
水稻川香29B近等基因导入系苗期根系形态、叶片生理生化特性与抗旱性的关系
2022-07-28 05:37:50李其勇朱从桦李星月武丙琳符慧娟
干旱地区农业研究 2022年4期
李其勇,朱从桦,李星月,易 军,武丙琳,符慧娟,张 鸿
(1. 四川省农业科学院植物保护研究所/农业部西南作物有害生物综合治理重点实验室,四川 成都 610066; 2. 四川省农业科学院作物研究所,四川 成都 610066)
水稻是我国最主要的粮食作物之一,常年种植面积3 000万公顷,产量2亿吨左右,是世界上水稻产量第一、种植面积第二的国家[1]。水稻栽培耗水量大,有研究得出每生产1 kg谷粒需要3 000 L水[2], 而我国水资源十分贫乏,且时空分布极不均衡。水资源短缺和频繁的干旱日益成为制约水稻生产的重要环境因素[3],测算研究表明干旱影响下,未来全球水稻产量的平均预期损失率可能达到13.1%(±0.4%),水稻干旱风险波动幅度和干旱风险增长的区域比例都将显著增加[4]。干旱胁迫可导致水稻种子发芽率下降[5]、幼苗成苗率降低[6],在分蘖期、孕穗期、灌浆成熟期等主要生育期遭受干旱胁迫,水稻的光合生产能力显著下降,产量和稻米品质均有所降低[7-8],不利于大田生产。为应对日益严重的干旱风险对水稻生产的挑战,在水资源时空因素限制下,应用抗旱品种是解决抗旱栽培的重要途径之一,而筛选鉴定抗旱水稻品种或育种材料,科学评价及准确预测水稻的抗旱性对水稻抗旱育种和栽培都具有重要意义。
水稻抗旱机制复杂,不同品种或材料在不同水分胁迫环境、不同生育阶段,其表现出的抗旱机制也存在差异[9-11],水稻对水分胁迫适应亦是多种途径共同作用的结果[12-14]。正由于此,不同研究者提出的水稻抗旱性鉴定指标很多,且不统一,主要包括抗旱系数、抗旱指数、产量相关指标、外观形态表型指标(芽、根系、叶片等)、生长发育指标、生理生化代谢指标[15-18]。相较于全生育期干旱胁迫处理,苗期进行干旱胁迫处理可将抗旱鉴定工作前移,具有工作量小、耗时短、可大批量进行的特点,得到了较多关注。苗期主要采用反复干旱法进行干旱胁迫研究[6,19-20],筛选到了部分鉴定指标,如最大根长、根数、根鲜重、叶鲜重、超氧化物歧化酶(SOD)活性、还原性谷胱甘肽(GSH)含量等指标,并评价获得了一批抗旱材料。但苗期进行抗旱鉴定评价指标仍未统一,且多是采用水稻品种,在分析过程中多是形态指标、生理指标,为进一步研究干旱胁迫对水稻材料苗期的影响并筛选抗旱鉴定指标,以5份近等基因导入系及其轮回亲本川香29B为材料,在温室内进行苗期反复干旱胁迫,测定苗期水稻在水分胁迫下的形态、渗透调节物质、激素类物质、保护酶活性等多类指标,分析指标与抗旱性关系,筛选水稻苗期抗旱性鉴定的指标,并利用入选指标建立水稻苗期抗旱预测方程,为水稻苗期抗旱性鉴定及预测提供参考。
1 材料与方法
1.1 供试材料
供试材料为5份近等基因导入系及其轮回亲本川香29B(优质籼稻),由四川省农业科学院作物所提供。以保持系川香29B(优异籼稻保持系,已组配出多个优质高产品种在生产上应用)为轮回亲本,ASOMINORI(全球水稻分子育种计划的核心种质,耐旱性强)作供体亲本,从连续回交2代后再自交的材料中通过初步耐旱试验获得材料C1、C2和C3;从连续回交3代后再自交的材料中通过初步耐旱试验获得材料C4和C5。以此5份川香29B近等基因导入系(Chuanxiang 29B NIILs)为供试材料,以保持系川香29B作为对照(表1)。
表1 川香29B近等基因导入系材料及编号Table 1 Material codes of Chuanxiang 29B near-isogenic introgression lines
1.2 试验设计
于2017年6—7月在中国水稻研究所富阳基地(30°5′N,119°55′E)温室内开展试验,富阳属中纬度亚热带季风气候,6、7月的平均气温、降雨量、日照时长分别为26.5℃和28.6℃、112 mm和218 mm、98 h和220 h,试验土质为黏性水稻土。挑选籽粒饱满的水稻种子,浸种48 h,于37℃恒温培养箱中催芽,待种子露白后播种。采用大钵育秧,每钵3粒,置于大棚培育出苗,每个材料播种12盘,每盘144钵,长至3叶1心时,选择生长均匀一致的秧苗,进行干旱胁迫。采用T×C二因素完全随机试验设计,T因素为水分管理,设置2个水平,CK为正常浇水管理,T为定期进行干旱胁迫;C因素为供试材料,即6个供试水稻材料,见表1,共计12个处理,每个处理设置3次重复。干旱胁迫(T)包括2次干旱处理过程,其中第1次干旱处理,记为T1:每个材料选择3盘,停止浇水,当所有品种叶片在干旱胁迫后中午前后出现萎蔫,叶片出现严重枯萎(所有叶片均严重卷曲成针状),每个材料出现整株“暂时枯死”,作为第1次干旱胁迫结束点;第2次干旱处理,记为T2:第1次干旱取样调查结束,立即浇透水,后续不再浇水,待所有品种再度萎蔫卷曲成针状,50%叶尖出现枯黄,每个材料品种超过5%的植株出现整株“暂时枯死”,作为第2次干旱胁迫终点。
1.3 测定项目和方法
分别在第1次干旱处理结束、第2次干旱处理结束时,每个处理选取代表性植株进行取样和测定。
1.3.1 株高、干物质量和根系形态指标 每盘取30株秧苗,用直尺测定株高,将秧苗分为地上部和根系于105℃杀青30 min,80℃烘干至恒重,称重计算地上部和根系干重,并计算根冠比;根系流水洗净后采用Epson Expression 1000xl根系扫描仪,辅以WinRHIZO软件,测量根系总长、总体积、总表面积和平均直径。
1.3.2 叶片SPAD值和色素含量 每盘选取30株秧苗,用SPAD-502型叶绿素测定仪,测定SPAD值。然后剪取顶部第一展叶,混合后称取0.1 g,加入10 ml提取液(乙醇∶丙酮=1∶1),浸提24 h后用分光光度法测定叶绿素a(Chlorophyll a)、叶绿素b(Chlorophyll b)、类胡萝卜素(Carotenoid)含量。
1.3.3 叶片渗透调节物质和抗氧化物质 各处理均选取秧苗顶部第1展叶测定。可溶性糖含量、游离氨基酸含量、脯氨酸(Proline, Pro)含量、丙二醛(MDA)含量、还原型谷胱甘肽(GSH)含量、可溶性蛋白含量采用苏州科铭生物技术有限公司试剂盒以微量法测定,Vc含量采用上海源叶生物科技有限公司ELISA试剂盒测定,重复3次。
1.3.4 叶片激素含量测定 各处理选取秧苗顶部第1展叶,采用上海源叶生物科技有限公司的ELISA试剂盒测定生长素(IAA)、脱落酸(ABA)、细胞分裂素(CTK)、赤霉素(GA)和乙烯(ETH)含量,重复3次。
1.3.5 叶片酶活性测定 各处理均选取秧苗顶部第1展叶测定。采用上海源叶生物科技有限公司ELISA检测试剂盒测定1-吡咯啉-5-羧酸合成酶(Pyrroline 5 carboxylate synthetase, P5CS)、鸟氨酸转氨酶(Ornithine δ-aminotransferase, δ-OAT)、脯氨酸脱氢酶(Proline dehydrogenase, ProDH),采用苏州科铭生物技术有限公司试剂盒以微量法测定超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性,均重复3次。
1.4 数据分析
用Microsoft Office Excel 2013和DPS 14.50进行数据整理、运算和分析,用最小显著差法LSD(P<0.05)检验,采用Orgin9.1作图。
反复干旱存活率(%)=(第1次干旱后的存活率+第2次干旱后存活率)/2
相关性分析、回归分析采用指标相对值进行分析,计算方法:
指标相对值=干旱胁迫处理指标测定值/正常水分处理指标测定值
2 结果与分析
2.1 干旱胁迫对水稻川香29B近等基因导入系苗期株高、干物质积累量及根冠比的影响
从表2可得,第1次干旱胁迫后,6份供试材料平均株高比对照略有增加,两次干旱胁迫后株高与CK均无显著差异。两次干旱胁迫均显著降低了水稻地上部干重,第2次干旱胁迫后降低幅度更大;两次干旱胁迫均显著提高了水稻根系干重和根冠比,第2次干旱胁迫后增加幅度更大。从供试材料来说, C3、C4、C6的地上部干重在第1次干旱胁迫后表现增长,在第2次干旱胁迫后降低幅度较其他材料小,C4、C5、C6的根系干重在两次干旱胁迫后增幅均排前三名,C1、C5、C6根冠比在两次干旱胁迫后增幅均为前三名。综上,干旱胁迫后秧苗地上部干物质降低,而根系干重显著增加,根冠比提高,这是水稻苗期适应干旱环境的重要途径。

表2 干旱胁迫对川香29B近等基因导入系苗期株高、干物质积累量及根冠比的影响Table 2 Effects of drought stress on plant height, dry matter accumulation and root-shoot ratio of Chuanxiang 29B NIILs at seedling stage
2.2 干旱胁迫对水稻川香29B近等基因导入系苗期根系形态的影响
从表3可得,平均总根长在第1次干旱胁迫后显著增长,在第2次干旱胁迫后显著降低,根表面积、根粗、根体积在两次干旱胁迫后均显著降低,表明相对较轻的干旱处理(第1次干旱胁迫)促进了总根长的伸长,但对其他根系指标均为抑制作用,较重的干旱胁迫(第2次干旱胁迫)则对根系完全表现为抑制效应,但抑制效应存在材料差异。C1、C2、C6的总根长两次干旱胁迫下均表现为增长,第1次干旱胁迫后增长幅度更高,同时这3份材料的根表面积在第1次干旱胁迫后增长,增幅为8.41%~31.44%;6份供试材料的根粗、根体积在两次干旱胁迫后均表现减小。说明干旱胁迫下,水稻根粗、根体积均受抑,通过总根长和根表面积增大而适应干旱环境,但不同材料间适应能力不同。

表3 干旱胁迫对川香29B近等基因导入系根系形态指标的影响Table 3 Effects of drought stress on root morphological indicators of Chuanxiang 29B NIILs
2.3 干旱胁迫对水稻川香29B近等基因导入系叶片SPAD值和色素含量的影响
从表4可得,两次干旱胁迫下水稻幼苗叶片SPAD值、叶绿素a含量、叶绿素b含量平均值均显著降低,第1次干旱胁迫后类胡萝卜素含量显著降低,而第2次干旱胁迫后类胡萝卜素含量显著提高。从供试材料来说,6份供试材料的叶片SPAD值、叶绿素a含量、叶绿素b含量在两次干旱胁迫后均表现降低,表现出明显的抑制效应;C2的类胡萝卜素含量在两次干旱胁迫后均表现增长,C3的类胡萝卜素含量在第1次干旱胁迫后增长,在第2次干旱胁迫后减少,C4、C5、C6则正好相反。说明干旱胁迫降低叶片SPAD值、叶绿素a含量、叶绿素b含量,导致干物质合成减弱而抑制幼苗生长。

表4 干旱胁迫对川香29B近等基因导入系叶片SPAD值和色素含量的影响Table 4 Effects of drought stress on SPAD value and pigment content in leaves of Chuanxiang 29B NIILs
2.4 干旱胁迫对水稻川香29B近等基因导入系叶片渗透性物质及氧化还原物质的影响
从表5可得,两次干旱胁迫下,水稻幼苗可溶性糖、氨基酸平均值均表现降低,第1次干旱胁迫后降幅大于第2次干旱胁迫;可溶性蛋白含量、脯氨酸含量平均值均表现增长,第1次干旱胁迫后差异均达显著水平;还原型谷胱甘肽含量第1次干旱胁迫后显著提高,在第2次干旱胁迫后显著降低;Vc含量在两次干旱胁迫后均显著降低,第2次干旱胁迫降幅更大;丙二醛含量在两次干旱胁迫后均显著提高,第1次干旱胁迫增幅更大。
2.5 干旱胁迫对水稻川香29B近等基因导入系叶片激素含量的影响
从表6可得,干旱胁迫下,生长素、细胞分裂素、赤霉素均表现降低,其中赤霉素两次干旱胁迫后均显著降低,脱落酸、乙烯均表现为显著升高,5个指标在第2次干旱胁迫后差异均达显著水平。从供试材料来说,6份材料的生长素、细胞分裂素、赤霉素在两次干旱胁迫后均表现降低,而脱落酸、乙烯在两次干旱胁迫后均表现为增加。
2.6 干旱胁迫对水稻川香29B近等基因导入系苗期叶片保护性酶活性的影响
从表7可得,干旱胁迫均显著提高了水稻幼苗叶片中各类保护性酶活性,其中CAT、SOD、POD在第1次干旱胁迫后增幅高于第2次干旱胁迫,而吡咯碄-5-羧酸合成酶、鸟氨酸转氨酶、脯氨酸脱氢酶则是第2次干旱胁迫后增幅高于第1次干旱胁迫。所有供试材料的6个保护性酶活性在两次干旱胁迫后均表现为增长。说明干旱胁迫下,幼苗叶片保护性酶活性提高,以此抵御环境的不利影响。

表7 干旱胁迫对川香29B近等基因导入系叶片保护性酶活性的影响Table 7 Effects of drought stress on the activities of protective enzymes in Chuanxiang 29B NIILs leaves
2.7 干旱胁迫对水稻川香29B近等基因导入系幼苗存活率的影响
从图1可以看出,参试材料经干旱胁迫后,存活率均存在不同幅度下降,两次干旱胁迫后各材料存活率排序较为一致。第1次干旱胁迫后,6份材料干旱存活率在74.88%~90.86%之间,C1干旱存活率显著高于其他5份材料,C2、C5、C6干旱存活率居中;第2次干旱存活率在66.34%~82.86%之间,C1干旱存活率最高,C2、C5、C6干旱存活率居中,C3、C4干旱存活率显著低于前述4份材料;对于反复干旱存活率,C1反复干旱存活率显著高于其他5份材料,C3、C4反复存活率显著低于其他4份材料。据此可得,C1抗旱性最强, C3、C4抗旱性最弱,C2、C5和C6抗旱性中等。
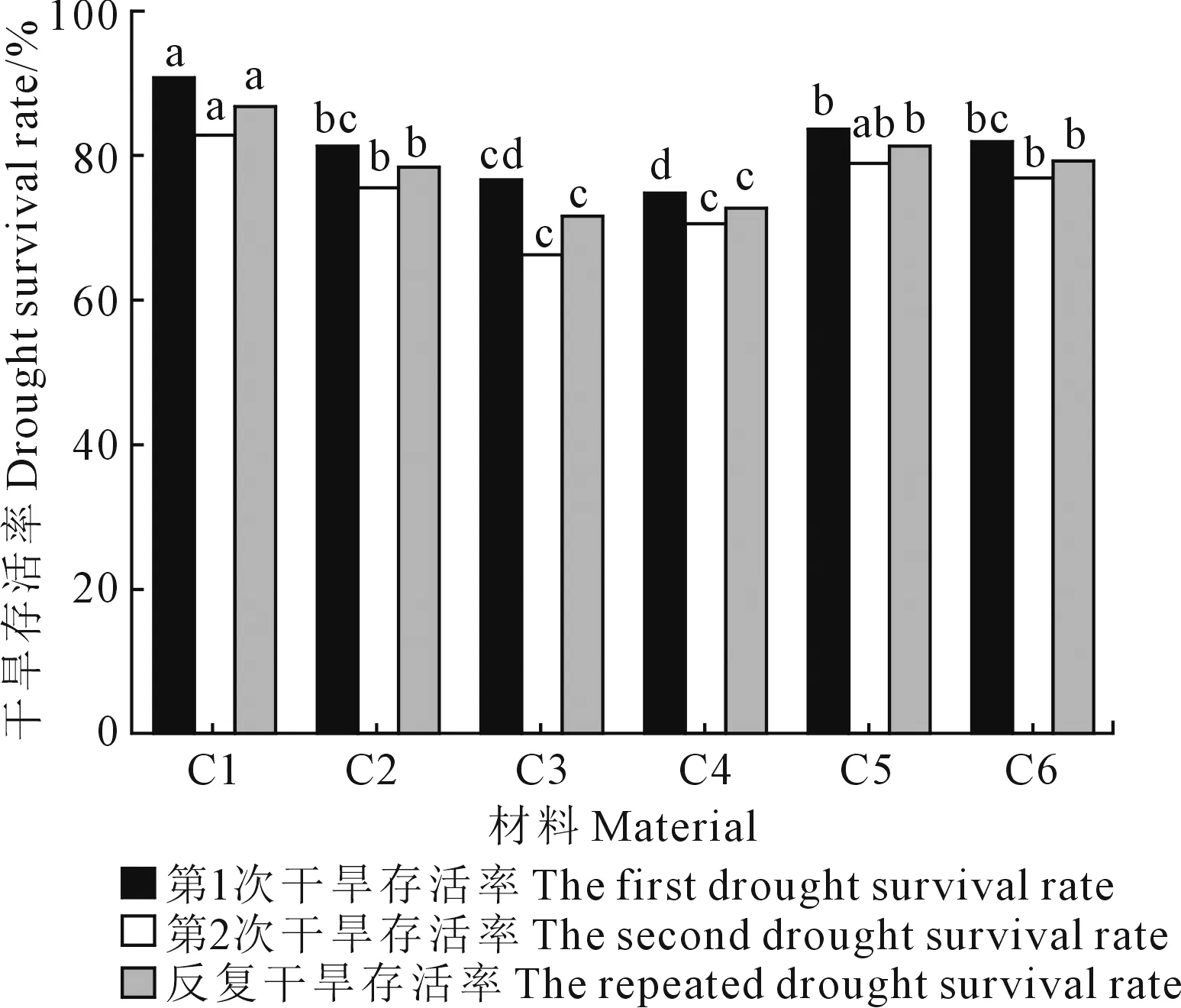
注:不同小写字母表示同一指标不同材料间的差异显著(P<0.05)。Note: The different lowercase letters indicate significant differences among different materials within the same index (P<0.05). 图1 干旱胁迫对川香29B近等基因导入系幼苗干旱存活率的影响Fig.1 Effects of drought stress on drought survival rate of Chuanxiang 29B NIILs seedlings
2.8 干旱胁迫后指标相对值与苗期反复干旱存活率的相关性分析
从表8可知,水稻苗期反复干旱存活率与第1次干旱胁迫后叶片中游离氨基酸含量、脱落酸含量的相对值呈显著正相关关系;与第2次干旱胁迫根表面积的相对值呈显著正相关关系,与根粗、过氧化物酶、叶片可溶性糖含量呈显著负相关关系。
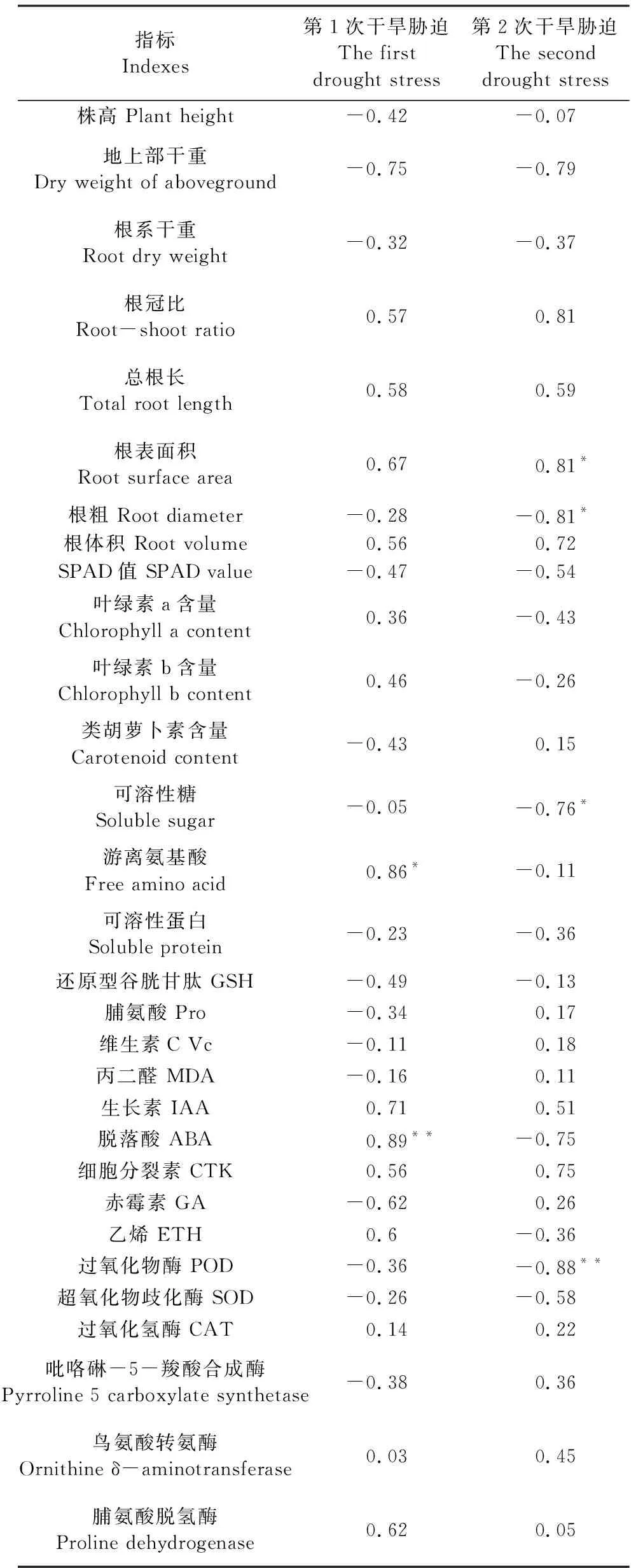
表8 苗期生理形态指标相对值与反复干旱存活率的相关系数Table 8 Correlation coefficient between the relative values of physiological morphological indicators and repeated drought survival rate at seedling stage
2.9 水稻苗期抗旱性预测
以反复干旱存活率作为因变量,第2次干旱胁迫后指标的相对值作自变量,通过逐步回归分析,得回归方程:
Y=98.90-6.82X1+20.20X2+7.08X3-33.80X4
式中,X1、X2、X3、X4分别代表第2次干旱胁迫后总根长、根表面积、叶片中GSH和POD酶活性的相对值,方程决定系数R2=0.999,F=794.53**,达极显著。利用回归方程对观察值进行拟合计算,拟合相对误差在0.003%~0.019%之间。可用这4项指标作为反复干旱后预测水稻早期抗旱性的指标。
3 讨 论
3.1 水稻近等基因系苗期水分胁迫下根系形态与抗旱性关系
干旱胁迫信号传递到水稻根系,首先引发根系变化,相关抗旱基因开始表达[21-22],同时信号进一步向地上部分传递,进而调控地上部光合生产、物质转运、形态建成等生理生化过程,进而影响产量形成。根系是植株吸收水分的重要器官,根系健壮发达可提高植株吸水能力,减轻干旱胁迫效应,因此根系发达程度常被作为抗旱鉴定指标。前人研究表明,根体积、总根长、最长根长、根干重、根冠比、根系相对含水量等根系相关指标可作为抗旱性鉴定的指标[23-25]。本研究中,水稻苗期两次干旱胁迫后,供试材料根系干重、根冠比均比对照增加,在第1次干旱胁迫后,抗旱性较强的品种C1、C2和C6总根长、根表面积比对照增加,在第2次干旱胁迫后,C1、C6总根长、根表面积呈增加趋势,表明根系变长、面积扩大是水稻早期适应干旱环境的重要形态变化。相关分析表明,第2次干旱胁迫后根表面积相对值与反复干旱存活率显著正相关,逐步回归分析中第2次干旱胁迫后总根长、根表面积纳入预测方程。综上可得,根表面积、总根长可以作为鉴定水稻抗旱性的形态指标。
3.2 水稻近等基因系苗期水分胁迫后叶片生理生化特性与抗旱性关系
干旱胁迫降低植株的叶绿素含量,但类胡萝卜素含量随时间呈增加趋势[26],本研究中C2、C4、C6在第2次干旱胁迫后类胡萝卜素含量高于对照,与前人研究结果一致。在干旱胁迫下,植株可溶性蛋白质、游离氨基酸、可溶性糖等渗透调节物质增加[27],AsA、GSH等抗氧化物质含量提高[6],SOD、CAT、POD等保护性酶活性上调[28-29],并通过植物激素类调节抗旱性[30-32],这些物质变化一定程度上反映了植株应对干旱胁迫的能力。前人研究得出可溶性蛋白质、FAA、AsA、GSH和MDA含量及POD、SOD、CAT活性的相对值与反复干旱存活率呈显著或极显著的相关性,可作为水稻苗期抗旱性鉴定的生理生化指标[6, 33]。本研究结果表明,干旱胁迫导致水稻叶片SPAD值降低,叶绿素a和叶绿素b含量显著减少,幼苗叶片可溶性糖、氨基酸和Vc含量显著下降,可溶性蛋白质、脯氨酸和MDA含量明显上升,第1次干旱胁迫后GSH含量增加,第2次干旱胁迫后GSH含量降低,生长素、细胞分裂素、赤霉素两次干旱胁迫下均降低,脱落酸、乙烯均表现升高,SOD、POD、CAT、吡咯碄-5-羧酸合成酶、鸟氨酸转氨酶、脯氨酸脱氢酶两次干旱胁迫后均升高,表明干旱胁迫下水稻幼苗在渗透调节物质、氧化还原物质上产生了相应变化,并在激素、保护性酶的作用下进行调节以抵御不良环境因子。经过回归分析,叶片中GSH和POD的相对值进入回归方程,作为反复干旱后预测水稻早期抗旱性的指标。
3.3 苗期反复干旱法的利用及不同时期抗旱性比较
苗期采用反复干旱法进行干旱胁迫可以鉴定作物的抗旱性[34],操作简便、直观快速,得到了大量应用。本研究采用反复干旱存活率评价,得出供试材料抗旱性强弱排序为:C1>C5>C6>C2>C4>C3,这个结果与全生育期评价结果并不一致,王兴荣等[35]对大豆种质资源研究得出苗期和全生育期抗旱性呈不显著负相关,这与本研究结果类似,与小麦抗旱性的研究结果[36]相似。不同时期的评价结果不一致,可能是由于植株处于营养生长、生殖生长的不同阶段其生长发育的中心活动并不相同,苗期重在保持存活和植株生长,成熟期则在产量形成,其受干旱影响的后果并不相同,不同时期干旱胁迫下用于评价的指标也并不尽相同,由此造成评价结果的差异。同时,作物抗旱性是遗传与环境互作而决定的多基因性状,同时通过对评价抗旱性所采用的形态学性状、产量和产量相关性状以及根部性状的遗传分析表明,这些性状属于多基因控制的复杂性状,具有显著的加性、显性及上位性遗传效应,并与环境互作作用显著[37],这进一步增加了评价工作的复杂性。潘雅姣等[38]通过研究对水稻材料在苗期和分蘖期进行干旱胁迫前后DNA甲基化情况,得出干旱胁迫会引起水稻DNA甲基化水平及状态发生改变,且这种改变具有一定品种特异性和时空特异性,同一材料在不同时期下都会存在差异。因此,水稻抗旱性决定机制复杂,不同生育时期间、不同环境条件下都存在差异,而苗期与全生育期存在的差异原因有待于更深一步研究,明确其不同生育期干旱下的内在联系,以便更准确地开展抗旱性鉴定。
4 结 论
1)反复干旱存活率可以直观、快速地鉴定水稻苗期抗旱性,可作为苗期抗旱鉴定首要指标。通过反复干旱法进行苗期胁迫,以反复干旱存活率评价,苗期川香29B/ASOMINORI//川香29B///川香29B(C1)抗旱性最强。
2)通过相关分析和回归分析表明,根表面积、总根长、根粗可作为水稻苗期抗旱鉴定的形态指标,叶片中GSH含量和POD活性的相对值等可作为鉴定水稻苗期抗旱性的生理生化指标。
干旱对粮食作物生产带来的巨大影响难以忽视,必须总结形成相应的问题化解方案,近年来在抗旱品种选育、抗旱基因资源挖掘、抗旱生理生化机制等方面开展了很多研究,育成抗旱品种是应对干旱因子的重要手段,如何科学、准确、统一地鉴定评价品种、材料的抗旱性,从而对不同作物材料、品种等形成标准化的抗旱鉴定结果以便于横向比较,是十分必要的,这有待更全面深入研究。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:09:12
今日农业(2021年9期)2021-11-26 07:41:24
建材发展导向(2021年11期)2021-07-28 06:57:22
今日农业(2020年23期)2020-12-15 03:48:26
当代水产(2020年10期)2020-03-17 07:02:48
当代水产(2019年8期)2019-10-12 08:57:26
今日农业(2019年12期)2019-08-13 00:50:10
浙江农业学报(2017年1期)2017-05-17 06:13:36
西南农业学报(2016年4期)2016-05-17 05:42:13
山西农经(2015年7期)2015-07-10 15:00:57
