Health and Medical Research Council code of conduct for the use of animals in research (Nguyen et al.,2017).
Animals
Six adult female Sprague-Dawley rats (9 weeks old,body weight 250-300 g) were acquired from the Animal Resource Centre (Perth,WA,Australia). Rats were housed in cages on a 12-hour dark-light cycle with unlimited access to food and water. Each cage was provided with environmental enrichment. Animals were assigned randomly to either; (1) mild contusion SCI group (SCI;n= 3),or (2) sham surgery group (Sham;n= 3) (Figure 1).
Surgery and euthanasia
Rats were anaesthetised with 2% isoflurane (Thermo Fisher Scientific,NSW,Australia) in O2(flow rate of 1 L/min),once a surgical plane of anaesthesia was established,the fur above the thoracic region was shaved and iodine applied to the exposed skin. A subcutaneous injection of local anaesthetic (0.2 mL bupivacaine,Sigma-Aldrich,Castle Hill,NSW,Australia) was administered at the site of SCI or sham surgery. Each rat was given analgesics (buprenorphine hydrochloride -Temgesic 0.03 mg/kg,subcutaneously [s.c.]),antibiotics (cephazolin sodium 33 mg/kg,s.c.) and Hartman’s replacement solution (compound sodium lactate 15 mL/kg,s.c.). A midline incision was made from the mid to lower thoracic region and subcutaneous tissues cleared from the spinous process of the T10 vertebral body. A bilateral laminectomy of the T10 vertebrae exposed the dorsal surface of the spinal cord. Using a NYU/MASCIS weight-drop impactor,the vertebral column of each rat was stabilised with clamps attached to the T9 and T11 vertebrae and the exposed spinal cord subjected to a mild weight-drop contusion injury (6.5 mm,10 g,2.5 mm impactor head diameter). The surgical incision was closed in layers and sutured,and the animals were returned to a warmed cage where they were observed closely during recovery. During the next 24 hours,each rat received two further doses of analgesics (buprenorphine hydrochloride -Temgesic 0.03 mg/kg,s.c.),antibiotics (cephazolin sodium 33 mg/kg,s.c.) and Hartman’s replacement solution and underwent manual bladder expression (Nguyen et al.,2017).
At the end of this 24-hour period,the rats were deeply anaesthetised and euthanized using pentobarbital sodium (Lethabarb,1 mL/kg,intraperitoneally [i.p.]). The brain of each rat was carefully removed and transferred to HBSS buffer before being snap frozen in liquid nitrogen. The brains were stored at -80°C until microdissection.
Microdissections
The prefrontal cortex,amygdala,lateral thalamus,dorsal and ventral hippocampus,hypothalamus,and periaqueductal gray regions were microdissected using our previously described methods (Chiu et al.,2007; Castorina et al.,2019),and with reference to a stereotaxic atlas of the rat brain (Paxinos and Watson,2006). The brain was sectioned into smaller tissue blocks by making,three complete coronal cuts at specific antero-posterior (AP) levels using a sterile,chilled razor blade,cleaned in “RNA-ase Away”. The first coronal section was made at the anterior border of the optic chiasm (+0.3 mm anterior to bregma),the second at the posterior border of the interpeduncular fossa,and the third immediately posterior to the inferior colliculi as the midbrain aqueduct opens in the fourth ventricle (approximately -4.6 mm to -7.8 mm caudal to bregma). Each section created a tissue block that included one or more of the regions of interest,as detailed below:
Prefrontal cortex: In the most anterior tissue block we isolated the medial prefrontal cortex,the first coronal section was cut between bregma levels +4.68 mm and +2.52 mm (Figure 2A). Thereafter,we removed the anterior olfactory nucleus which occupies the ventral 1.5 mm of this section (Figure 2B) and then made two parasagittal cuts approximately 1 mm lateral to the midline abutting the forceps minor.
Amygdala: Similar to the prefrontal cortex,a 2-mm thick coronal section was cut between -1.92 mm and -3.96 mm caudal to bregma (Figure 2C). To obtain a block that included the entire amygdaloid complex,we used the opening of the lateral ventricle as a reference point to further dissect the triangular shaped area of the amygdala from each side located approximately 4 mm from the midline (Figure 2D).
Thalamus: To obtain lateral thalamus samples,we used the same tissue blocks used to isolate the amygdala. The hypothalamus was isolated from the remaining tissue block and using the internal capsule as the lateral boundary and the dorsal opening of the 3rdventricle as the upper boundary we micro-dissected the lateral thalamus from each side as two semi-rectangular tissue samples (~8 mm × ~2 mm) (Figure 2D).Dorsal hippocampus: was isolated from the same tissue block used to isolate the amygdala and lateral thalamus. The right and left dorsal hippocampi are easily identified and isolated. The tissue was separated from the cortical layer above and the corpus callosum (Figure 2D).
Ventral hippocampus: The ventral hippocampus was dissected from the most caudal tissue block and was isolated from a smaller tissue block approximately -4.6 mm to -7.8 mm caudal to bregma (Figure 2E). The ventral hippocampus was isolated from the surrounding cortex under a dissecting microscope using the thin layer of white matter surrounding the lateral boundary for reference,the tissue was removed with Dumont #7 forceps (Fine Science Tools,North Vancouver BC,Canada) (Figure 2F).
Periaqueductal gray (PAG): The tissue block used to isolate the ventral hippocampus was used to isolate the PAG. The PAG is a tubular-shaped region of the midbrain surrounding the cerebral aqueduct which resulted in a tissue sample of about 2 mm diameter (Figure 2F).
Each of the brain regions were weighed and immediately snap-frozen in liquid nitrogen and stored at -80°C for subsequent RNA extraction.
RNA extraction and cDNA synthesis
Each of the brain regions obtained from the SCI and sham groups were processed for RNA extraction,following TRI reagent manufacturer’s protocol,with minor modifications (Sigma-Aldrich). To extract RNA from samples we used 1 mL TRI reagent (Sigma-Aldrich) and 0.2 mL chloroform. We then precipitated the RNA with 0.5 mL 2-propanol at 12,000 ×gfor 15 minutes at 4°C (Castorina et al.,2014). We washed the pellet with 75% ethanol,left to air dry and re-dissolved in 30 μL milli-Q H2O. Final RNA concentration was measured with a spectrophotometer (Nanodrop ND-1000® spectrophotometer,Wilmington,DE,USA). To obtain single-stranded cDNAs we used the Tetro cDNA synthesis kit (Bioline,Sydney,NSW,Australia). We incubated 1 μg of total RNA with reverse transcriptase (200 U/μL); Oligo-(dT)18 primer (100 nM); 0.5 mM dNTP mix,RNase-inhibitor (10 U/μL) at 45°C for 40 minutes in a final volume of 20 μL. Temperature was finally increased to 85°C for 5 minutes to terminate the reaction.
Real time qPCR analysis
To analyze changes in steady-state levels of GFAP and Iba1 transcripts between SCI and Sham rats we used the CFX96 TouchTMReal-Time PCR Detection System (BioRad,Gladesville,NSW,Australia). The ribosomal protein 18S was used as the housekeeping gene. qPCR experiments were carried out by following a modified protocol,adapted from our previous study (Castorina et al.,2013). 3 μL of diluted cDNA (10 ng/μL),5 μL of SensiFAST SYBR®No-ROX master mix (Bioline),0.8 μL of 5 μM forward primer,0.8 μL of 5 μM reverse primer and 0.4 μL of MilliQH2O were added to a final volume of 10 μL per reaction. Differentially expressed genes were analysed using the ∆∆Ct method and are expressed as mean fold change (Schmittgen and Livak,2008). The ∆∆Ct of each sample was obtained by subtracting the calibrator (Sham) ∆Ct to the target sample ∆Ct and then applying the formula 2-∆∆Ct. Baseline measurements were set to 1. PCR product specificity was assessed by melting curve analysis,with each gene displaying an individual peak. The sequences of the genes used in this study are listed inTable 1.
Sodium dodecyl sulphate-polyacrylamide gel electrophoresis and Western blot
To obtain our protein lysates,brain tissues taken from different regions were homogenised using a sterilized conical pestle in RIPA Buffer (1:5 w/v,Sigma-Aldrich) containing a Protease Inhibitor cocktail (cOmpleteTM,Mini,EDTA-free Protease Inhibitor Cocktail,Sigma-Aldrich). Samples were then cleared by centrifugation at 12,000 ×gfor 10 minutes. Protein quantification was performed using the BCA assay (Thermo Fisher Scientific).
Denatured proteins (30 μg) were prepared by adding 4× Laemmli buffer (Bio-Rad,Gladesville,NSW,Australia) and β-mercaptoethanol (Sigma-Aldrich) and heating for 10 minutes at 70°C. Samples were run on a gradient Tris-glycine gel (4-20%,Bio-Rad),with a molecular weight ladder (Bio-Rad). Gels were transferred to a polyvinylidene difluoride (PVDF) membrane using the Trans-Blot Turbo instrument (Bio-Rad) (Giunta et al.,2010). Once transfer was completed,membranes were washed thoroughly with TBS + 0.1% Tween 20 (Sigma-Aldrich) (TBST 1×). Membranes were then blocked in 5% dry non-fat skim milk in TBST with slow agitation (50-60 r/min) for 1 hour at room temperature (RT).
Membranes were incubated with either GFAP (Abcam,Cambridge,Massachusetts,US,Cat# ab68428; dilution 1:2000) or Iba1 primary antibodies (Abcam,Cat# ab178846; dilution 1:500) in blocking buffer overnight at 4°C with slow agitation. Thereafter,membranes were washed 3× with TBST,followed by 3 × 5 minutes long washes. Finally,membranes were incubated with a secondary antibody (horse radish peroxidase-conjugated goat anti-rabbit IgG) for 1 hour at RT,diluted at 1:10,000 in blocking buffer. Membranes were finally washed to remove excess secondary antibody (Bucolo et al.,2012). Blots were revealed by chemiluminescence method (Clarity Western ECL,Bio-Rad) using the Bio-Rad ChemiDoc MP Imaging System (Bio-Rad).
Statistical analysis
This is an exploratory study. Given the intrinsic nature of these types of studies,we could not predicta priorifor any obvious changes in the pattern of gene and protein expression for any of the tested markers. Therefore,we conducted a priori power calculation using the online tool ClinCalc.com (https://clincalc.com/stats/samplesize.aspx) to calculate the right sample size on the assumption that a gene or protein expression fold change of ≥ 1.5 would have been considered biologically relevant. Based on this assumption and by estimating an inter-experiment standard deviation of 0.2 (20% variation),using a power of 80% and an alpha value of 0.05,our power calculations revealed thatn= 3 per group was enough to provide sufficient statistical power.
Data are reported as the mean ± SEM. Comparisons between groups were assessed using the unpaired Student’st-test.P≤ 0.05 was considered statistically significant. Data analyses were performed using GraphPad Prism version 8.0.0 for Windows (GraphPad Software,San Diego,CA,USA).
Results
Surgical induction of mild spinal cord injury
There were no adverse events during our surgical procedures and all rats recovered well after surgery. Rats with SCI showed signs of hind limb movement impairment,consistent with the location and severity of the injury. Sham rats (controls) did not show any signs of locomotor impairment as reported in our previous work (Nguyen et al.,2017).
Acute changes in GFAP mRNA and protein expression in the rat brain following SCI
The acute effects of SCI on the expression of GFAP in the amygdala and PAG,two regions pivotal in mediating emotional coping behaviours and that play important roles in the development of the behavioural dysfunction comorbid with injury and trauma (Keay and Bandler,2001; Phelps and LeDoux,2005; Mor et al.,2015; Motta et al.,2017),were evaluated.Figures 3Aand4Ashow that GFAP mRNA and protein expression levels were significantly upregulated in the amygdala compared to Sham controls (t10= 2.261 andt6= 3.061,P< 0.05),whereas in the PAG,the expression of GFAP mRNA and proteins were not significantly affected 24 hours after SCI (Figures 3Band4B;t10= 1.144 andt6= 0.872,P> 0.05 for GFAP).Figures 3Cand4Cshow that GFAP expression in the prefrontal cortex was unchanged 24 hours after SCI (t10= 0.630 for mRNA andt6= 0.936,P> 0.05 for GFAP),whereas hypothalamic GFAP mRNA and protein expression was significantly increased at this time (Figures 3Dand4D,t10= 2.490 andt6= 3.123,P< 0.05).
The thalamus receives a substantial spinal input and is the critical relay for somatosensory inputs to the cerebral cortex (Yuan et al.,2016); it also receives substantial inputs from spinal recipient brainstem regions,including projections from the ventro-lateral portion of the PAG carrying deep noxious inputs (Floyd et al.,1996),therefore we sought to determine if SCI altered GFAP expression in the thalamus. We report a surprising,and robust decline in GFAP transcript levels at this acute,24-hour time-point (Figure 3E,t10= 3.488,P< 0.01),which was further confirmed by protein analyses (Figure 4E,t6= 2.684,P< 0.01).
GFAP expression in the hippocampus was also significantly reduced in the dorsal hippocampus both at the mRNA (Figure 3F,t10= 2.500,P< 0.05) and even more robustly at the protein level (Figure 4F,t6= 4.029,P< 0.01),but not in the ventral hippocampus of SCI rats (Figure 3Gand4G,t10= 1.474 andt6= 1.659,P> 0.05).
Acute changes in Iba1 mRNA and protein expression in the rat brain following SCI
24 hours post SCI the mRNA and protein expression of the microglial marker Iba1 were unaffected in the amygdala (Figures 5Aand6A,t10= 0.514 andt6= 0.936,P> 0.05); the PAG (Figures 5Band6B,t10= 0.186 andt6= 1.059,P> 0.05); the prefrontal cortex (Figures 5Cand6C;t10= 1.342 andt6= 0.216,P> 0.05); the thalamus (Figures 5Eand6E,t10= 1.148 andt6= 0.406,P> 0.05) and the hypothalamus (Figures 5Dand6D,t10= 0.248 andt6= 0.307,P> 0.05). In contrast,Iba1 mRNA and protein expression levels in the hippocampus were reduced in both the dorsal hippocampus (Figures 5Fand6F,t10= 2.292 andt6= 3.739,P< 0.05 andP< 0.01,respectively) and the ventral hippocampus (Figures 5Gand6G,t10= 2.251 andt6= 2.453,P< 0.05).
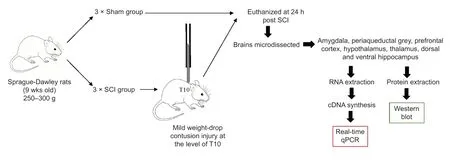
Figure 1|Flowchart of the experimental procedure.
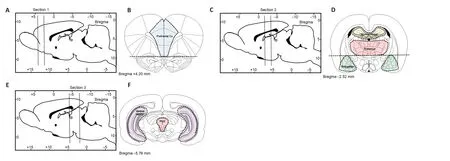
Figure 2|Schematic depicting rat brain microdissection procedures.
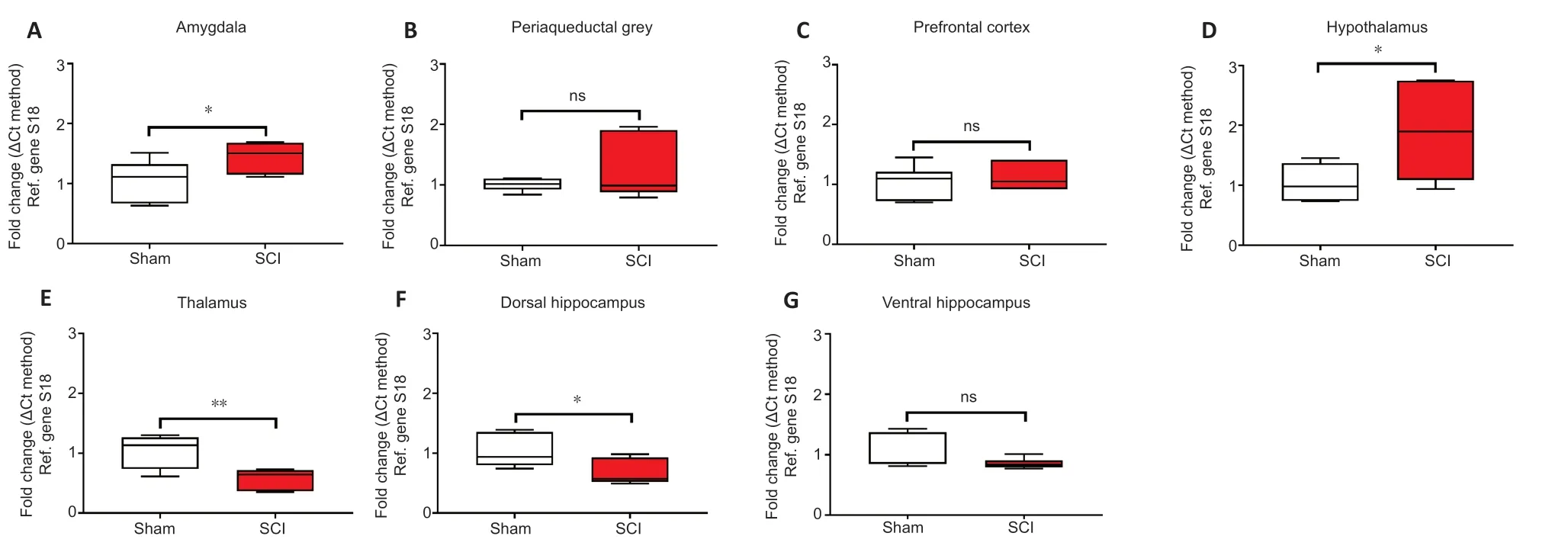
Figure 3|Real-time qPCR data showing the differential mRNA expression of GFAP in the amygdala (A),periaqueductal gray (B),prefrontal cortex (C),hypothalamus (D),thalamus (E),dorsal and ventral hippocampus (F and G) in sham and spinal cord injured rats.
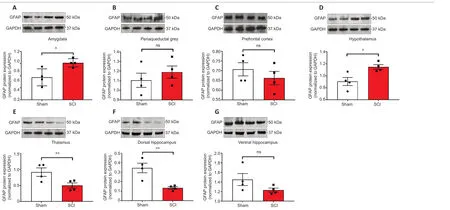
Figure 4|Western blot analyses of GFAP protein expression in the amygdala,periaqueductal gray,prefrontal cortex,hypothalamus,thalamus,ventral & dorsal hippocampus of sham-injured (Sham) and spinal cord injury rats.
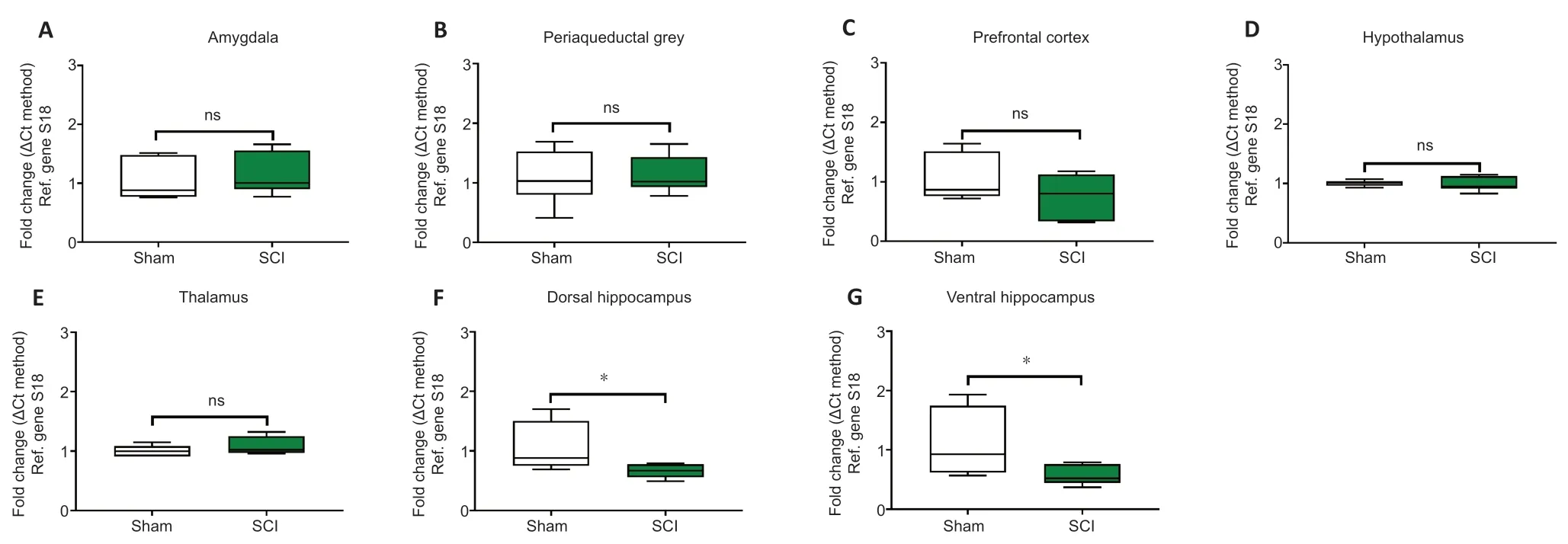
Figure 5|Real-time qPCR data showing the differential mRNA expression of Iba1 in the amygdala (A),periaqueductal gray,(B) prefrontal cortex (C),hypothalamus (D),thalamus (E),dorsal and ventral hippocampus (F and G) of sham-injured (Sham) and spinal cord injury rats (SCI).
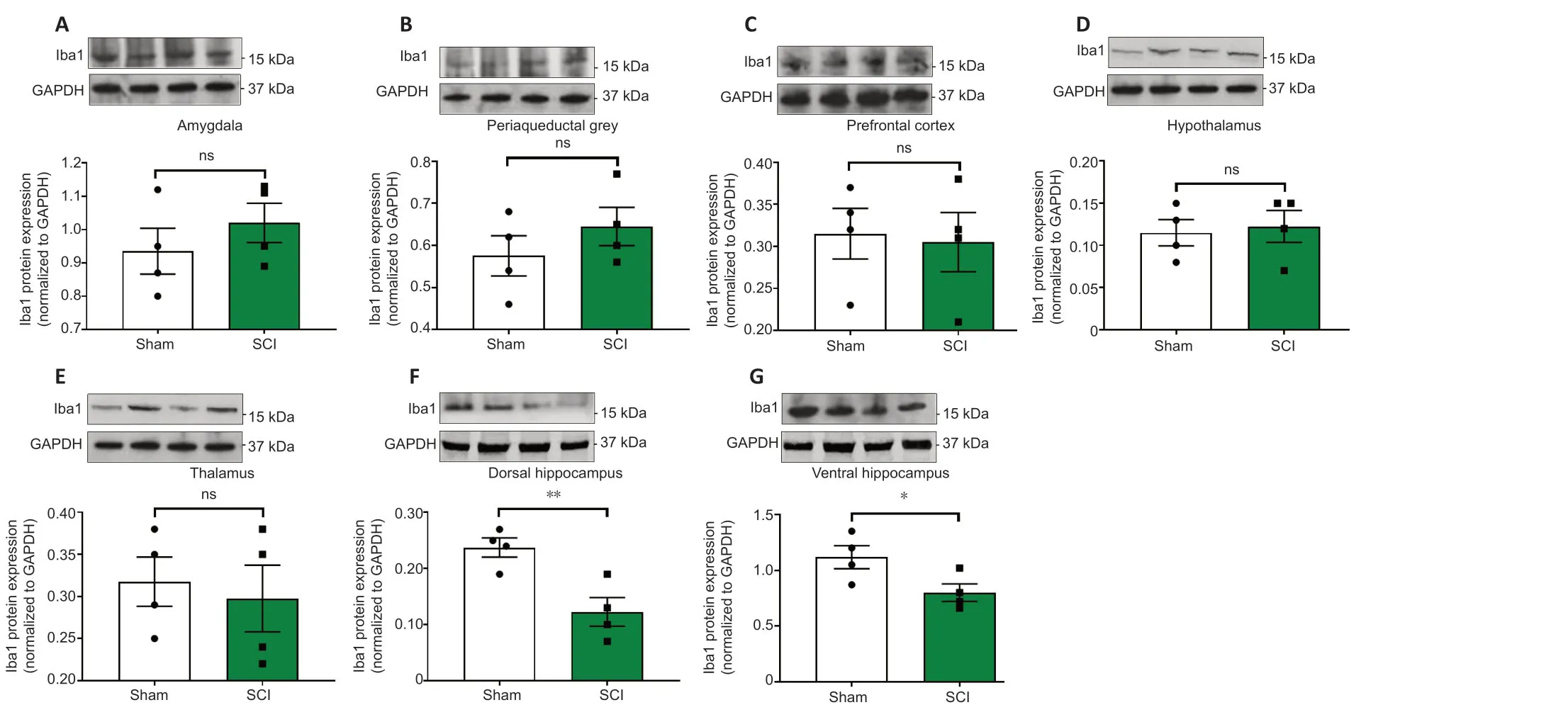
Figure 6|Western blots analyses of Iba1 protein expression in the amygdala,periaqueductal gray,prefrontal cortex,hypothalamus,thalamus,ventral & dorsal hippocampus of sham-injured (Sham) and spinal cord injury rats.
Discussion
In this study,we identified early changes in GFAP and Iba1 mRNA and protein expression levels in the female rat brain following a mild SCI. We interrogated astrocyte- and microglial-specific cell markers,as our main goal was to detect early disruptions within the glial compartment following SCI. Given the complex architectural organization of the brain and the differential involvement of specific brain regions in the development of the comorbidities associated with spinal cord injury our focus was on brain areas controlling the affective,cognitive and sensory responses to traumatic stressors,for these measurements.
There is documented evidence of sexual dimorphism in the behavioral responses to acute stress and as in several types of high order cognitive and affective functions (Rubinow and Schmidt,2019). Apparently,these differences account for hormonal,sex chromosomes and their interaction with the environment (Rubinow and Schmidt,2019),with studies indicating that female rats,as opposed to males,develop distinct coping strategies in response to stress (Youssef et al.,2018). Interestingly,in a recent work it has been demonstrated that stressed-susceptible brain regions such as the prefrontal cortex or the hippocampus exhibit higher activation patterns in male vs. female in rats exposed to acute immobilization stress,but not in the forced swimming test (Sood et al.,2018). These data pinpoint the sexually dimorphic response strictly depends on the specific nature of the stressor,a topic that warrants further investigations. In this exploratory study,we utilised female rats as these are conventionally used in spinal cord injury studies due to their better compliance to the surgical procedure compared with male rats (Datto et al.,2015). A further reason was ethical,as in our experience male rats subjected to SCI often develop a severe form of autotomy (self-mutilation) directed to their hind-limbs and sometimes genitalia,an adverse event that we have never observed in female rats. Interestingly,the exaggerated autotomy in male SCI rats also seems to occur after neurectomy of peripheral nerves,as shown in other studies (Wagner et al.,1995).
Our analyses identified regionally specific changes in GFAP gene and protein expression in several supraspinal structures. To contrast,changes in Iba1 expression were restricted to the dorsal and ventral hippocampus,brain regions critical in integrating memory formation,spatial navigation and emotional regulation (Schultz and Engelhardt,2014). To our knowledge,this is the first evidence describing acute changes in supraspinal GFAP and Iba1 mRNA and protein regulation,24 hours post-SCI.
SCI and GFAP expression in the brain
Accumulating evidence shows that the activity of astrocytes is crucial in determining the behavioral outputs of both the amygdala and hypothalamus,via a process that involves the selective regulatory activity of specific synapses by activated astrocytes (Martin-Fernandez et al.,2017; Chen et al.,2019). At the cellular level,astrocytes express receptors for both noradrenaline (β2-adrenergic receptors) and glucocorticoids (Hertz et al.,2010; Jauregui-Huerta et al.,2010),each of which play different roles in modulating the calcium influx and ATP release of individual astrocytes (Chen et al.,2019).
During the initial response to an acute stressor such as a traumatic injury,noradrenaline release precedes that of the glucocorticoids,cortisol and/or corticosterone (Pearson-Leary et al.,2015; Chen et al.,2019). This suggests that increased noradrenergic activity likely predominates in the immediate phase of the response to traumatic spinal cord injury. In view of this,and considering the different temporal activation patterns and anatomical distributions of astrocytic β2-adrenergic receptors and glucocorticoids in the brain (Gao et al.,2016),it is possible that the regionally distinct patterns of GFAP mRNA and protein regulation that we observed after SCI might be linked to differential exposure of these discrete brain regions to increasing norepinephrine (NE) and glucocorticoids after the spinal cord injury.
GFAP gene and protein levels were significantly increased in the stress-responsive amygdala and hypothalamus,each of which receives strong afferent drive from the noradrenergic locus coeruleus (Palkovits et al.,1980; Kawakami et al.,1984). The locus coeruleus is reliably activated by acute stressors and it is tempting to suggest that a strong activation of this noradrenergic region immediately following SCI,could lead to significant release of norepinephrine in the amygdala and hypothalamus,leading to increased astrocyte activity,as reflected by the induction of GFAP expression reported here.
In contrast,GFAP transcripts and proteins were reliably decreased in the lateral thalamus. The lateral thalamus,encompassing key somatosensory thalamic relays,is a critical source of somatosensory inputs both between different subcortical areas and the cortex (Herrero et al.,2002). Unlike the amygdala and hypothalamus,the thalamus is not strongly regulated by ascending noradrenergic pathways (Simpson et al.,2006). It is however particularly sensitive to the effects of deafferentation triggered by SCI,and many populations of thalamic neurons respond to SCI by immediately increasing their firing activities (Alonso-Calvino et al.,2016). It is therefore possible that the decline in GFAP mRNAs and proteins we report reflects a compensatory mechanism in which the astrocytes surrounding hyperactive thalamic neurons diminish their activity in an effort to dampen the effects of deafferentation.
The hippocampus is particularly vulnerable to both acute and chronic stressors,including those triggered by physical trauma (Jing et al.,2017). This brain structure has a highly conserved architectural organisation along its dorso-ventral axis (Schultz and Engelhardt,2014),with the dorsal hippocampus critical for spatial navigation and memory and the ventral hippocampus regulating emotional processing and expression (Amaral and Witter,1989). Despite the architectural similarities,several studies have pinpointed significant differences in the transcriptional and proteomic profiles of the dorsal and ventral sub-regions of the hippocampus in response to stress (Maggio and Segal,2009; Pierard et al.,2017; Floriou-Servou et al.,2018). An important observation of this study was that SCI significantly reduced both gene and protein expression of the astrocytic marker GFAP in the dorsal,but not the ventral part of the hippocampus. Reduced GFAP expression has been previously reported in the hippocampus and prefrontal cortex in a rat model of depression (Eldomiaty et al.,2020). By contrast,mild cortical contusion has shown to increase hippocampal GFAP mRNA levels as early as after 12 hours post-injury (Hinkle et al.,1997). In line with our hypothesis that a spinal trauma can predispose an individual to the development of comorbid behavioral dysfunctions,it is perhaps not surprising that GFAP expression is reduced.
SCI and Iba1 expression in the brain
Iba1 is a microglia-specific calcium binding protein bothin vitroandin vivo,whose expression reflects cellular polarization state (Ito et al.,1998). In this study,we observed that Iba1 mRNA and protein expression were selectively reduced in the dorsal and ventral regions of the hippocampus after SCI. At first consideration these findings appear counterintuitive,however,the ‘shock’ suffered by these vulnerable brain regions as a consequence of the physical trauma of spinal cord injury,might well-reflect the impacts of the shock evoked in animal models evaluating the central effects of electroconvulsive therapy (Jinno and Kosaka,2008). Our data suggest that spinal cord trauma triggers immediate plastic changes in the hippocampus that are associated with attenuated microglia activity,in this acute post injury phase. Considering the recently identified role of microglia in synapse turnover (Wang et al.,2020),it is not unreasonable to suggest that attenuated Iba1 expression might reflect the pathological increase in neuronal plasticity that occurs after a traumatic experience such as SCI.
Conclusions
Our data provides evidence for early changes in glial activity in several brain regions involved in the development of behavioral comorbidities following SCI. Glial activity changes show clear regional specificity,and it is the activity of astrocytes that is most strongly affected during this period. We also identified attenuated Iba1 mRNA and protein expression in the hippocampus,which is consistent with rapid and adaptive neuroplasticity in this region. However,whilst the changes in the expression of the glial markers were remarkable,it should be noted that the associated comorbid changes in higher order cognitive functions and affective behaviors may require long-term modulations occurring at molecular,cellular and systemic levels. Therefore,additional investigations addressing the changes in glial activity over time are warranted. Nonetheless,these findings provide the first evidence of early supraspinal glial expression changes following spinal cord injury which could lay the foundations for the subsequent development of affective and cognitive dysfunction that is comorbid with SCI in many individuals (summarized inTable 2).

Table 2|The topographical disruptions of GFAP and Iba1 mRNA and protein expression levels seen in response to spinal cord injury after 24 hours
Acknowledgments:We would like to thank Ms Mercedes Ballesteros (University of Technology Sydney,NSW,Australia) and Ms Sarah Osvath (University of Technology Sydney,NSW,Australia) for the technical support to the researchers of the Laboratory of Cellular and Molecular Neuroscience (LCMN).
Author contributions:Writing—original draft preparation: JAP;conducted the experiments and substantial contributed to original draft preparation: MM; conducted the experiments,reviewed and the edited the paper: CAG; writing—review and editing: KAK; helped in the methodology,formal analyses and final revision: GM and GAB; conceived the study,obtained and administered the funding,revised the manuscript: AC. All authors approved the final version of the manuscript.
Conflicts of interest:The authors declare no conflicts of interest.
Financial support:The University of Technology Sydney (UTS) Start-up Research Grant 2018 funded this study (to AC).
Institutional review board statement:All procedures were carried out with the approval of the University of Technology Sydney Institutional Animal Care and Ethics Committee (UTS ACEC13-0069).
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement:Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal,and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License,which allows others to remix,tweak,and build upon the work non-commercially,as long as appropriate credit is given and the new creations are licensed under the identical terms.

- 中国神经再生研究(英文版)的其它文章
- A Drosophila perspective on retina functions and dysfunctions
- Celeboxib-mediated neuroprotection in focal cerebral ischemia: an interplay between unfolded protein response and inflammation
- Pramipexole,a dopamine D3/D2 receptor-preferring agonist,attenuates reserpine-induced fibromyalgia-like model in mice
- Effects of delayed repair of peripheral nerve injury on the spatial distribution of motor endplates in target muscle
- Neurorehabilitation using a voluntary driven exoskeletal robot improves trunk function in patients with chronic spinal cord injury: a single-arm study
- Gene and protein expression profiles of olfactory ensheathing cells from olfactory bulb versus olfactory mucosa